Effects of Different Grazing Intensities on Soil C, N, and P in an Alpine Meadow on the Qinghai—Tibetan Plateau, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
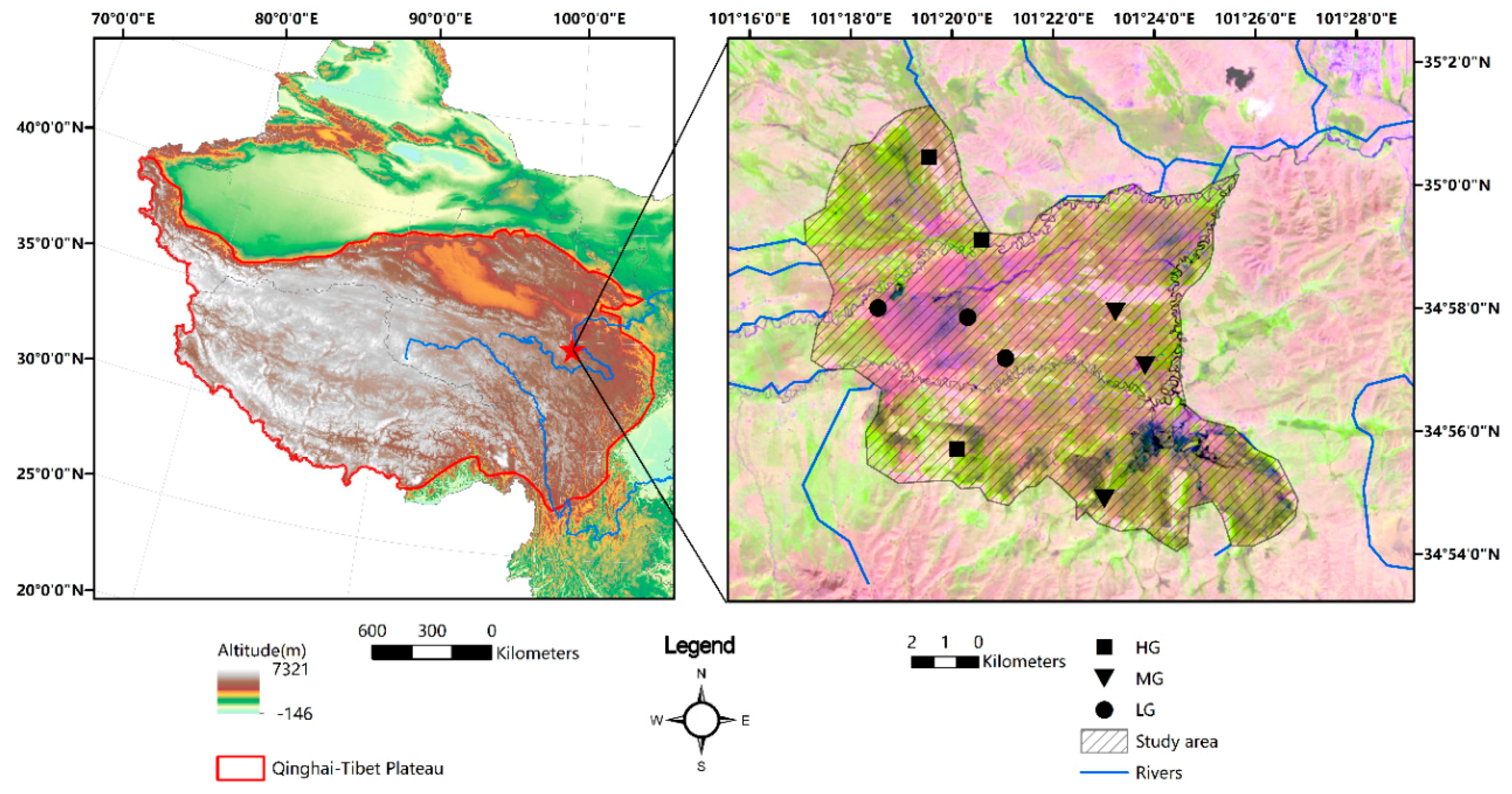
2.1. Study Area
2.2. Experimental Design and Sampling
2.3. Data Statistics and Analysis
3. Results
3.1. Effect of Grazing on Biomass
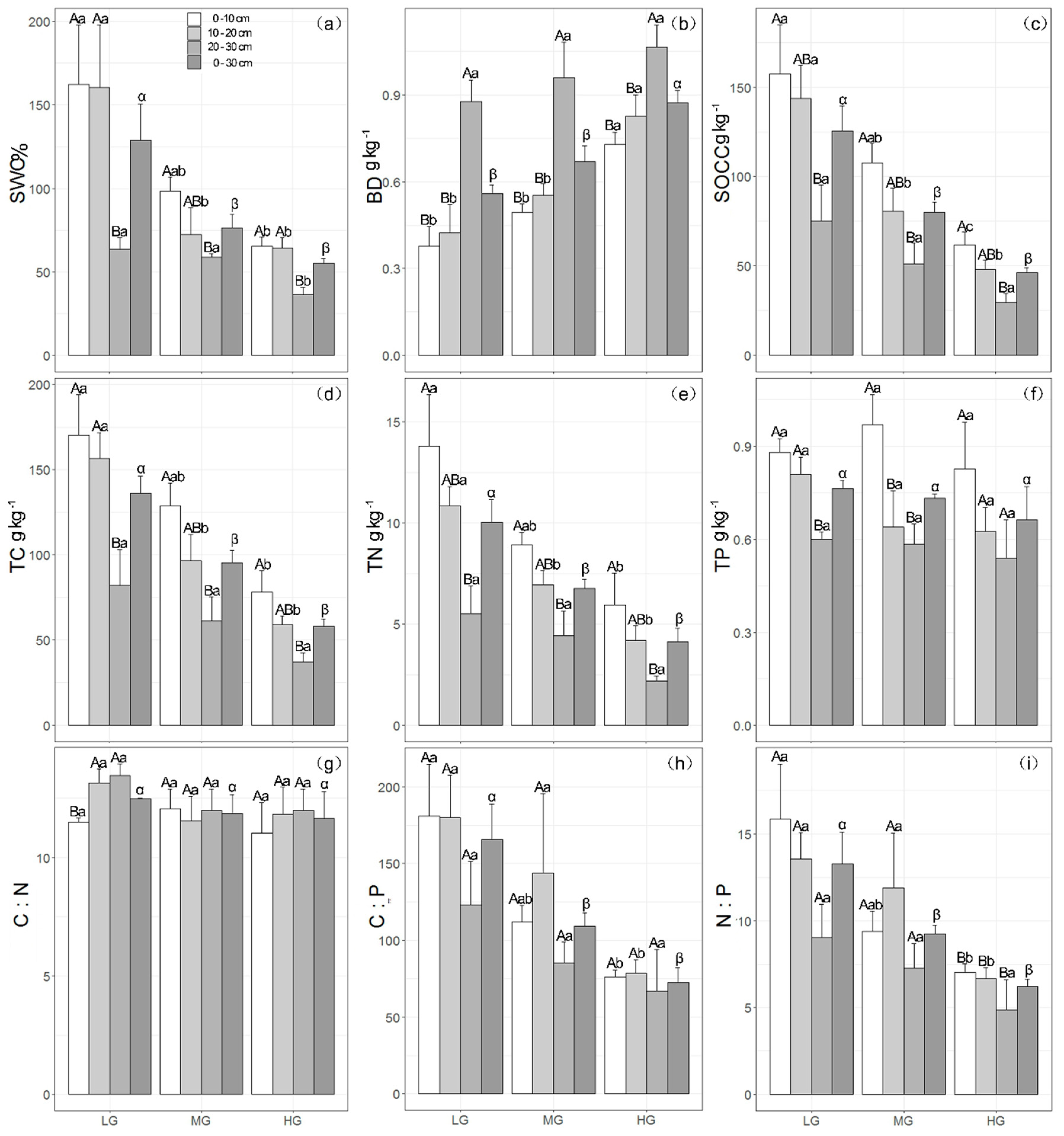
3.2. Effect of Grazing on Soil Physical Properties
3.3. Effect of Grazing on Soil Chemical Properties
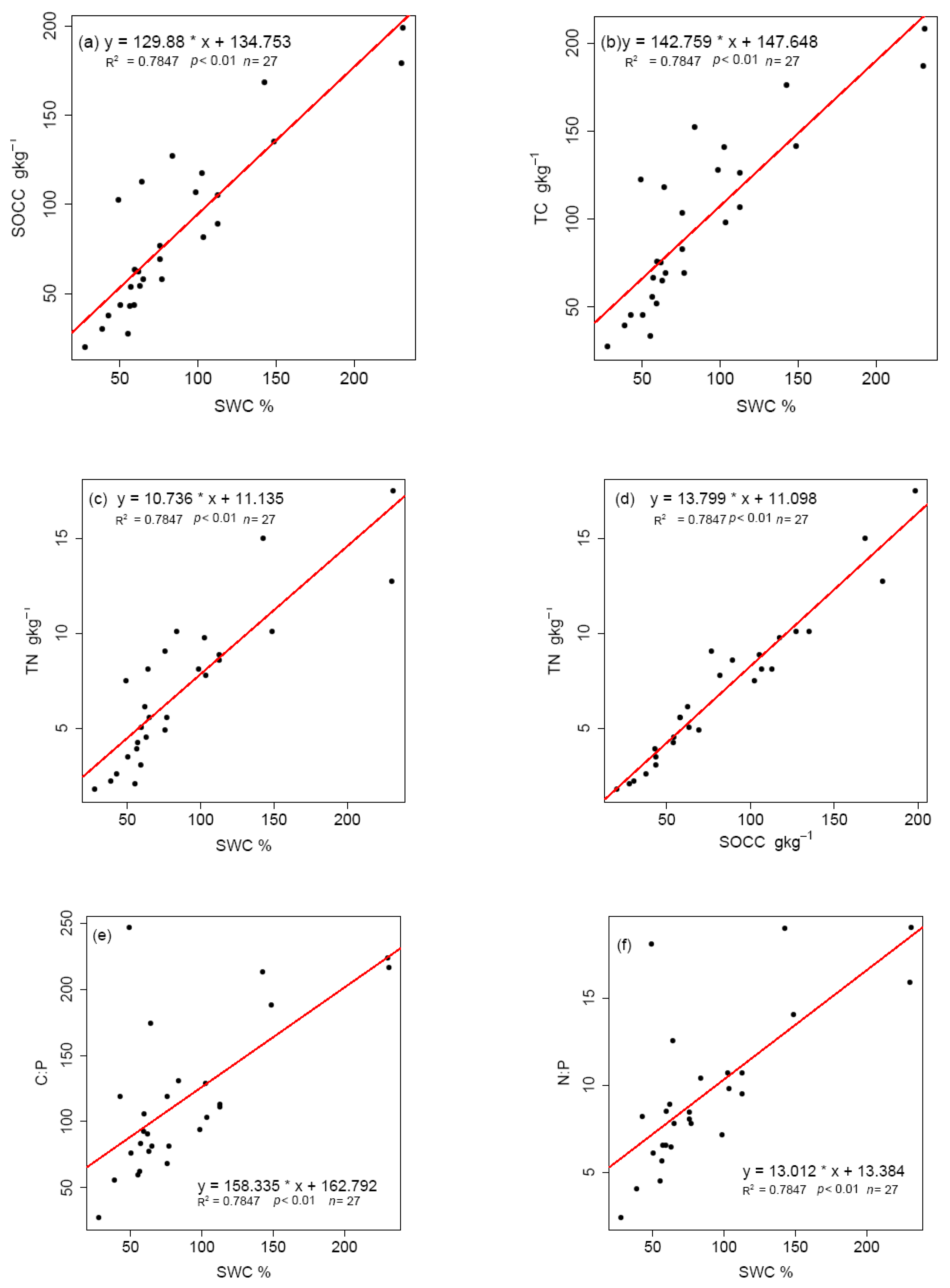
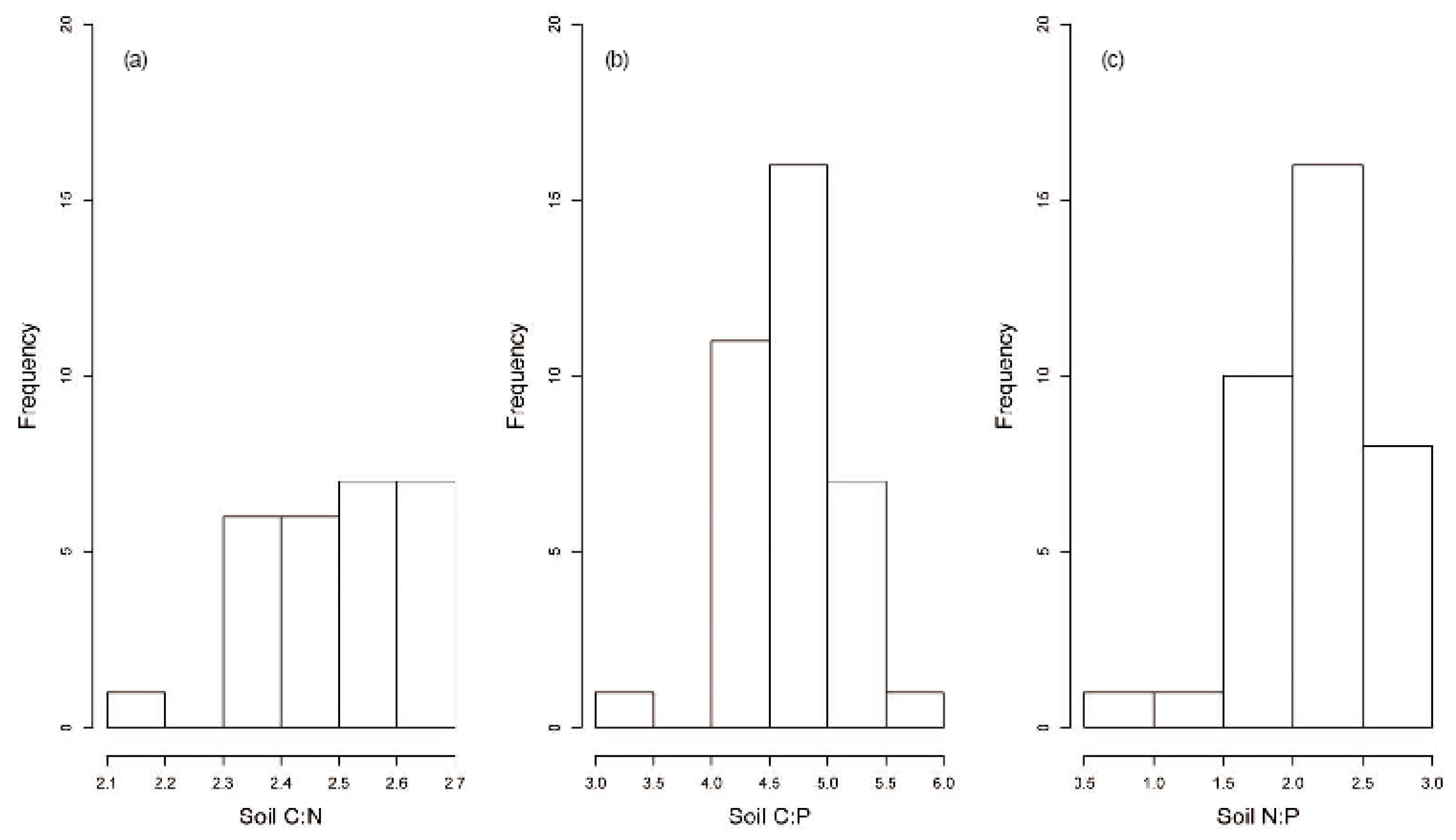
3.4. Effect of Grazing on Stoichiometric Variation in Soil C, N, and P
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- Yang, Z.; Zhu, Q.; Zhan, W.; Xu, Y.; Zhu, E.; Gao, Y.; Li, S.; Zheng, Q.; Zhu, D.; He, Y.; et al. The linkage between vegetation and soil nutrients and their variation under different grazing intensities in an alpine meadow on the eastern Qinghai-Tibetan Plateau. Ecol. Eng. 2018, 110, 128–136. [Google Scholar] [CrossRef]
- Gao, Y.H.; Luo, P.; Wu, N.; Chen, H.; Wang, G.X. Grazing Intensity Impacts on Carbon Sequestration in an Alpine Meadow on the Eastern Tibetan Plateau. Res. J. Agric. Biol. Sci. 2007, 3, 642–647. [Google Scholar]
- Rui, Y.; Wang, Y.; Chen, C.; Zhou, X.; Wang, S.; Xu, Z.; Duan, J.; Kang, X.; Lu, S.; Luo, C. Warming and grazing increase mineralization of organic P in an alpine meadow ecosystem of Qinghai-Tibet Plateau, China. Plant Soil 2012, 357, 73–87. [Google Scholar] [CrossRef]
- Wen, L.; Dong, S.; Li, Y.; Li, X.; Shi, J.; Wang, Y.; Liu, D.; Ma, Y. Effect of Degradation Intensity on Grassland Ecosystem Services in the Alpine Region of Qinghai-Tibetan Plateau, China. PLoS ONE 2013, 8, e58432. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Ding, K.; Li, Z. Comparison of soil carbon and nitrogen stocks at grazing-excluded and yak grazed alpine meadow sites in Qinghai-Tibetan Plateau, China. Ecol. Eng. 2016, 87, 203–211. [Google Scholar] [CrossRef]
- Su, Y.Z.; Li, Y.L.; Cui, J.Y.; Zhao, W.Z. Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, northern China. Catena 2005, 59, 267–278. [Google Scholar]
- Zheng, S.; Ren, H.; Li, W.; Lan, Z. Scale-dependent effects of grazing on plant C: N: P stoichiometry and linkages to ecosystem functioning in the Inner Mongolia grassland. PLoS ONE 2012, 7, e51750. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Huang, H.Z.; Zhang, Z.N.; Wu, G.L. Effects of grazing on the soil properties and C and N storage in relation to biomass allocation in an alpine meadow. J. Soil Sci. Plant Nutr. 2011, 11, 27–39. [Google Scholar] [Green Version]
- Talore, D.G.; Tesfamariam, E.H.; Hassen, A.; Toit, J.D.; Klampp, K.; Jean-Francois, S. Long-term impacts of grazing intensity on soil carbon sequestration and selected soil properties in the arid Eastern Cape, South Africa. J. Sci. Food Agric. 2016, 96, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Shi, P.; Zhang, X.; Zou, C.B. Effects of grazing exclusion on carbon sequestration and plant diversity in grasslands of China—A meta-analysis. Ecol. Eng. 2016, 94, 647–655. [Google Scholar] [CrossRef] [Green Version]
- Qasim, S.; Gul, S.; Shah, M.H.; Hussain, F.; Ahmad, S.; Islam, M.; Rehman, G.; Yaqoob, M.; Shah, S.Q. Influence of grazing exclosure on vegetation biomass and soil quality. Int. Soil Water Conserv. Res. 2017, 5, 62–68. [Google Scholar] [CrossRef]
- Yan, L.; Zhou, G.; Zhang, F. Effects of different grazing intensities on grassland production in China: A meta-analysis. PLoS ONE 2013, 8, e81466. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Zhou, X.; He, Y.; Shao, J.; Hu, Z.; Liu, R.; Zhou, H.; Hosseinibai, S. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis. Glob. Chang. Biol. 2017, 23, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; He, J.S.; Zhang, S.; Lechowicz, M.J. Tradeoffs between forage quality and soil fertility: Lessons from Himalayan rangelands. Agric. Ecosyst. Environ. 2016, 234, 31–39. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.; Wang, H.; Huang, Q. Comprehensive analysis of grazing intensity impacts soil organic carbon: A case study in typical steppe of Inner Mongolia, China. Appl. Soil Ecol. 2018. [Google Scholar] [CrossRef]
- Niu, K.; He, J.S.; Lechowicz, M.J. Grazing-induced shifts in community functional composition and soil nutrient availability in Tibetan alpine meadows. J. Appl. Ecol. 2016, 53, 1554–1564. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, G.; Mou, N.; Xie, Y.; Xu, Z.; Li, Y. Assessment of Carbon Storage and Its Influencing Factors in Qinghai-Tibet Plateau. Sustainability 2018, 10, 1864. [Google Scholar] [CrossRef]
- Liu, S.; Zamanian, K.; Schleuss, M.; Zarebanadkouki, M.; Kuzyakov, Y. Degradation of Tibetan grasslands: Consequences for carbon and nutrient cycles. Agric. Ecosyst. Environ. 2017, 252, 93–104. [Google Scholar] [CrossRef]
- Lu, X.; Yan, Y.; Jian, S.; Zhang, X.; Chen, Y.; Wang, X.; Cheng, G. Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: Effects of grazing exclusion. Ecol. Evol. 2015, 5, 4492–4504. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.; Hastings, A.; Chadwick, D.R.; Jones, D.L.; Evans, C.D.; Jones, M.B.; Rees, R.M.; Smith, P. Critical review of the impacts of grazing intensity on soil organic carbon storage and other soil quality indicators in extensively managed grasslands. Agric. Ecosyst. Environ. 2018, 253, 62–81. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.K.; Wen, L.; Li, Y.Y.; Wang, X.X.; Zhu, L.; Li, X.Y. Soil-Quality Effects of Grassland Degradation and Restoration on the Qinghai-Tibetan Plateau. Soil Sci. Soc. Am. J. 2012, 76, 2256–2264. [Google Scholar] [CrossRef]
- Hu, Z.; Li, S.; Guo, Q.; Niu, S.; He, N.; Li, L.; Yu, G. A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China. Glob. Chang. Biol. 2016, 22, 1385–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Roldán, E.; Paz-Ferreiro, J.; Bardgett, R.D. Grazing exclusion affects soil and plant communities, but has no impact on soil carbon storage in an upland grassland. Agric. Ecosyst. Environ. 2012, 149, 118–123. [Google Scholar] [CrossRef]
- Steffens, M.; Kölbl, A.; Totsche, K.U.; Kögel-Knabner, I. Grazing effects on soil chemical and physical properties in a semiarid steppe of Inner Mongolia (P.R. China). Geoderma 2008, 143, 63–72. [Google Scholar] [CrossRef]
- Raiesi, F.; Riahi, M. The influence of grazing exclosure on soil C stocks and dynamics, and ecological indicators in upland arid and semi-arid rangelands. Ecol. Indic. 2014, 41, 145–154. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Paul, E.A. Soil Microbiology, Ecology, and Biochemistry, 3rd ed.; Paul, E.A., Ed.; Academic Press: Burlington, MA, USA, 2007; pp. 389–430. [Google Scholar]
- Sophie, Z.-B.; Maria, K.K.; Maria, M.; Josep, P.; Andreas, R.; Jordi, S.; Wolfgang, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar]
- Zhao, F.; Jiao, S.; Ren, C.; Di, K.; Jian, D.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Land use change influences soil C, N, and P stoichiometry under ‘Grain-to-Green Program’ in China. Sci. Rep. 2015, 5, 10195. [Google Scholar]
- McDowell, R.W.; Stewart, I. The phosphorus composition of contrasting soils in pastoral, native and forest management in Otago, New Zealand: Sequential extraction and 31P NMR. Geoderma 2006, 130, 176–189. [Google Scholar] [CrossRef]
- Stutter, M.I.; Shand, C.A.; George, T.S.; Blackwell, M.S.A.; Dixon, L.; Bol, R.; MacKay, R.L.; Richardson, A.E.; Condron, L.M.; Haygarth, P.M. Land use and soil factors affecting accumulation of phosphorus species in temperate soils. Geoderma 2015, 257–258, 29–39. [Google Scholar] [CrossRef]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Gao, Y.H.; Schumann, M.; Chen, H.; Ning, W.; Peng, L. Impacts of grazing intensity on soil carbon and nitrogen in an alpine meadow on the eastern Tibetan Plateau. Appl. Ecol. Environ. Res. 2009, 6, 69–79. [Google Scholar] [CrossRef]
- Shibin, L.; Per-Marten, S.; Yakov, K. Carbon and Nitrogen Losses from Soil Depend on Degradation of Tibetan Kobresia Pastures. Land Degrad. Dev. 2017, 28, 1253–1262. [Google Scholar]
- Bing, H.; Wu, Y.; Zhou, J.; Sun, H.; Luo, J.; Wang, J.; Yu, D. Stoichiometric variation of carbon, nitrogen, and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of Eastern Tibetan Plateau. J. Soils Sediments 2016, 16, 405–416. [Google Scholar] [CrossRef]
- Feng, D.; Bao, W. Review of the temporal and spatial patterns of soil C:N:P stoichiometry and its driving factors. Chin. J. Appl. Environ. Biol. 2017, 23, 400–408. [Google Scholar]
- Tipping, E.; Somerville, C.J.; Luster, J. The C:N:P:S stoichiometry of soil organic matter. Biogeochemistry 2016, 130, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Li, X.; Li, R.; Tian, L.; Zhang, T. Effect of grazing exclusion on ecosystem respiration among three different alpine grasslands on the central Tibetan Plateau. Ecol. Eng. 2016, 94, 599–607. [Google Scholar] [CrossRef]
- Ni, J. Carbon storage in grasslands of China. J. Arid Environ. 2002, 50, 205–218. [Google Scholar] [CrossRef]
- Tian, H.; Wang, S.; Liu, J.; Pan, S.; Chen, H.; Zhang, C.; Shi, X. Patterns of soil nitrogen storage in China. Glob. Biogeochem. Cycles 2006, 20. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Ma, Z.; Zhang, F.; Su, C. Suggestion on protection and construction of grassland in Zeku County, Qinghai Province. Grassland Turf 2009, 3, 95–97. [Google Scholar]
- Chang, X.; Zhu, X.; Wang, S.; Cui, S.; Luo, C.; Zhang, Z.; Wilkes, A. Impacts of management practices on soil organic carbon in degraded alpine meadows on the Tibetan Plateau. Biogeosciences 2014, 11, 3495–3503. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, L.; Wei, Y.; Li, H.; Li, Y. Effects of Degradation Degree on Soil Physicochemical Properties and Soil Water-Holding Capacity in Zeku Alpine Meadow in the Headwater Region of Three Rivers in China. Chin. J. Grassland 2017, 39, 54–61. [Google Scholar]
- Hafner, S.; Unteregelsbacher, S.; Seeber, E.; Lena, B.; Xu, X.; Li, X.; Guggenberger, G.; Miehe, G.; Kuzyakov, Y. Effect of grazing on carbon stocks and assimilate partitioning in a Tibetan montane pasture revealed by 13CO2 pulse labeling. Glob. Chang. Biol. 2012, 18, 528–538. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, W.; Zhang, L.; Gong, B.; Xu, C.; Feng, H.; Wu, Z. A comparative analysis of herdsmen and eco-immigrant’s livelihood under ecological compensation of the Source Area of Three Rivers: A case study base on household investivation in Zeku Country, Qinghai Province. Pratacultural Sci. 2014, 31, 1178–1184. [Google Scholar]
- Yang, Y.; Mohammat, A.; Feng, J.; Zhou, R.; Fang, J. Storage, patterns and environmental controls of soil organic carbon in China. Biogeochemistry 2007, 84, 131–141. [Google Scholar] [CrossRef]
- Shang, Z.; Feng, Q.; Wu, G.; Ren, G.; Long, R. Grasslandification has significant impacts on soil carbon, nitrogen and phosphorus of alpine wetlands on the Tibetan Plateau. Ecol. Eng. 2013, 58, 170–179. [Google Scholar] [CrossRef]
- An, H.; Li, G. Effects of grazing on carbon and nitrogen in plants and soils in a semiarid desert grassland, China. J. Arid Land 2015, 7, 341–349. [Google Scholar] [CrossRef]
- Ge, G.; Li, Z.; Fan, F.; Chu, G.; Hou, Z.; Liang, Y. Soil biological activity and their seasonal variations in response to long-term application of organic and inorganic fertilizers. Plant Soil 2010, 326, 31. [Google Scholar] [CrossRef]
- Yuan, J.; Ouyang, Z.; Zheng, H.; Xu, W. Effects of different grassland restoration approaches on soil properties in the southeastern Horqin sandy land, northern China. Appl. Soil Ecol. 2012, 61, 34–39. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Liu, Y.; Xiao, L.; Huang, Y. How Fencing Affects the Soil Quality and Plant Biomass in the Grassland of the Loess Plateau. Int. J. Environ. Res. Public Health 2017, 14, 1117. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Hao, X.; Zhao, M.; Wang, M.; Benh, E.; Walter, W.; Wang, M. Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 125, 21–32. [Google Scholar] [CrossRef]
- Ritchie, M.E. Plant compensation to grazing and soil carbon dynamics in a tropical grassland. PeerJ 2014, 2, e233. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Wu, X. Effects of grazing intensity on soil organic carbon of rangelands in Xilin Gol League, Inner Mongolia, China. J. Geog. Sci. 2016, 26, 1550–1560. [Google Scholar] [CrossRef]
- Rickart, E.A.; Bienek, K.G.; Rowe, R.J. Impact of Livestock Grazing on Plant and Small Mammal Communities in the Ruby Mountains, Northeastern Nevada. West. N. Am. Nat. 2013, 73, 505–515. [Google Scholar] [CrossRef]
- Guo, Y.; Du, Q.; Li, G.; Ni, Y.; Zhang, Z.; Ren, W.; Hou, X. Soil phosphorus fractions and arbuscular mycorrhizal fungi diversity following long-term grazing exclusion on semi-arid steppes in Inner Mongolia. Geoderma 2016, 269, 79–90. [Google Scholar] [CrossRef]
- Wang, X.; Yan, Y.; Cao, Y. Impact of historic grazing on steppe soils on the northern Tibetan Plateau. Plant Soil 2012, 354, 173–183. [Google Scholar] [CrossRef]
- McSherry, M.E.; Ritchie, M.E. Effects of grazing on grassland soil carbon: A global review. Glob. Chang. Biol. 2013, 19, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wu, J.; Clark, C.M.; Pan, Q.; Zhang, L.; Chen, S.; Wang, Q.; Han, X.; Wisley, B. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J. Appl. Ecol. 2012, 49, 1204–1215. [Google Scholar] [CrossRef] [Green Version]
- Feng, R.; Long, R.; Shang, Z.; Ma, Y.; Dong, S.; Wang, Y. Establishment of Elymus natans improves soil quality of a heavily degraded alpine meadow in Qinghai-Tibetan Plateau, China. Plant Soil 2010, 327, 403–411. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Domanski, G. Carbon input by plants into the soil: Review. J. Plant Nutr. Soil Sci. 2015, 163, 421–431. [Google Scholar] [CrossRef]
- Li, Y.S.; Wu, L.H.; Lu, X.H.; Zhao, L.M.; Fan, Q.L.; Zhang, F.S. Soil microbial biomass as affected by non-flooded plastic mulching cultivation in rice. Biol. Fertil. Soils 2006, 43, 107–111. [Google Scholar] [CrossRef]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Hiltbrunner, D.; Schulze, S.; Hagedorn, F.; Schmidt, M.W.I.; Zimmmermann, S. Cattle trampling alters soil properties and changes soil microbial communities in a Swiss sub-alpine pasture. Geoderma 2012, 170, 369–377. [Google Scholar] [CrossRef]
- Jiao, F.; Shi, X.R.; Han, F.P.; Yuan, Z.Y. Increasing aridity, temperature and soil pH induce soil C-N-P imbalance in grasslands. Sci. Rep. 2016, 6, 19601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raliya, R.; Tarafdar, J.C.; Biswas, P. Enhancing the Mobilization of Native Phosphorus in the Mung Bean Rhizosphere Using ZnO Nanoparticles Synthesized by Soil Fungi. J. Agric. Food Chem. 2016, 64, 3111–3118. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, W.H. An Analysis of Global Change. Biogeochemistry 1997, 2, 40–41. [Google Scholar]
- Simpson, R.J.; Stefanski, A.; Marshall, D.J.; Moore, A.D.; Richardson, A.E. Management of soil phosphorus fertility determines the phosphorus budget of a temperate grazing system and is the key to improving phosphorus efficiency. Agric. Ecosyst. Environ. 2015, 212, 263–277. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Ellert, B.H.; Bettany, J.R. Calculation of organic matter and nutrients stored in soils under contrasting management regimes. Can. J. Soil Sci. 1995, 75, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Berthrong, S.T.; Piñeiro, G.; Jobbágy, E.G.; Jackson, R.B. Soil C and N changes with afforestation of grasslands across gradients of precipitation and plantation age. Ecol. Appl. 2012, 22, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Sites | n | SL (cm) | GI (Yaks ha−1) | ABio (g m−2) | BBio (g m−2) | SOCC (g kg−1) | TN (g kg−1) | TP (g kg−1) | C:N:P | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
| Hongyuan, QTP | 3 | 0–15 | 0.00 | 49.08 | 4.05 | 0.92 | 49:4:1 | [1] | ||
| 3 | 0–15 | 0.71 | 67.85 | 5.93 | 1.06 | 68:6:1 | ||||
| 3 | 0–15 | 1.20 | 59.03 | 5.03 | 0.98 | 59:5:1 | ||||
| 3 | 0–15 | 1.58 | 54.89 | 4.72 | 1.02 | 55:5:1 | ||||
| Hongyuan, QTP | 5 | 0–30 | 1.20 | 521 | 1798 | 9795 * | [2] | |||
| 5 | 0–30 | 2.00 | 589 | 2482 | 10158 * | |||||
| 5 | 0–30 | 2.90 | 392 | 2923 | 11729 * | |||||
| Maqu, QTP | 3 | 0–15 | 1.08 | 299.7 | 715.5 | 43.13 | 4.06 | 0.61 | 71:7:1 | [8] |
| 3 | 0–15 | 1.36 | 231.5 | 1478.3 | 52.08 | 4.26 | 0.76 | 68:6:1 | ||
| 3 | 0–15 | 2.13 | 114.2 | 1899.4 | 60.83 | 5.70 | 0.80 | 76:7:1 | ||
| Zeku, QTP | 3 | 0–10 | 0.19 | 123.3 ± 3.8 | 1219.3 ± 193.9 | 157.4 ± 27.5 | 13.8 ± 2.6 | 0.9 ± 0.0 | 175:15:1 | This |
| 3 | 0–10 | 0.53 | 83.7 ± 9.9 | 1152.4 ± 203.4 | 107.7 ± 11.0 | 8.9 ± 0.6 | 1.0 ± 0.1 | 108:8:1 | study | |
| 3 | 0–10 | 1.42 | 20.0 ± 1.6 | 861.6 ± 116.5 | 61.6 ± 7.5 | 5.9 ± 1.6 | 0.8 ± 0.2 | 77:7:1 |
| SL (cm) | SOCS (t ha−1) | STNS (t ha−1) | STPS (t ha−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LG | MG | HG | LG | MG | HG | LG | MG | HG | |
| 0–10 | 56.4 ± 15.7de | 53.3 ± 11.8de | 44.3 ± 4.4de | 61.4 ± 14.7DE | 63.8 ± 14.1DE | 55.9 ± 9.1DE | 0.33 ± 0.10γ | 0.47 ± 0.05γ | 0.59 ± 0.12γ |
| 10–20 | 57.7 ± 11.4de | 43.6 ± 7.3de | 39.0 ± 0.9de | 63.9 ± 17.7DE | 52.1 ± 8.9DE | 47.9 ± 3.1DE | 0.35 ± 0.17γ | 0.36 ± 0.14γ | 0.51 ± 0.12γ |
| 20–30 | 64.6 ± 29.7d | 46.3 ± 11.4de | 30.8 ± 7.3e | 70.2 ± 29.6D | 55.4 ± 13.7DE | 39.2 ± 8.1E | 0.53 ± 0.08γ | 0.55 ± 0.03γ | 0.59 ± 0.27γ |
| 0–30 | 208.7 ± 33.4a | 158.5 ± 8.5b | 120.9 ± 5.1c | 227.2 ± 22.2A | 189.6 ± 10.0B | 151.4 ± 12.0C | 1.26 ± 0.15β | 1.47 ± 0.20αβ | 1.72 ± 0.42α |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Zhang, Z.; Shi, L.; Zhou, Y.; Yang, M.; Cao, J.; Wu, S.; Lei, G. Effects of Different Grazing Intensities on Soil C, N, and P in an Alpine Meadow on the Qinghai—Tibetan Plateau, China. Int. J. Environ. Res. Public Health 2018, 15, 2584. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112584
Li G, Zhang Z, Shi L, Zhou Y, Yang M, Cao J, Wu S, Lei G. Effects of Different Grazing Intensities on Soil C, N, and P in an Alpine Meadow on the Qinghai—Tibetan Plateau, China. International Journal of Environmental Research and Public Health. 2018; 15(11):2584. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112584
Chicago/Turabian StyleLi, Gang, Zhi Zhang, Linlu Shi, Yan Zhou, Meng Yang, Jiaxi Cao, Shuhong Wu, and Guangchun Lei. 2018. "Effects of Different Grazing Intensities on Soil C, N, and P in an Alpine Meadow on the Qinghai—Tibetan Plateau, China" International Journal of Environmental Research and Public Health 15, no. 11: 2584. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112584