Salmonella enterica Serovar Typhimurium and Escherichia coli Survival in Estuarine Bank Sediments

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Inoculum Development
2.2. Collection and Processing of Bank Sediment
2.3. Experimental Matrix
2.4. Experimental Procedure
2.5. Collection and Processing of Samples
2.6. Enumeration of E. coli and S. Typhimurium and Calculation of Die-Off Rates
3. Results and Discussion
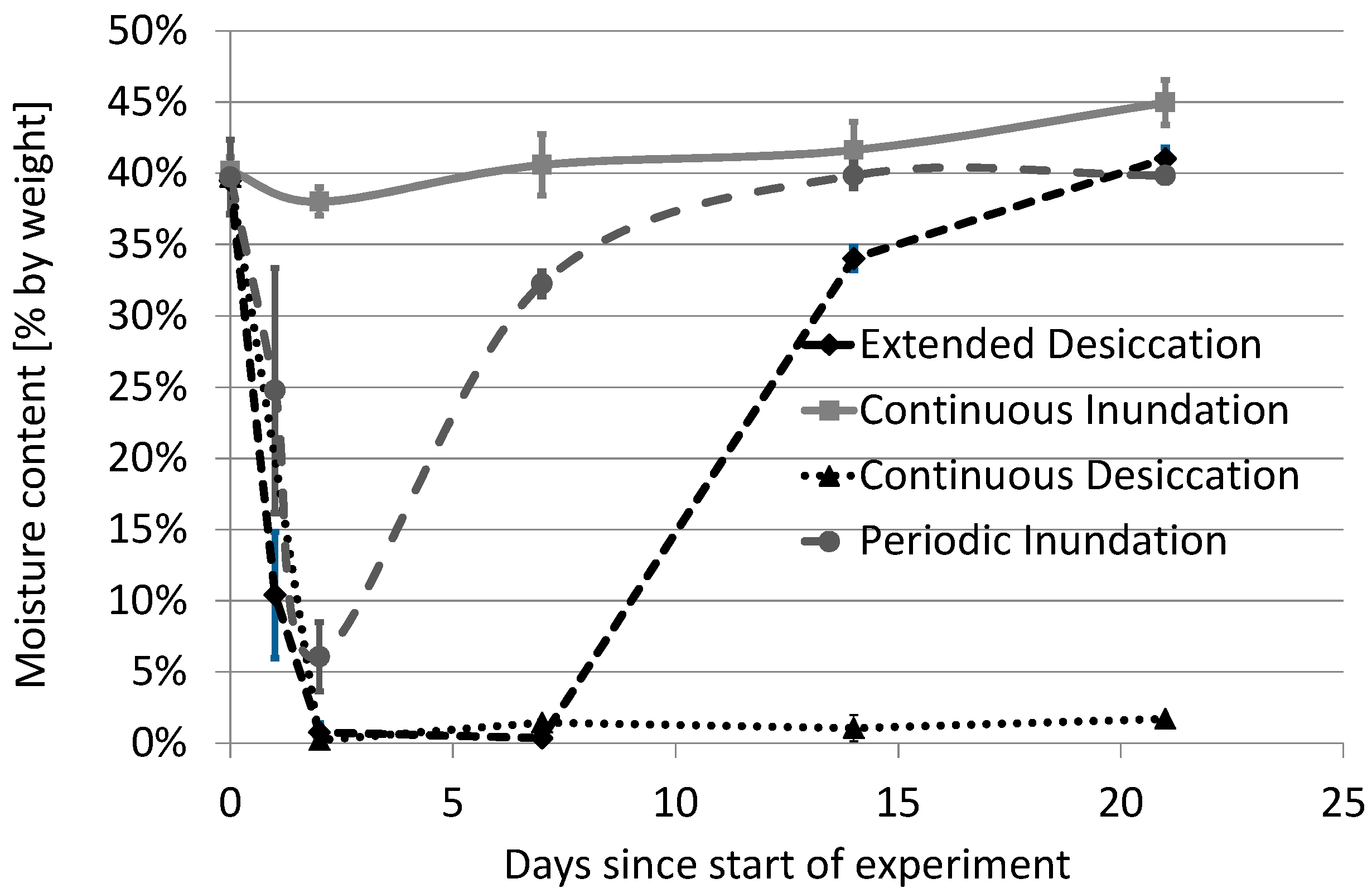
3.1. The Impact of Experimental Design
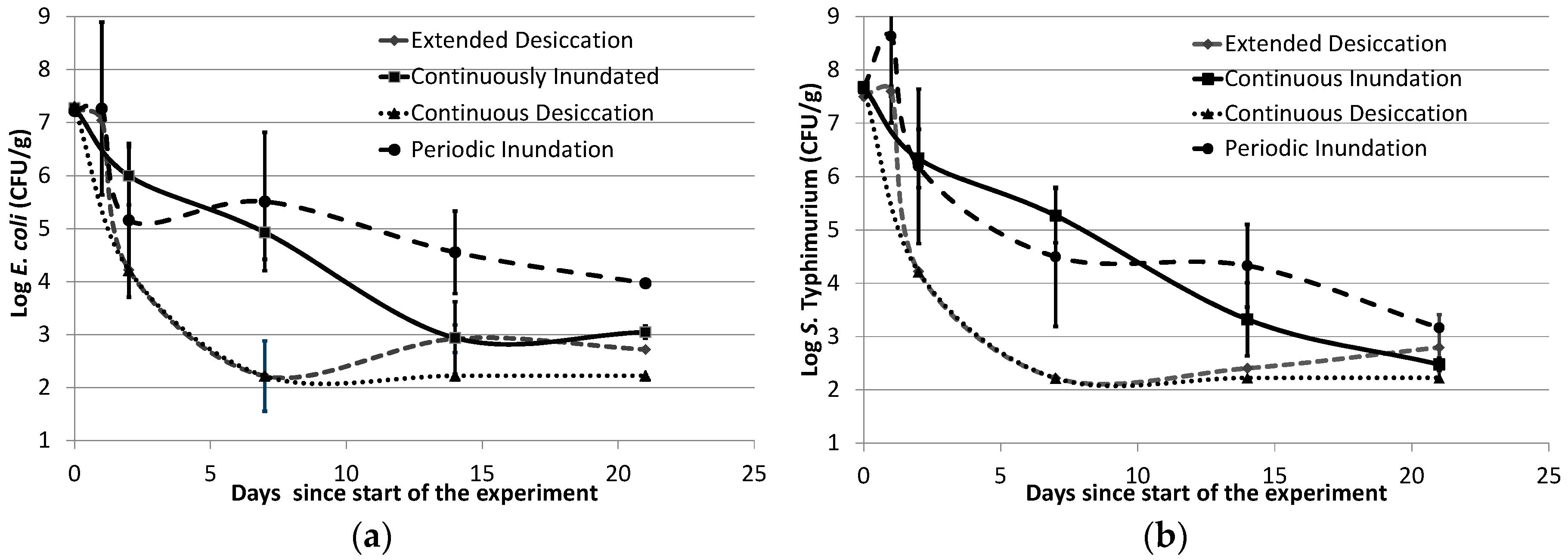
3.2. Survival of E. coli and S. Typhimurium
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Effect of Autoclaving

Appendix B
Changes in Sediments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time during Experiment | E. coli (cfu/g dw) | S. Typhimurium (cfu/g dw) | TP (mg/g dw) | TN (mg/g dw) |
|---|---|---|---|---|
| Original bank conditions—not inoculated, not autoclaved | ||||
| Day = 0 | 540 | Nil | 0.21 * | 0.77 * |
| Autoclaved, not inoculated | ||||
| Day = 0 | Nil | Nil | 0.18 * | 0.78 * |
| Day = 21 (PI) | Nil | Nil | 0.4 (12) | 1.4 (5) |
| Autoclaved, spiked with E. coli, S. Typhimurim and mixed culture | ||||
| Day = 0 | ** | ** | 0.20 * | 0.77 * |
| Day = 21 (PI) | ** | ** | 0.4 (2) | 1.3 (1) |
| Autoclaved, spiked with E. coli, S. Typhimurim | ||||
| Day = 0 | *** | *** | 0.21 * | 0.81 * |
| Day = 21 (ED) | *** | *** | 0.4 (7) | 1.4 (3) |
| Day = 21 (CI) | *** | *** | 0.4 (12) | 1.3 (8) |
| Day = 21 (CD) | *** | *** | 0.4 (4) | 1.4 (0) |
| Day = 21 (PI) | *** | *** | 0.4 (3) | 1.4 (3) |
| Time during Experiment | TP (mg/L) | TN (mg/L) | pH | EC (mS/cm) | Turbidity (NTU) |
|---|---|---|---|---|---|
| Fresh Inflow | |||||
| 0.13 (37) | 0.9 (4) | 6.6 (4) | 0 (17) | 2.14 * | |
| Extended Desiccation | |||||
| Day = 7 | 7.4 (2) | 0.5 (8) | |||
| Day = 14 | 5.8 (6) | 1.7 (6) | |||
| Day = 21 | 0.25 (48) | 6.4 (48) | 5.7 (7) | 1 (38) | 8.1 (62) |
| Non-Inoculated | |||||
| Day = 7 | 0.27 (45) | 13 (22) | 7.2 (1) | 2.5 (20) | |
| Day = 14 | 0.17 (42) | 4.2 (67) | 6.1 (1) | 0.3 (6) | |
| Day = 21 | 5.8 (10) | 2.7 (46) | 4.4 (37) | ||
| Continuously Inundated | |||||
| Day = 7 | 7.5 (0) | 2.4 (10) | |||
| Day = 14 | 6.7 (2) | 0.7 (1) | |||
| Day = 21 | 0.37 (25) | 3.6 (19) | 7.1 (0) | 0.4 (4) | 12.3 (44) |
| Periodically Inundated | |||||
| Day = 7 | 0.27 (19) | 10.83 (19) | 6.7 (3) | 1.9 (23) | |
| Day = 14 | 0.2 (12) | 5.8 (9) | 6 (9) | 0.7 (1) | |
| Day = 21 | 0.25 (13) | 12.2 (43) | 5.8 (11) | 1.7 (36) | 4.4 (30) |
| Nutritionally Augmented | |||||
| Day = 7 | 0.25 (43) | 10.33 (49) | 0.1 (1) | 1.8 (46) | |
| Day = 14 | 0.23 (11) | 7.27 (12) | 6.4 (9) | 0.9 (23) | |
| Day = 21 | 6.3 (14) | 1.9 (7) | 3.7 (27) | ||
References
- Ross, D.A. Introduction to Oceanography; Harper Collins College Publishers: New York, NY, USA, 1995; ISBN 978-0-673-46938-0. [Google Scholar]
- Mallin, M.A.; Williams, K.E.; Esham, E.C.; Lowe, R.P. Effect of human development on bacteriological water quality in coastal watersheds. Ecol. Appl. 2000, 10, 1047–1056. [Google Scholar] [CrossRef]
- Catalao-Dionisio, L.P.; Joao, M.; Ferreiro, V.S.; Fidalgo, M.L.; Garcia-Rosado, M.E.; Borrego, J.J. Occurrence of Salmonella spp. in estuarine and coastal waters of Portugal. Antonie Van Leeuwenhoek 2000, 78, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Samhan, F.A.; Kronlein, M.R.; Fakher, U.; Kronlein, C.; Stedtfeld, R.D.; Hashsham, S.A. Detection and occurrence of indicator organisms and pathogens. Water Environ. Res. 2009, 81, 959–980. [Google Scholar] [CrossRef] [PubMed]
- Henry, R.; Schang, C.; Chandrasena, G.I.; Deletic, A.; Edmunds, M.; Jovanovic, D.; Kolotelo, P.; Schmidt, J.; Williamson, R.; McCarthy, D. Environmental monitoring of waterborne Campylobacter: Evaluation of the Australian standard and a hybrid extraction-free MPN-PCR method. Front. Microbiol. 2015, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Edberg, S.C.; Allen, M.J.; Smith, D.B.; Kriz, N.J. Enumeration of total coliforms and Escherichia coli from source water by the defined substrate technology. Appl. Environ. Microbiol. 1990, 56, 366–369. [Google Scholar] [PubMed]
- Odonkor, S.T.; Ampofo, J.K. Escherichia coli as an indicator of bacteriological quality of water: An overview. Microbiol. Res. 2013, 4, e2. [Google Scholar] [CrossRef]
- Australian Government. Guidelines for Managing Risks in Recreational Water; NHMRC: Canberra, Australia, 2008.
- Schang, C.; Lintern, A.; Cook, P.L.M.; Osborne, C.A.; McKinley, A.; Schmidt, J.; Coleman, R.; Rooney, G.; Henry, R.; Deletic, A.; et al. Presence and survival of culturable Campylobacter spp. and Escherichia coli in temperate urban estuaries. Sci. Total Environ. 2016, 569–570, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Hood, M.A.; Ness, G.E. Survival of Vibrio cholerae and Escherichia coli in estuarine waters and sediments. Appl. Environ. Microbiol. 1982, 43, 578–584. [Google Scholar] [PubMed]
- Venkatramanan, S.; Ramkumar, T.; Anithamary, I. Distribution of grain size, clay mineralogy and organic matter of surface sediments from Tirumalairajanar Estuary, Tamilnadu, east coast of India. Arab. J. Geosci. 2013, 6, 1371–1380. [Google Scholar] [CrossRef]
- Solo-Gabriele, H.M.; Wolfert, M.A.; Desmarais, T.R.; Palmer, C.J. Sources of Escherichia coli in a Coastal Subtropical Environment. Appl. Environ. Microbiol. 2000, 66, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Desmarais, T.R.; Solo-Gabriele, H.M.; Palmer, C.J. Influence of soil on fecal indicator organisms in a tidally influenced subtropical environment. Appl. Environ. Microbiol. 2002, 68, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- US-EPA. Fact Sheet: Final Third Drinking Water Contaminant Candidate List (CCL 3); Office of Water: Washington, DC, USA, 2009.
- OzFoodNet Working Group. Burden and Causes of Foodborne Disease in Australia: Annual Report of the OzFoodNet Network, 2005; Office of Health Protection, Australian Government Department of Health and Ageing: Canberra, Australia, 2006; Volume 30.
- Chandran, A.; Mohamed Hatha, A.A. Relative survival of Escherichia coli and Salmonella typhimurium in a tropical estuary. Water Res. 2005, 39, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Goto, D.; Yan, T. Effects of autochthonous microbial community on the die-off of fecal indicators in tropical beach sand. FEMS Microbiol. Ecol. 2010, 74, 214–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanjugi, P.; Harwood, V.J. The influence of predation and competition on the survival of commensal and pathogenic fecal bacteria in aquatic habitats. Environ. Microbiol. 2013, 15, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Vdović, N.; Obhođaš, J.; Pikelj, K. Revisiting the particle-size distribution of soils: Comparison of different methods and sample pre-treatments. Eur. J. Soil Sci. 2010, 61, 854–864. [Google Scholar] [CrossRef]
- McCarthy, D.T. Modelling microorganisms in urban stormwater. Ph.D. Thesis, Department of Civil Engineering, Monash University, Melbourne, Australia, 2008. [Google Scholar]
- Bratieres, K.; Fletcher, T.D.; Deletic, A.; Zinger, Y. Nutrient and sediment removal by stormwater biofilters: A large-scale design optimisation study. Water Res. 2008, 42, 3930–3940. [Google Scholar] [CrossRef] [PubMed]
- Australia Standard. AS 1289.2.1.1: Methods of Testing Soils for Engineering Purposes—Soil Moisture Content Tests—Determination of the Moisture Content of a Soil-Oven Drying Method (Standard Method); SAI Global: Sydney, Australia, 2005. [Google Scholar]
- Roszak, D.B.; Grimes, D.J.; Colwell, R.R. Viable but nonrecoverable stage of Salmonella enteritidis in aquatic systems. Can. J. Microbiol. 1984, 30, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Bogosian, G.; Sammons, L.E.; Morris, P.J.; O’Neil, J.P.; Heitkamp, M.A.; Weber, D.B. Death of the Escherichia coli K-12 strain W3110 in soil and water. Appl. Environ. Microbiol. 1996, 62, 4114–4120. [Google Scholar] [PubMed]
- Nguyen, H.T.M.; Le, Q.T.P.; Garnier, J.; Janeau, J.L.; Rochelle-Newall, E. Seasonal variability of faecal indicator bacteria numbers and die-off rates in the Red River basin, North Viet Nam. Sci. Rep. 2016, 6, 21644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitcomb, B.W.; Schisterman, E.F. Assays with lower detection limits: Implications for epidemiological investigations. Paediatr. Perinat. Epidemiol. 2008, 22, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Mallmann, W.L.; Litsky, W. Survival of Selected Enteric Organisms in Various Types of Soil. Am. J. Public Health Nations Health 1951, 41, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Pommepuy, M.; Guillaud, J.F.; Dupray, E.; Derrien, A.; Le Guyader, F.; Cormier, M. Enteric bacteria survival factors. Water Sci. Technol. 1992, 25, 93–103. [Google Scholar] [CrossRef]
- Pote, J.; Haller, L.; Kottelat, R.; Sastre, V.; Arpagaus, P.; Wildi, W. Persistence and growth of faecal culturable bacterial indicators in water column and sediments of Vidy Bay, Lake Geneva, Switzerland. J. Environ. Sci. 2009, 21, 62–69. [Google Scholar] [CrossRef]
- Garzio-Hadzick, A.; Shelton, D.R.; Hill, R.L.; Pachepsky, Y.A.; Guber, A.K.; Rowland, R. Survival of manure-borne E. coli in streambed sediment: Effects of temperature and sediment properties. Water Res. 2010, 44, 2753–2762. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.A.; Gunnison, D.; Lanza, G.R. Survival of pathogenic bacteria in various freshwater sediments. Appl. Environ. Microbiol. 1987, 53, 633–638. [Google Scholar] [PubMed]
- Malham, S.K.; Rajko-Nenow, P.; Howlett, E.; Tuson, K.E.; Perkins, T.L.; Pallett, D.W.; Wang, H.; Jago, C.F.; Jones, D.L.; McDonald, J.E. The interaction of human microbial pathogens, particulate material and nutrients in estuarine environments and their impacts on recreational and shellfish waters. Environ. Sci. Process. Impacts 2014, 16, 2145–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, T.L.; Clements, K.; Baas, J.H.; Jago, C.F.; Jones, D.L.; Malham, S.K.; McDonald, J.E. Sediment composition influences spatial variation in the abundance of human pathogen indicator bacteria within an estuarine environment. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Gerba, C.P.; McLeod, J.S. Effects of sediments on the survival of Escherichia coli in marine waters. Appl. Environ. Microbiol. 1976, 32, 114–120. [Google Scholar] [PubMed]
- Sinton, L.W.; Davies-Colley, R.J.; Bell, R.G. Inactivation of enterococci and fecal coliforms from sewage and meatworks effluents in seawater chambers. Appl. Environ. Microbiol. 1994, 60, 2040–2048. [Google Scholar] [PubMed]
- Davies-Colley, R.J.; Donnison, A.M.; Speed, D.J.; Ross, C.M.; Nagels, J.W. Inactivation of faecal indicator micro-organisms in waste stabilisation ponds: Interactions of environmental factors with sunlight. Water Res. 1999, 33, 1220–1230. [Google Scholar] [CrossRef]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the Environment. Microbiol. Mol. Biol. Rev. 2012, 76, 685–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, A.F.; Pramer, D. An evaluation of factors affecting the survival of Escherichia coli in sea water. II. Salinity, pH, and nutrients. Appl. Microbiol. 1960, 8, 247–250. [Google Scholar] [PubMed]
- Korajkic, A.; McMinn, B.R.; Shanks, O.C.; Sivaganesan, M.; Fout, G.S.; Ashbolt, N.J. Biotic Interactions and Sunlight Affect Persistence of Fecal Indicator Bacteria and Microbial Source Tracking Genetic Markers in the Upper Mississippi River. Appl. Environ. Microbiol. 2014, 80, 3952–3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, P.; Morris, N.Z.; Wellington, E.M.H. Quantitative molecular detection of Salmonella typhimurium in soil and demonstration of persistence of an active but non-culturable population. FEMS Microbiol. Ecol. 1998, 27, 351–363. [Google Scholar] [CrossRef]
- Chen, Z.; Diao, J.; Dharmasena, M.; Ionita, C.; Jiang, X.; Rieck, J. Thermal Inactivation of Desiccation-Adapted Salmonella spp. in Aged Chicken Litter. Appl. Environ. Microbiol. 2013, 79, 7013–7020. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Hill, C. Stress adaptation in foodborne pathogens. Annu. Rev. Food Sci. Technol. 2015, 6, 191–210. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.K.; Gottschall, N.; Wilkes, G.; Grégoire, D.S.; Topp, E.; Pintar, K.D.M.; Sunohara, M.; Marti, R.; Lapen, D.R. Rainfall-induced runoff from exposed streambed sediments: An important source of water pollution. J. Environ. Qual. 2015, 44, 236–247. [Google Scholar] [CrossRef] [PubMed]
- An, Y.J.; Kampbell, D.H.; Breidenbach, G.P. Escherichia coli and total coliforms in water and sediments at lake marinas. Environ. Pollut. 2002, 120, 771–778. [Google Scholar] [CrossRef]
- Grimes, D.J. Release of Sediment-Bound Fecal Coliforms by Dredging. Appl. Microbiol. 1975, 29, 109–111. [Google Scholar] [PubMed]
- D’Aoust, J.Y.; Maurer, J. Salmonella species. In Food Microbiology: Fundamentals and Frontiers; Doyle, M.P., Beuchat, L.R., Eds.; ASM Press: Washington, DC, USA, 1997. [Google Scholar]
- Stephenson, G.R.; Rychert, R.C. Bottom Sediment: A Reservoir of Escherichia coli in Rangeland Streams. J. Range Manag. 1982, 35, 119–123. [Google Scholar] [CrossRef]
- Chudoba, E.A.; Mallin, M.A.; Cahoon, L.B.; Skrabal, S.A. Stimulation of fecal bacteria in ambient waters by experimental inputs of organic and inorganic phosphorus. Water Res. 2013, 47, 3455–3466. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, C.M.; Coote, B.G.; Ashbolt, N.J.; Stevenson, I.M. Relationships between indicators, pathogens and water quality in an estuarine system. Water Res. 1996, 30, 2045–2054. [Google Scholar] [CrossRef]
- Tate, R.L. Cultural and Environmental Factors Affecting the Longevity of Escherichia coli in Histosols. Appl. Environ. Microbiol. 1978, 35, 925–929. [Google Scholar] [PubMed]
- Zibilske, L.M.; Weaver, R.W. Effect of Environmental Factors on Survival of Salmonella typhimurium in Soil1. J. Environ. Qual. 1978, 7, 593–597. [Google Scholar] [CrossRef]
- Fujioka, R.S.; Hashimoto, H.H.; Siwak, E.B.; Young, R.H. Effect of sunlight on survival of indicator bacteria in seawater. Appl. Environ. Microbiol. 1981, 41, 690–696. [Google Scholar] [PubMed]
- Crane, S.R.; Moore, J.A. Modeling enteric bacterial die-off: A review. Water Air Soil Pollut. 1986, 27, 411–439. [Google Scholar] [CrossRef]
- Semenov, A.V.; van Overbeek, L.; Termorshuizen, A.J.; van Bruggen, A.H. Influence of aerobic and anaerobic conditions on survival of Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in Luria-Bertani broth, farm-yard manure and slurry. J. Environ. Manag. 2011, 92, 780–787. [Google Scholar] [CrossRef] [PubMed]



| Configuration | No. of Replicates | Inoculation with E. coli and S. Typhimurium | Extent of Inundation |
|---|---|---|---|
| Continuous desiccation (CD) | 3 | Yes | Never |
| Extended desiccation (ED) | 3 | Yes | Starts on Day 7; periodic (cycles of 12.4 h) |
| Periodic inundation (PI) | 3 | Yes | Periodic (cycles of 12.4 h) |
| Continuous inundation (CI) | 3 | Yes | Continuous |
| Non-inoculated (NI) | 2 | No | Periodic (cycles of 12.4 h) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddiqee, M.H.; Henry, R.; Coulthard, R.; Schang, C.; Williamson, R.; Coleman, R.; Rooney, G.; Deletic, A.; McCarthy, D. Salmonella enterica Serovar Typhimurium and Escherichia coli Survival in Estuarine Bank Sediments. Int. J. Environ. Res. Public Health 2018, 15, 2597. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112597
Siddiqee MH, Henry R, Coulthard R, Schang C, Williamson R, Coleman R, Rooney G, Deletic A, McCarthy D. Salmonella enterica Serovar Typhimurium and Escherichia coli Survival in Estuarine Bank Sediments. International Journal of Environmental Research and Public Health. 2018; 15(11):2597. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112597
Chicago/Turabian StyleSiddiqee, Mahbubul H., Rebekah Henry, Rebecca Coulthard, Christelle Schang, Richard Williamson, Rhys Coleman, Graham Rooney, Ana Deletic, and David McCarthy. 2018. "Salmonella enterica Serovar Typhimurium and Escherichia coli Survival in Estuarine Bank Sediments" International Journal of Environmental Research and Public Health 15, no. 11: 2597. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112597