Fluoride Exposure Induces Inhibition of Sodium/Iodide Symporter (NIS) Contributing to Impaired Iodine Absorption and Iodine Deficiency: Molecular Mechanisms of Inhibition and Implications for Public Health

{kind=link}
Abstract
:1. Introduction
1.1. Dietary Sources of Iodine
1.2. Metabolism of Iodine
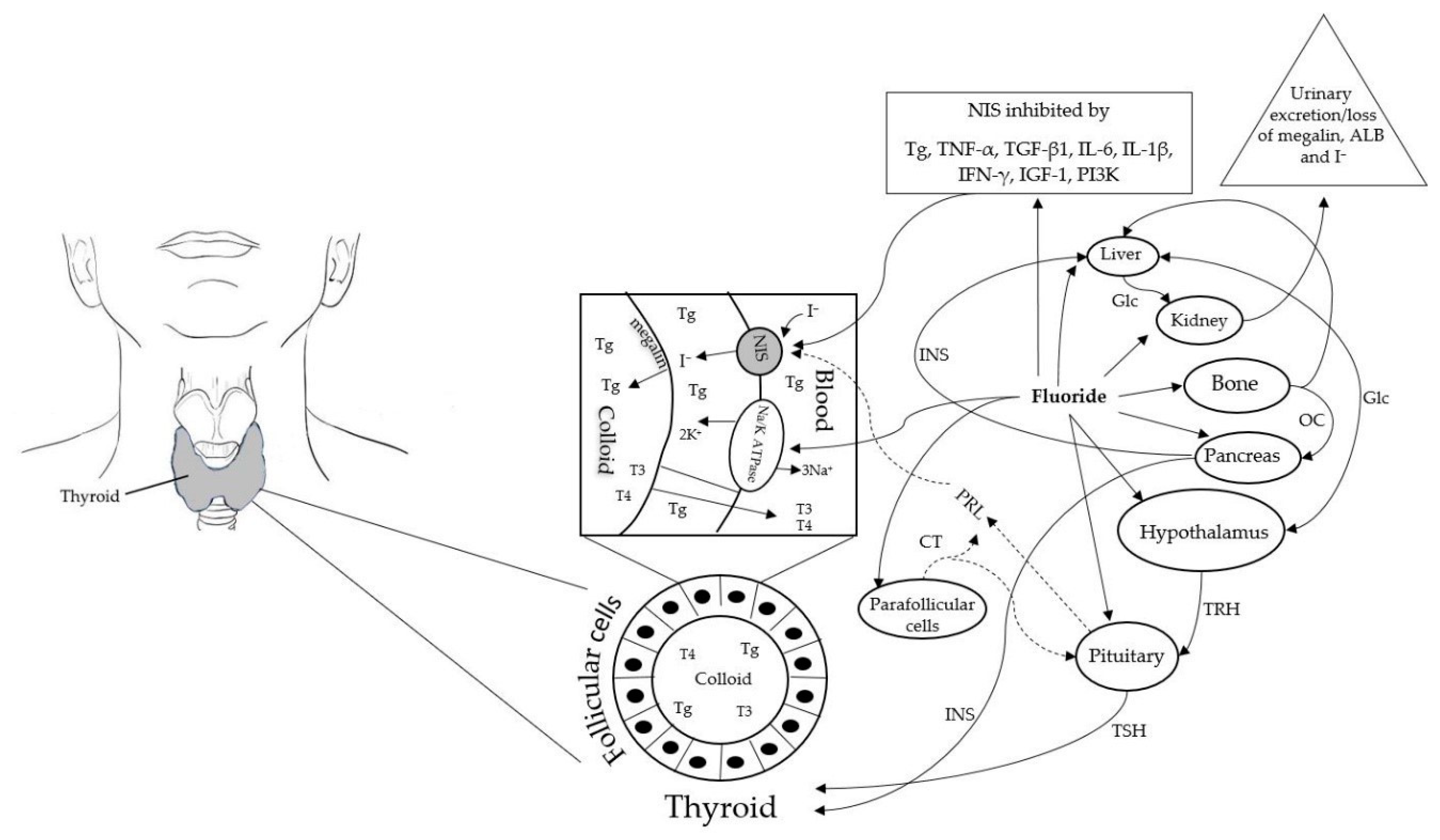
2. Molecular Mechanisms of Fluoride Inhibition of Iodine Homeostasis
2.1. Molecular Mechanisms of F Inhibition of Na+/K+-ATPase Activity
2.2. Deciphering the Molecular Mechanisms of F Inhibition of NIS Expression and Activity
3. Discussion
4. Additional Perspectives
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zimmermann, M.B.; Jooste, P.L.; Pandav, C.S. Iodine-deficiency disorders. Lancet 2008, 372, 1251–1262. [Google Scholar] [CrossRef]
- Underwood, E.J. Trace Elements in Human and Animal Nutrition, 4th ed.; Academic Press: New York, NY, USA, 1977; p. 545. ISBN 0-12-709065-7. [Google Scholar]
- Hetzel, B.S. SOS for a Billion—The Nature and Magnitude of Iodine Deficiency Disorders, 2nd ed.; Hetzel, B.S., Pandav, C.V., Eds.; Oxford University Press: New Delhi, India, 1997; pp. 1–29. [Google Scholar]
- De Benoist, B.; McLean, E.; Andersson, M.; Rogers, L. Iodine Deficiency in 2007: Global Progress since 2003. Food Nutr. Bull. 2008, 29, 195–202. [Google Scholar] [CrossRef]
- WHO/UNICEF/International Council for the Control of Iodine Deficiency Disorders. Assessment of Iodine Deficiency Disorders and Monitoring Their Elimination: A Guide for Programme Managers, 3rd ed.; World Health Organization: Geneva, Switzerland, 2007; Available online: http://apps.who.int/iris/bitstream/handle/10665/43781/9789241595827_eng.pdf (accessed on 19 January 2019).
- Iodine Status Worldwide. WHO Global Database on Iodine Deficiency; World Health Organization: Geneva, Switzerland, 2004; Available online: https://apps.who.int/iris/handle/10665/43010 (accessed on 19 January 2019).
- Iodine Global Network. Global Iodine Nutrition Scorecard 2015; Iodine Global Network: Zurich, Switzerland, 2015; Available online: http://ign.org/cm_data/Scorecard_2015_August_26_new.pdf (accessed on 19 January 2019).
- Pearce, E.N.; Lazarus, J.H.; Moreno-Reyes, R.; Zimmermann, M.B. Consequences of iodine deficiency and excess in pregnant women: An overview of current knowns and unknowns. Am. J. Clin. Nutr. 2016, 104 (Suppl. 3), 918S–923S. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Review of Fluoride in Drinking Water, U.S.; The National Academic Press: Washington, DC, USA, 2006. [Google Scholar]
- Li, Y.; Zhang, J.; Li, Z. Prevention of iodine deficiency in high fluoride areas in Tianjin City, China. Fluoride 1998, 31, S18. [Google Scholar]
- Scientific Committee on Food. Opinion of the Scientific Committee on Food on the Tolerable Upper Intake Level of Iodine. SCF/CS/NUT/UPPLEV/26 Final 2002 October 7. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/sci-com_scf_out146_en.pdf (accessed on 19 January 2019).
- Susheela, A.K.; Bhatnagar, M.; Vig, K.; Mondal, N.K. Excess fluoride ingestion and thyroid hormone derangements in children living in Delhi, India. Fluoride 2005, 38, 151–161. [Google Scholar]
- Singla, S.; Shashi, A. Thyroid peroxidase activity as toxicity target for fluoride in patients with thyroid dysfunction. Curr. Res. Microbiol. Biotechnol. 2013, 1, 53–57. [Google Scholar]
- Malin, A.J.; Riddell, J.; McCague, H.; Till, C. Fluoride exposure and thyroid function among adults living in Canada: Effect modification by iodine status. Environ. Int. 2018, 121, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Verma, K.G.; Verma, P.; Sidhu, G.K.; Sachdeva, S. A comparative study of fluoride ingestion levels, serum thyroid hormone & TSH level derangements, dental fluorosis status among school children from endemic and non-endemic fluorosis areas. SpringerPlus 2014, 3, 7. [Google Scholar] [PubMed]
- Yang, Y.; Wang, X.; Guo, X. Effects of high iodine and high fluorine on children’s intelligence and the metabolism of iodine and fluorine. Chin. J. Epidemiol. 1994, 15, 296–298. [Google Scholar]
- Xue, G.; Li-Ju, D.; Yue, B.A. Influence of high fluoride concentration in drinking water on the function of children’s Thyroid. Henan J. Prev. Med. 2009, 03. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-HNYF200903004.htm (accessed on 19 January 2019).
- Mikhailets, N.D.; Balabolkin, M.I.; Rakitin, V.A.; Danilov, I.P. Thyroid function during prolonged exposure to fluorides. Probl. Endocrinol. 1996, 42, 6–9. [Google Scholar]
- Iodine Status on the Island of Ireland; Safefood Ireland: Cork, Ireland, 2018; p. 7. Available online: https://www.safefood.eu/SafeFood/media/SafeFoodLibrary/Documents/Publications/Research%20Reports/Iodine-Status-on-the-island-of-Ireland-23032018.pdf (accessed on 19 January 2019).
- O’Sullivan, V.; O’Connell, B.C. Water fluoridation, dentition status and bone health of older people in Ireland. Community Dent. Oral Epidemiol. 2015, 43, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.B.; Aeberli, I.; Melse-Boonstra, A.; Grimci, L.; Bridson, J.; Chaouki, N.; Mbhenyane, X.; Jooste, P.L. Iodine treatment in children with subclinical hypothyroidism due to chronic iodine deficiency decreases thyrotropin and C-peptide concentrations and improves the lipid profile. Thyroid 2009, 19, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Hollowell, J.G.; Staehling, N.W.; Hannon, W.H.; Flanders, D.W.; Gunter, E.W.; Maberly, G.F.; Braverman, L.E.; Pino, S.; Miller, D.T.; Garbe, P.L.; et al. Iodine nutrition in the United States. Trends and public health implications: Iodine excretion data from National Health and Nutrition Examination Surveys I and III (1971–1974 and 1988–1994). J. Clin. Endocrinol. Metab. 1998, 83, 3401–3408. [Google Scholar]
- Li, M.; Ma, G.; Boyages, S.; Eastman, C. Re-emergence of iodine deficiency in Australia. Asia Pac. J. Clin. Nutr. 2001, 10, 200–203. [Google Scholar] [CrossRef]
- Thomson, C.D.; Packer, M.A.; Butler, J.A.; Duffield, A.J.; O’Donaghue, K.L.; Whanger, P.D. Urinary selenium and iodine during pregnancy and lactation. J. Trace Elem. Med. Biol. 2001, 14, 210–217. [Google Scholar] [CrossRef]
- Cooper, G.J.S.; Croxson, M.S.; Ibbertson, H.K. Iodine intake in an urban environment: A study of urine iodide excretion in Auckland. N. Z. Med. J. 1984, 97, 142–145. [Google Scholar]
- Simpson, E.O.; Thaler, B.I.; Paulin, J.M.; Phelan, E.L.; Cooper, G.J.S. Iodide excretion in a salt-restriction trial. N. Z. Med. J. 1984, 97, 890–893. [Google Scholar]
- Smyth, P.; Burns, R.; Casey, M.; Mullan, K.; O’Herlihy, C.; O’Dowd, C. Iodine Status over Two Decades: Influence of Seaweed Exposure. Ir. Med. J. 2016, 109, 421. [Google Scholar] [PubMed]
- Peckham, S.; Lowery, D.; Spencer, S. Are fluoride levels in drinking water associated with hypothyroidism prevalence in England? A large observational study of GP practice data and fluoride levels in drinking water. J. Epidemiol. Community Health 2015, 69, 619–624. [Google Scholar] [CrossRef]
- Kheradpisheh, Z.; Mirzaei, M.; Mahvi, A.H.; Mokhtari, M.; Azizi, R.; Fallahzadeh, H.; Ehrampoush, M.H. Impact of Drinking Water Fluoride on Human Thyroid Hormones: A Case-Control Study. Sci. Rep. 2018, 8, 2674. [Google Scholar] [CrossRef]
- Trabelsi, M.; Guermazi, F.; Zeghal, N. Effect of fluoride on thyroid function and cerebellar development in mice. Fluoride 2001, 34, 165–173. [Google Scholar]
- Bouaziz, H.; Soussia, L.; Guermazi, F.; Zeghal, N. Fluoride-induced thyroid proliferative changes and their reversal in female mice and their pups. Fluoride 2005, 38, 185–192. [Google Scholar]
- Bouaziz, H.; Ammar, E.; Ghorbel, H.; Ketata, S.; Jamoussi, K.; Ayadi, F.; Guermazi, F.; Zeghal, N. Effect of fluoride ingested by lactating mice on the thyroid function and bone maturation of their suckling pups. Fluoride 2004, 37, 133–142. [Google Scholar]
- Ahmed, O.M.; Abd El-Tawab, S.M.; Ahmed, R.G. Effects of experimentally induced maternal hypothyroidism and hyperthyroidism on the development of rat offspring: I. The development of the thyroid hormones-neurotransmitters and adenosinergic system interactions. Int. J. Dev. Neurosci. 2010, 28, 437–454. [Google Scholar] [CrossRef]
- Ren, D.; Li, K.; Liu, D. A study of the intellectual ability of 8–14 year-old children in high fluoride, low iodine areas. Chin. J. Control Endem. Dis. 1989, 4, 251. [Google Scholar]
- Lin, F.F.; Aihati, H.T.; Zhao, H.X.; Jin, L.; Jiang, J.-Y.; Maimaiti, A. The relationship of a low-iodine and high-fluoride environment to subclinical cretinism in Xinjiang. Endem. Dis. Bull. 1991, 6, 62–67. [Google Scholar]
- Hong, F.; Cao, Y.; Wang, H. A study of fluorine effects on children’s intelligence development under different environments. Chin. Prim. Health Care 2001, 15, 56–57. [Google Scholar]
- Ge, Y.; Ning, H.; Wang, S.; Wang, J. Effects of high fluoride and low iodine on brain histopathology in offspring rats. Fluoride 2005, 38, 127–132. [Google Scholar]
- Wang, J.D.; Ge, Y.M.; Ning, H.M.; Wang, S.L. Effects of high fluoride and low iodine on biochemical indexes of the brain and learning-memory of offspring rats. Fluoride 2004, 37, 201–208. [Google Scholar]
- Zhao, W.; Zhu, H.; Yu, Z.; Aoki, K.; Misumi, J.; Zhang, X. Long term effects of various iodine and fluorine doses on the thyroid and fluorosis in mice. Endocr. Regul. 1998, 32, 63–70. [Google Scholar]
- Ge, Y.; Ning, H.; Wang, S.; Wang, J. DNA Damage in Thyroid Gland Cells of Rats Exposed to Long-Term Intake of High Fluoride and Low Iodine. Fluoride 2005, 38, 318–323. [Google Scholar]
- Cinar, A.; Selsuk, M. Effects of chronic fluorosis on thyroxine, triiodothyronine, and protein-bound iodine in cows. Fluoride 2005, 38, 65–68. [Google Scholar]
- Bildik, A.; Camas, H. The research of some specific liver enzyme activities and PBI values in the blood serums of sheep with fluorosis. Kafkas Univ. Fen Bil. Derg. 1996, 1, 16–23. [Google Scholar]
- Guan, Z.Z.; Zhuang, Z.J.; Yang, P.S.; Pan, S. Synergistic action of iodine-deficiency and fluorine intoxication on rat thyroid. Chin. Med. J. 1988, 101, 679–684. [Google Scholar] [PubMed]
- Kundel, M.; Thorenz, U.R.; Petersen, J.H.; Huang, R.J.; Bings, N.H.; Hoffmann, T. Application of mass spectrometric techniques for the trace analysis of short-lived iodine-containing volatiles emitted by seaweed. Anal. Bioanal. Chem. 2012, 402, 3345–3357. [Google Scholar] [CrossRef]
- Gilfedder, B.S.; Petre, M.; Biester, H. Iodine speciation in rain and snow: Implications for the atmospheric iodine sink. J. Geophys. Res. 2007, 112, D07301. [Google Scholar] [CrossRef]
- Baker, A.R. Marine Aerosol Iodine Chemistry: The Importance of Soluble Organic Iodine. Environ. Chem. 2005, 2, 295–298. [Google Scholar] [CrossRef]
- McFiggans, G.; Coe, H.; Burgess, R.; Allan, J.; Cubison, M.; Alfarra, M.R.; Saunders, R.; Saiz-Lopez, A.; Plane, J.M.C.; Wevill, D.J.; et al. Direct evidence for coastal iodine particles from Laminaria macroalgae–linkage to emissions of molecular iodine. Atmos. Chem. Phys. 2004, 4, 701–713. [Google Scholar] [CrossRef]
- Carperter, L.J. Iodine in the marine boundary layer. Chem. Rev. 2003, 103, 4953–4962. [Google Scholar] [CrossRef]
- O’Dowd, C.D.; Jimenez, J.L.; Bahreini, R.; Flagan, R.C.; Seinfeld, J.H.; Hämeri, K.; Pirjola, L.; Kulmala, M.; Jennings, S.G.; Hoffmann, T. Marine aerosol formation from biogenic iodine emissions. Nature 2002, 417, 632–636. [Google Scholar] [CrossRef] [PubMed]
- McFiggans, G.; Plane, J.M.C.; Allan, B.J.; Carpenter, L.J.; Coe, H.; O’Dowd, C. A modeling study of iodine chemistry in the marine boundary layer. J. Geophys. Res. 2000, 105, 14371–14385. [Google Scholar] [CrossRef]
- Duce, R.A.; Winchester, J.W.; Van Nahl, T.W. Iodine, bromine, and chlorine in winter aerosols and snow from Barrow, Alaska. Tellus 1966, 18, 238–248. [Google Scholar] [CrossRef]
- Rohner, F.; Zimmermann, M.; Jooste, P.; Pandav, C.; Caldwell, K.; Raghavan, R.; Raiten, D.J. Biomarkers of Nutrition for Development—Iodine Review. J. Nutr. 2014, 144, 1322S–1342S. [Google Scholar] [CrossRef] [PubMed]
- Smyth, P.P.; Burns, R.; Huang, R.J.; Hoffman, T.; Mullan, K.; Graham, U.; Seitz, K.; Platt, U.; O’Dowd, C. Does iodine gas released from seaweed contribute to dietary iodine intake? Environ. Geochem. Health 2011, 33, 389–397. [Google Scholar] [CrossRef]
- Vought, R.L.; London, W.T.; Brown, F.A. A note on atmospheric iodine and its absorption in man. J. Clin. Endocrinol. Metab. 1964, 24, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Nagataki, S. The average of dietary iodine intake due to the ingestion of seaweeds is 1.2 mg/day in Japan. Thyroid 2008, 18, 667–668. [Google Scholar] [CrossRef]
- O’Brien, B.; Gleeson, D.; Jordan, K. Iodine concentrations in milk. Ir. J. Agric. Food Res. 2013, 52, 209–216. [Google Scholar]
- Borucki Castro, S.I.; Berthiaume, R.; Laffey, P.; Fouquet, A.; Beraldin, F.; Robichaud, A.; Lacasse, P. Iodine concentration in milk sampled from Canadian farms. J. Food Prot. 2010, 73, 1658–1663. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.; Miller, P.; Baxter, M.; Appleton, G.; Crews, H.; Croasdale, M. Bromine and iodine in 1997 UK total diet study samples. J. Environ. Monit. 2001, 3, 361–365. [Google Scholar] [CrossRef]
- Pennington, J.A.T. Iodine Concentrations in US Milk: Variation Due to Time, Season, and Region. J. Dairy Sci. 1990, 73, 3421–3427. [Google Scholar] [CrossRef]
- Galton, D.M.; Petersson, L.G.; Erb, H.N. Milk iodine residues in herds practicing iodophor premilking teat disinfection. J. Dairy Sci. 1986, 69, 267–271. [Google Scholar] [CrossRef]
- Haldimann, M.; Alt, A.; Blanc, A.; Blondeau, K. Iodine content of food groups. J. Food Compos. Anal. 2005, 18, 461–471. [Google Scholar] [CrossRef]
- Pennington, J.A.T.; Schoen, S.A.; Salmon, G.D.; Young, B.; Johnson, R.D.; Marts, R.W. Composition of core foods in the U.S. food supply. J. Food Compos. Anal. 1995, 8, 171–217. [Google Scholar] [CrossRef]
- Lu, Y.L.; Wang, N.J.; Zhu, L.; Wang, G.X.; Wu, H.; Kuang, L.; Zhu, W.M. Investigation of iodine concentration in salt, water and soil along the coast of Zhejiang, China. J. Zhejiang Univ. Sci. B 2005, 6, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.M.; Laurberg, P.; Nohr, S.; Jorgensen, A.; Andersen, S. Iodine in drinking water varies by more than 100-fold in Denmark. Importance for iodine content of infant formulas. Eur. J. Endocrinol. 1999, 140, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, D.C. Iodine in the U.K. Environment with Particular Reference to Agriculture. J. Appl. Ecol. 1979, 16, 269–279. [Google Scholar] [CrossRef]
- Leung, A.M.; Braverman, L.E.; Pearce, E.N. History of U.S. iodine fortification and supplementation. Nutrients 2012, 4, 1740–1746. [Google Scholar] [CrossRef] [PubMed]
- Pesce, L.; Kopp, P. Iodide transport: Implications for health and disease. Int. J. Pediatr. Endocrinol. 2014, 2014, 8. [Google Scholar] [CrossRef]
- Nicola, J.P.; Basquin, C.; Portulano, C.; Reyna-Neyra, A.; Paroder, M.; Carrasco, N. The Na+/I− symporter mediates active iodide uptake in the intestine. Am. J. Physiol. Cell Physiol. 2009, 296, C654–C662. [Google Scholar] [CrossRef] [PubMed]
- Brown-Grant, K. Extrathyroidal iodide concentrating mechanisms. Physiol. Rev. 1961, 41, 189–213. [Google Scholar] [CrossRef]
- Ahad, F.; Ganie, S.A. Iodine, Iodine metabolism and Iodine deficiency disorders revisited. Indian J. Endocrinol. Metab. 2010, 14, 13–17. [Google Scholar] [PubMed]
- Caillou, B.; Troalen, F.; Baudin, E.; Talbot, M.; Filetti, S.; Schlumberger, M.; Bidart, J.M. Na+/I− symporter distribution in human thyroid tissues: An immunohistochemical study. J. Clin. Endocrinol. Metab. 1998, 83, 4102–4106. [Google Scholar] [CrossRef]
- Eskandari, S.; Loo, D.D.F.; Dai, G.; Levy, O.; Wright, E.M.; Carrasco, N. Thyroid Na+/I− symporter mechanism, stoichiometry, and specificity. J. Biol. Chem. 1997, 272, 27230–27238. [Google Scholar] [CrossRef]
- Jomaa, B. Thyroid Toxicogenomics: A Multi-Organ Paradigm. In Toxicogenomics-Based Cellular Models; Kleinjans, J., Ed.; Elsevier Academic Press: Cambridge, MA, USA, 2014; pp. 159–189. [Google Scholar]
- Dohán, O.; De la Vieja, A.; Paroder, V.; Riedel, C.; Artani, M.; Reed, M.; Ginter, C.S.; Carrasco, N. The sodium/iodide Symporter (NIS): Characterization, regulation, and medical significance. Endocr. Rev. 2003, 24, 48–77. [Google Scholar] [CrossRef] [PubMed]
- Hingorani, M.; Spitzweg, C.; Vassaux, G.; Newbold, K.; Melcher, A.; Pandha, H.; Vile, R.; Harrington, K. The Biology of the Sodium Iodide Symporter and its Potential for Targeted Gene Delivery. Curr. Cancer Drug Targets 2010, 10, 242–267. [Google Scholar] [CrossRef] [PubMed]
- Dai, G.; Levy, O.; Carrasco, N. Cloning and characterization of the thyroid iodide transporter. Nature 1996, 379, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Spitzweg, C.; Joba, W.; Eisenmenger, W.; Heufelder, A.E. Analysis of human sodium iodide symporter gene expression in extrathyroidal tissues and cloning of its complimentary deoxyribonucleic acids from salivary gland, mammary gland and gastric mucosa. J. Clin. Endocrinol. Metab. 1998, 83, 1746–1751. [Google Scholar] [CrossRef] [PubMed]
- Di Jeso, B.; Arvan, P. Thyroglobulin from Molecular and Cellular Biology to Clinical Endocrinology. Endocr. Rev. 2015, 37, 2–36. [Google Scholar] [CrossRef]
- Lisi, S.; Pinchera, A.; McCluskey, R.T.; Willnow, T.E.; Refetoff, S.; Marcocci, C.; Vitti, P.; Menconi, F.; Grasso, L.; Luchetti, F.; et al. Preferential megalin-mediated transcytosis of low-hormonogenic thyroglobulin: A control mechanism for thyroid hormone release. Proc. Natl. Acad. Sci. USA 2003, 100, 14858–14863. [Google Scholar] [CrossRef]
- Marinò, M.; Pinchera, A.; McCluskey, R.T. Megalin in thyroid physiology and pathology. Thyroid 2001, 11, 47–56. [Google Scholar] [CrossRef]
- Marinò, M.; Zheng, G.; Chiovato, L.; Pinchera, A.; Brown, D.; Andrews, D.; McCluskey, R.T. Role of megalin (gp330) in transcytosis of thyroglobulin by thyroid cells. A novel function in the control of thyroid hormone release. J. Biol. Chem. 2000, 275, 7125–7137. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ishido, Y.; Hiroi, N.; Ishii, N.; Suzuki, K. The Emerging Roles of Thyroglobulin. Adv. Endocrinol. 2014, 2014, 189194. [Google Scholar] [CrossRef]
- Abrams, G.M.; Larsen, P.R. Triiodothyronine and thyroxine in the serum and thyroid glands of iodine-deficient rats. J. Clin. Investig. 1973, 52, 2522–2531. [Google Scholar] [CrossRef] [PubMed]
- De Vijlder, J.J. Primary congenital hypothyroidism: Defects in iodine pathways. Eur. J. Endocrinol. 2003, 149, 247–256. [Google Scholar] [CrossRef]
- Zimmermann, M.B. Iodine and iodine deficiency disorders. In Present Knowledge in Nutrition, 10th ed.; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 554–567. [Google Scholar]
- Zimmermann, M.B. Methods to assess iron and iodine status. Br. J. Nutr. 2008, 99 (Suppl. 3), S2–S9. [Google Scholar] [CrossRef]
- Alexander, W.D.; Harden, R.M.; Harrison, M.T.; Shimmins, J. Some aspects of the absorption and concentration of iodide by the alimentary tract in man. Proc. Nutr. Soc. 1967, 26, 62–66. [Google Scholar] [CrossRef] [PubMed]
- DeGroot, L.J. Kinetic analysis of iodine metabolism. J. Clin. Endocrinol. Metab. 1966, 26, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Stanbury, J.B.; Brownell, G.L.; Riggs, D.S.; Perinetti, H.; Itoiz, J.; Del Castillo, E.B. Endemic goiter. In The Adaptation of Man to Iodine Deficiency; Stanbury, J.B., Ed.; Harvard University Press: Cambridge, MA, USA, 1954; pp. 1–209. [Google Scholar]
- Wayne, E.J.; Koutras, D.A.; Alexander, W.D. Clinical Aspects of Iodine Metabolism; Blackwell Scientific: Oxford, UK, 1964. [Google Scholar]
- Zimmermann, M.B. Iodine deficiency. Endocr. Rev. 2009, 30, 376–408. [Google Scholar] [CrossRef]
- Glinoer, D. The regulation of thyroid function in pregnancy: Pathways of endocrine adaptation from physiology to pathology. Endocr. Rev. 1997, 18, 404–433. [Google Scholar] [CrossRef]
- Karam, G.A.; Hakimi, H.; Rezaeian, M.; Gafarzadeh, H.; Rashidinejad, H.; Khaksari, M. Thyroid function in mothers who gave birth to neonates with transient congenital hypothyroidism. Pak. J. Med. Sci. 2009, 25, 568–572. [Google Scholar]
- Azizi, F.; Smyth, P. Breastfeeding and maternal and infant iodine nutrition. Clin. Endocrinol. 2009, 70, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.L.; Luckyanov, N.; Bouville, A.; VanMiddlesworth, L.; Weinstock, R.M. Transfer of 131I into human breast milk and transfer coefficients for radiological dose assessments. Health Phys. 2002, 82, 796–806. [Google Scholar] [CrossRef]
- Waugh, D.T. Molecular Mechanisms of Fluoride inhibition of Na+, K+-ATPase activity: Implications for Public Health and Health Inequalities. Int. J. Environ. Res. Public Health 2019, in press. [Google Scholar]
- Pekary, A.E.; Levin, S.R.; Johnson, D.G.; Berg, L.; Hershman, J.M. Tumor necrosis factor-alfa (TNF-alfa) and transforming growth factor beta1 (TGF-beta1) inhibit the expression and activity of Na+/K+ ATPase in FRTL-5 rat thyroid cells. J. Interferon Cytokine Res. 1997, 4, 185–195. [Google Scholar] [CrossRef]
- Ajjan, R.A.; Watson, P.F.; Findlay, C.; Metcalfe, R.A.; Crisp, M.; Ludgate, M.; Weetman, A.P. The sodium iodide symporter gene and its regulation by cytokines found in autoimmunity. J. Endocrinol. 1998, 158, 351–358. [Google Scholar] [CrossRef]
- Pekary, A.E.; Hershman, J.M. Tumor necrosis factor, ceramide, transforming growth factor-beta1, and aging reduce Na+/I− sym-porter messenger ribonucleic acid levels in FRTL-5 cells. Endocrinology 1998, 139, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Popławska-Kita, A.; Telejko, B.; Siewko, K.; Kościuszko-Zdrodowska, M.; Wawrusewicz-Kurylonek, N.; Krętowski, A.; Hryniewicka, J.; Dzięcioł, J.; Bauer, W.; Milewski, R.; et al. Decreased Expression of Thyroglobulin and Sodium Iodide Symporter Genes in Hashimoto’s Thyroiditis. Int. J. Endocrinol. 2014, 2014, 690704. [Google Scholar] [CrossRef]
- Pang, X.P.; Park, M.; Hershman, J.M. Transforming growth factor-beta blocks protein kinase-A-mediated iodide transport and protein kinase-C-mediated DNA synthesis in FRTL-5 rat thyroid cells. Endocrinology 1992, 131, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, A.; Ikeda, M.; Endo, T.; Kogai, T.; Miyazaki, A.; Onaya, T. Transforming growth factor-beta1 suppresses thyrotropin-induced Na+/I− symporter messenger RNA and protein levels in FRTL-5 rat thyroid cells. Thyroid 1997, 7, 789–794. [Google Scholar] [CrossRef]
- Spitzweg, C.; Joba, W.; Morris, J.C.; Heufelder, A.E. Regulation of sodium iodide symporter gene expression in FRTL-5 rat thyroid cells. Thyroid 1999, 9, 821–830. [Google Scholar] [CrossRef]
- Caturegli, P.; Hejazi, M.; Suzuki, K.; Dohan, O.; Carrasco, N.; Kohn, L.D.; Rose, N.R. Hypothyroidism in transgenic mice expressing IFN-y in the thyroid. Proc. Natl. Acad. Sci. USA 2000, 97, 1719–1724. [Google Scholar] [CrossRef]
- Cass, L.A.; Meinkoth, J.L. Ras signaling through PI3K confers hormone-independent proliferation that is compatible with differentiation. Oncogene 2000, 19, 924–932. [Google Scholar] [CrossRef] [PubMed]
- García, B.; Santisteban, P. PI3K is involved in the IGF-I inhibition of TSH-induced sodium/iodide symporter gene expression. Mol. Endocrinol. 2002, 16, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Royaux, I.E.; Suzuki, K.; Mori, A.; Katoh, R.; Everett, L.A.; Kohn, L.D.; Green, E.D. Pendrin, the protein encoded by the Pendred syndrome gene (PDS)is an apical porter of iodide in the thyroid and is regulated by thyroglobulin. Endocrinology 2000, 141, 839–845. [Google Scholar] [CrossRef]
- Suzuki, K.; Lavaroni, S.; Mori, A.; Ohta, M.; Saito, J.; Pietrarelli, M.; Singer, D.S.; Kimura, S.; Katoh, R.; Kawaoi, A.; et al. Autoregulation of thyroid-specific gene transcription by thyroglobulin. Proc. Natl. Acad. Sci. USA 1998, 95, 8251–8256. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Mori, A.; Saito, J.; Moriyama, E.; Ullianich, L.; Kohn, L.D. Follicular thyroglobulin suppresses iodide uptake by suppressing expression of the sodium iodide symporter gene. Endocrinology 1999, 140, 5422–5430. [Google Scholar] [CrossRef]
- Suzuki, K.; Mori, A.; Lavaroni, S.; Ulianich, L.; Miyagi, E.; Saito, J.; Nakazato, M.; Pietrarelli, M.; Shafran, N.; Grassadonia, A.; et al. Thyroglobulin regulates follicular function and heterogeneity by suppressing thyroid-specific gene expression. Biochimie 1999, 81, 329–340. [Google Scholar] [CrossRef]
- Suzuki, K.; Mori, A.; Lavaroni, S.; Katoh, R.; Kohn, L.D.; Kawaoi, A. Thyroglobulin: A master regulator of follicular function via transcriptional suppression of thyroid specific genes. Acta Histochem. 1999, 32, 111–119. [Google Scholar] [CrossRef]
- Ulianich, L.; Suzuki, K.; Mori, A.; Nakazato, M.; Pietrarelli, M.; Goldsmith, P.; Pacifico, F.; Consiglio, E.; Formisano, S.; Kohn, L.D. Follicular thyroglobulin (TG) suppression of thyroid-restricted genes involves the apical membrane asialoglycoprotein receptor and TG phosphorylation. J. Biol. Chem. 1999, 274, 25099–25107. [Google Scholar] [CrossRef]
- Kohn, L.D.; Suzuki, K.; Nakazato, M.; Royaux, I.; Green, E.D. Effects of thyroglobulin and pendrin on iodide flux through the thyrocyte. Trends Endocrinol. Metab. 2001, 12, 10–16. [Google Scholar] [CrossRef]
- Ohmori, M.; Endo, T.; Harii, N.; Onaya, T. A novel thyroid transcription factor is essential for thyrotropin-induced up-regulation of Na+/I− symporter gene expression. Mol. Endocrinol. 1998, 12, 727–736. [Google Scholar]
- Endo, T.; Kaneshige, M.; Nakazato, M.; Ohmori, M.; Harii, N.; Onaya, T. Thyroid transcription factor-1 activates the promoter activity of rat thytoid Na+/I− symporter gene. Mol. Endocrinol. 1997, 11, 1747–1755. [Google Scholar]
- Joba, W.; Spitzweg, C.; Schriever, K.; Heufelder, A.E. Analysis of human sodium/iodide symporter, thyroid transcription factor-1, and paired-box-protein-8 gene expression in benign thyroid diseases. Thyroid 1999, 9, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.F.; Ando, T.; Lin, R.Y.; Tomer, Y.; Latif, R. Thyrotropin receptor-associated diseases: From adenomata to Graves disease. J. Clin. Investig. 2005, 115, 1972–1983. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, B.; Campbell, V.; Tripp, J.H.; Spyer, G.; Hattersley, A.T.; Ellard, S. Premature birth and low birth weight associated with nonautoimmune hyperthyroidism due to an activating thyrotropin receptor gene mutation. Clin. Endocrinol. 2004, 60, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Rillema, J.A.; Yu, T.X.; Jhiang, S.M. Effect of prolactin on sodium iodide symporter expression in mouse mammary gland explants. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E769–E772. [Google Scholar] [CrossRef] [PubMed]
- Arturi, F.; Ferretti, E.; Presta, I.; Mattei, T.; Scipioni, A.; Scarpelli, D.; Bruno, R.; Lacroix, L.; Tosi, E.; Gulino, A.; et al. Regulation of iodide uptake and sodium/iodide symporter expression in the mcf-7 human breast cancer cell line. J. Clin. Endocrinol. Metab. 2005, 90, 2321–2326. [Google Scholar] [CrossRef] [PubMed]
- Kogai, T.; Taki, K.; Brent, G.A. Enhancement of sodium/iodide symporter expression in thyroid and breast cancer. Endocr. Relat. Cancer 2006, 13, 797–826. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, H.; Yu, X.; Wang, Y.; Yang, C.; Xu, H. Analysis of the Role of Insulin Signaling in Bone Turnover Induced by Fluoride. Biol. Trace Elem. Res. 2016, 171, 380–390. [Google Scholar] [CrossRef]
- Turner, C.H.; Garetto, L.P.; Dunipace, A.J.; Zhang, W.; Wilson, M.E.; Grynpas, M.D.; Chachra, D.; McClintock, R.; Peacock, M.; Stookey, G.K. Fluoride treatment increased serum IGF-1, bone turnover, and bone mass, but not bone strength, in rabbits. Calcif. Tissue Int. 1997, 61, 77–83. [Google Scholar] [CrossRef]
- Lau, W.K.H.; Farley, J.R.; Freeman, T.K.; Baylink, D.J. A proposed mechanism of the mitogenic action of fluoride on bone cells: Inhibition of the activity of osteoblastic acid phosphatase. Metabolism 1989, 38, 858–863. [Google Scholar]
- Ammann, P.; Rizzoli, R.; Caverzasio, J.; Bonjour, J.P. Fluoride potentiates the osteogenic effects of IGF-I in aged ovariectomized rats. Bone 1998, 22, 39–43. [Google Scholar] [CrossRef]
- McCarthy, T.L.; Centrella, M.; Raisz, L.G.; Canalis, E. Prostaglandin E2 stimulates insulin-like growth factor I synthesis in osteoblast-enriched cultures from fetal rat bone. Endocrinology 1991, 128, 2895–2900. [Google Scholar] [CrossRef]
- DiBattista, J.A.; Martel-Pelletier, J.; Wosu, L.O.; Sandor, T.; Antakly, T.; Pelletier, J.P. Glucocorticoid receptor mediated inhibition of interleukin-1 stimulated neutral metalloprotease synthesis in normal human chondrocytes. J. Clin. Endocrinol. Metab. 1991, 72, 316–326. [Google Scholar] [CrossRef]
- Raisz, L.G.; Fall, P.M.; Gabbitas, B.Y.; McCarthy, T.L.; Kream, B.E.; Canalis, E. Effects of prostaglandin Ez on bone formation in cultured fetal rat calvariae: Role of insulin- like growth factor I. Endocrinology 1993, 133, 1504–1510. [Google Scholar] [CrossRef]
- Bichell, D.P.; Rotwein, P.; McCarthy, T.L. Prostaglandin E2 rapidly stimulates insulin-like growth factor I gene expression in primary rat osteoblast cultures: Evidence for transcriptional control. Endocrinology 1993, 133, 1020–1028. [Google Scholar] [CrossRef]
- Fournier, T.; Riches, D.W.; Winston, B.W.; Rose, D.M.; Young, S.K.; Noble, P.W.; Lake, F.R.; Henson, P.M. Divergence in macrophage insulin-like growth factor-I (IGF-I) synthesis induced by TNF-alpha and prostaglandin E2. J. Immunol. 1995, 155, 2123–2133. [Google Scholar]
- Gutowska, I.; Baranowska-Bosiacka, I.; Siennicka, A.; Bakiewicz, M.; Machaliński, B.; Stachowska, E.; Chlube, D. Fluoride and generation of pro-inflammatory factors in human macrophages. Fluoride 2011, 44, 125–134. [Google Scholar]
- Gutowska, I.; Baranowska-Bosiacka, I.; Siennicka, A.; Telesiński, A.; Stańczyk-Dunaj, M.; Wesołowska, T.; Gąssowska, M.; Kłos, P.; Zakrzewska, H.; Machaliński, B.; et al. Activation of phospholipase A2 by low levels of fluoride in THP1 macrophages via altered Ca2+ and cAMP concentration. Prostaglandins Leukot. Essent. Fatty Acids 2012, 86, 99–105. [Google Scholar] [CrossRef]
- Gutowska, I.; Baranowska-Bosiacka, I.; Goschorska, M.; Kolasa, A.; Łukomska, A.; Jakubczyk, K.; Dec, K.; Chlubek, D. Fluoride as a factor initiating and potentiating inflammation in THP1 differentiated monocytes/macrophages. Toxicol. In Vitro 2015, 29, 1661–1668. [Google Scholar] [CrossRef]
- Yan, N.; Liu, Y.; Liu, S. Fluoride-Induced Neuron Apoptosis and Expressions of Inflammatory Factors by Activating Microglia in Rat Brain. Mol. Neurobiol. 2016, 53, 4449–4460. [Google Scholar] [CrossRef]
- Thangapandiyan, S.; Miltonprabu, S. Epigallocatechin gallate supplementation protects against renal injury induced by fluoride intoxication in rats: Role of Nrf2/HO-1 signaling. Toxicol. Rep. 2014, 1, 12–30. [Google Scholar] [CrossRef]
- Thangapandiyan, S.; Miltonprabu, S. Epigallocatechin gallate exacerbates fluoride-induced oxidative stress mediated testicular toxicity in rats through the activation of Nrf2 signaling pathway. Asian Pac. J. Reprod. 2015, 4, 272–287. [Google Scholar] [CrossRef]
- Liu, J.; Xia, T.; Zhang, M.; He, W.; He, P.; Chen, X.; Yang, K.; Wang, A. Screening of Environmental Response Genes Related to Dental Fluorosis. Fluoride 2006, 39, 195–201. [Google Scholar]
- Refsnes, M.; Becher, R.; Lâg, M. Fluoride-induced interleukin-6 and interleukin-8 synthesis in human epithelial lung cells. Hum. Exp. Toxicol. 1999, 18, 645–652. [Google Scholar] [CrossRef]
- Shanmugam, T.; Abdulla, S.; Yakulasamy, V.; Selvaraj, M.; Mathan, R. A mechanism underlying the neurotoxicity induced by sodium fluoride and its reversal by epigallocatechin gallate in the rat hippocampus: Involvement of NrF2/Keap-1 signaling pathway. J. Basic Appl. Zool. 2018, 79, 17. [Google Scholar] [CrossRef]
- Zhou, B.H.; Tan, P.P.; Jia, L.S.; Zhao, W.P.; Wang, J.C.; Wang, H.W. PI3K/AKT signaling pathway involvement in fluoride-induced apoptosis in C2C12 cells. Chemosphere 2018, 199, 297–302. [Google Scholar] [CrossRef]
- Fan, B.; Yu, Y.; Zhang, Y. PI3K-Akt1 expression and its significance in liver tissues with chronic fluorosis. Int. J. Clin. Exp. Pathol. 2015, 8, 1226–1236. [Google Scholar]
- Kanagaraj, V.V.; Panneerselvam, L.; Govindarajan, V.; Ameeramja, J.; Perumal, E. Caffeic acid, a phyto polyphenol mitigates fluoride induced hepatotoxicity in rats: A possible mechanism. Biofactors 2015, 41, 90–100. [Google Scholar] [CrossRef]
- Yu, Y.N.; Yang, D.; Zhu, H.Z.; Deng, C.N.; Guan, Z.Z. Expression of mRNA and protein of p38, Osx, PI3K and Akt1 in rat bone with chronic fluorosis. Chin. J. Pathol. 2012, 41, 622–626. [Google Scholar]
- Zhu, H.Z.; Yu, Y.N.; Deng, C.N.; Yang, D. Effect of fluoride on expression of phosphoinositide 3-kinase, protein kinase B1 mRNA and protein in bone tissue of rats. Chin. J. Endemiol. 2011, 30, 261–265. [Google Scholar]
- Lv, Y.G.; Kang, L.; Wu, G. Fluorosis increases the risk of postmenopausal osteoporosis by stimulating interferon γ. Biochem. Biophys. Res. Commun. 2016, 479, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, Y.; Gao, Y.; Yuan, M.; Manthari, R.K.; Wang, J.; Wang, J. TGF-β1 acts as mediator in fluoride-induced autophagy in the mouse osteoblast cells. Food Chem. Toxicol. 2018, 115, 26–33. [Google Scholar] [CrossRef]
- Sakallioğlu, E.E.; Muğlali, M.; Baş, B.M.; Gulbahar, Y.; Lütfioğlu, M.; Aksoye, A. Effects of Excessive Fluoride intake on Bone Turnover in Mandible: An Immunohistochemical Study in Rabbits. Fluoride 2014, 47, 23–30. [Google Scholar]
- Gao, Y.H.; Fu, S.B.; Sun, H.; Zhou, L.W.; Sun, D.J. Expression of the transforming growth factor-β superfamily in bone turnover of fluorosis. Chin. J. Endemiol. 2006, 25, 374–378. [Google Scholar]
- Yang, C.; Wang, Y.; Xu, H. Fluoride Regulate Osteoblastic Transforming Growth Factor-β1 Signaling by Mediating Recycling of the Type I Receptor ALK5. PLoS ONE 2017, 12, e0170674. [Google Scholar]
- Liu, X.L.; Song, J.; Liu, K.J.; Wang, W.P.; Xu, C.; Zhang, Y.Z.; Liu, Y. Role of inhibition of osteogenesis function by Sema4D/Plexin-B1 signaling pathway in skeletal fluorosis in vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2015, 35, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Yuan, R.; Sun, Y.P.; Lee, T.J.; Shah, G.V. Antiproliferative action of calcitonin on lactotrophs of the rat anterior pituitary gland: Evidence for the involvement of transforming growth factor beta 1 in calcitonin action. Endocrinology 2003, 144, 2164–2171. [Google Scholar] [CrossRef]
- Krishnamachari, K.A.V.R.; Sivakumar, B. Endemic genu valgum. A new dimension to the fluorosis problem in India. Fluoride 1976, 9, 185–200. [Google Scholar]
- Teotia, S.P.S.; Teotia, M.; Singh, R.K.; Teotia, N.P.S.; Taves, D.R.; Heels, S. Plasma fluoride, 25-hydroxycholecalciferol, immunoreactive parathyroid Hormone and calcitonin in patients with endemic skeletal fluorosis. Fluoride 1978, 11, 115–119. [Google Scholar]
- Ma, J.; Li, M.; Song, Y.; Tu, J.; Liu, F.; Liu, K. Serum Osteocalcin and Calcitonin in adult males with different fluoride exposures. Fluoride 2009, 42, 133–136. [Google Scholar]
- Chen, S.; Li, B.; Lin, S.; Huang, Y.; Zhao, X.; Zhang, M.; Xia, Y.; Fang, X.; Wang, J.; Hwang, S.A.; et al. Change of urinary fluoride and bone metabolism indicators in the endemic fluorosis areas of southern china after supplying low fluoride public water. Public Health 2013, 13, 15. [Google Scholar] [CrossRef]
- Shashi, A.; Singla, S. Parathyroid Function in Osteofluorosis. World J. Med. Sci. 2013, 8, 67–73. [Google Scholar]
- Ba, Y.; Zhu, J.Y.; Yang, Y.J.; Yu, B.; Huang, H.; Wang, G.; Ren, L.J.; Cheng, X.M.; Cui, L.X.; Zhang, Y.W. Serum calciotropic hormone levels, and dental fluorisis in children exposed to different concentrations of fluoride and iodine in drinking water. Chin. Med. J. 2010, 123, 675–679. [Google Scholar] [PubMed]
- Xu, Y.; Yuan, S.; Xie, Q. Effects of Fluorosis on induced secretion of rat Prolactin in vivo and in vitro. Fluoride 1997, 30, 173–178. [Google Scholar]
- Ortiz-Pérez, D.; Rodríguez-Martínez, M.; Martínez, F.; Borja-Aburto, V.H.; Castelo, J.; Grimaldo, J.I.; de la Cruz, E.; Carrizales, L.; Díaz-Barriga, F. Fluoride-induced disruption of reproductive hormones in men. Environ. Res. 2003, 93, 20–30. [Google Scholar] [CrossRef]
- Murugan, A.; Subramanian, A. Studies on the Biological Effects of Fluoride Intoxication in Dental Fluorosis Cases. Aust. J. Basic Appl. Sci. 2011, 5, 1362–1367. [Google Scholar]
- Shah, G.V.; Wang, W.; Grosvenor, C.E.; Crowley, W.R. Calcitonin inhibits basal and thyrotropin-releasing hormone-induced release of prolactin from anterior pituitary cells: Evidence for a selective action exerted proximal to secretagogue-induced increases in cytosolic Ca2+. Endocrinology 1990, 127, 621–628. [Google Scholar] [CrossRef]
- Martin, T.F. Calcitonin peptide inhibition of TRH-stimulated prolactin secretion Additional evidence for inhibitory regulation of phospholipase C. Trends Endocrinol. Metab. 1992, 3, 82–85. [Google Scholar] [CrossRef]
- Shah, G.V.; Pedchenko, V.; Stanley, S.; Li, Z.; Samson, W.K. Calcitonin is a physiological inhibitor of prolactin secretion in ovariectomized female rats. Endocrinology 1996, 137, 1814–1822. [Google Scholar] [CrossRef]
- Shah, G.V.; Chien, J.; Sun, Y.P.; Puri, S.; Ravindra, R. Calcitonin inhibits anterior pituitary cell proliferation in the adult female rats. Endocrinology 1999, 140, 4281–4291. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Liu, H.; Qu, W.; Guan, Z.; Zeng, Q.; Jiang, C.; Gao, H.; Zhang, C.; Lei, R.; et al. Modifying Effect of COMT Gene Polymorphism and a Predictive Role for Proteomics Analysis in Children’s Intelligence in Endemic Fluorosis Area in Tianjin, China. Toxicol. Sci. 2015, 144, 238–245. [Google Scholar] [CrossRef]
- Yasmin, S.; Ranjan, S.; D’Souza, D.; D’Souza, H. Effect of excess fluoride ingestion on human thyroid function in Gaya region, Bihar, India. Toxicol. Environ. Chem. 2013, 95, 1235–1243. [Google Scholar] [CrossRef]
- Hosur, M.B.; Puranik, R.S.; Vanaki, S.; Puranik, S. Study of thyroid hormones free triiodothyronine (FT3), free thyroxine (FT4) and thyroid stimulating hormone (TSH) in subjects with dental fluorosis. Eur. J. Dent. 2012, 6, 184–190. [Google Scholar] [PubMed]
- Karademir, S.; Mustafa, A.; Kuybulu, A.E.; Olgar, S.; Öktem, F. Effects of fluorosis on QT dispersion, heart rate variability and echocardiographic parameters in children. Anadolu Kardiyol. Derg. 2011, 1, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Chen, L.; Miang, Y.; Wu, M.; Chen, B. Fluoride and thyroid function in children in two villages in China. J. Toxicol. Environ. Health Sci. 2009, 1, 054–059. [Google Scholar]
- Bahijri, S.M.; Al-Fares, A.; Al-Khateeb, T.; Mufti, A.B. Hyperparathyroidism and Hypothyroidism in Individuals Consuming High Fluoride Intake in Jeddah-Saudi Arabia. Syrian Clin. Lab. Assoc. 2008, 4, 1428–1436. [Google Scholar]
- Jun, D.; Ying, W. Clinical Application of Determination of Serum Thyroid Hormones (TSH, T3, T4, FT3, FT4) Levels in Elderly Patients with Chronic Diseases. J. Radioimmunol. 2006, 03. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-FSMY200603057.htm (accessed on 19 January 2019).
- Ruiz-Payan, A. Chronic Effects of Fluoride on Growth, Blood Chemistry and Thyroid Hormones in Adolescents Residing in Three Communities in Northern Mexico; AAI3214004; ETD Collection for University of Texas: El Paso, TX, USA, 2006. [Google Scholar]
- Wang, X.; Wang, L.; Hu, P.; Guo, X.; Luo, X. Effects of high iodine and high fluorine on children’s intelligence and thyroid function. Chin. J. Endemiol. 2001, 20, 288–290. [Google Scholar]
- Xiaoli, L.; Zhongxue, F.; Jili, H.; Qinlan, W.; Hongyin, W. The Detection of Children’s T3, T4 and TSH Contents in Endemic Fluorosis Area. Endem. Dis. Bull. 1999, 14, 16–17. [Google Scholar]
- Yao, Y.; Zhou, J.; Wang, X.; Cui, Q.; Lin, F. Analysis on the Correlation between TSH and intelligence level of children with Dental Fluorosis from Endemic Fluorosis Regions. Lit. Inf. Prev. Med. 1996, 2, 26–27. [Google Scholar]
- Liu, Z.; Min, H.; Hanwen, W. An Investigation on the Serum Thyroid Hormones and Fluoride Concentrations in Patients with Endemic Fluorosis. Chin. J. Endemiol. 1988, 7, 216–218. [Google Scholar]
- Bachinskii, P.P.; Gutsalenko, O.A.; Naryzhniuk, N.D.; Sidora, V.D.; Shliakhta, A.I. Action of the body fluorine of healthy persons and thyroidopathy patients on the function of hypophyseal-thyroid the system. Probl. Endokrinol. 1985, 31, 25–29. [Google Scholar]
- Yanni, Y. Study on serum T4, T3 and TSH levels in patients with chronic skeletal fluorosis. Chin. J. Endemiol. 1985, 4, 242–243. [Google Scholar]
- Uller, R.P.; Van Herle, A.J.; Chopra, I.J. Comparison of alterations in circulating thyroglobulin, triiodothyronine and thyroxine in response to exogenous (bovine) and endogenous (human) thyrotropin. J. Clin. Endocrinol. Metab. 1973, 37, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Lo Gerfo, P.; Colacchio, T.A.; Colacchio, D.A.; Feind, C.R. Effect of TSH stimulation on serum thyroglobulin in metastatic thyroid cancer. J. Surg. Oncol. 1980, 14, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Okamura, K.; Ikenoue, H.; Shiroozu, A.; Yoshinari, M.; Fujishima, M. TSH dependent elevation of serum thyroglobulin in reversible primary hypothyroidism. Clin. Endocrinol. 1988, 29, 231–237. [Google Scholar] [CrossRef]
- Sanai, T.; Okamura, K.; Rikitake, S.; Fukuda, M.; Onozawa, K.; Sanematsu, M.; Takashima, T.; Miyazono, M.; Ikeda, Y. The high prevalence of reversible subclinical hypothyroidism with elevated serum thyroglobulin levels in chronic kidney disease patients. Clin. Nephrol. 2017, 87, 237–244. [Google Scholar] [CrossRef]
- Giovanella, L.; Imperiali, M.; Ferrari, A.; Palumbo, A.; Furlani, L.; Graziani, M.S.; Castello, R. Serum thyroglobulin reference values according to NACB criteria in healthy subjects with normal thyroid ultrasound. Clin. Chem. Lab. Med. 2012, 50, 891–893. [Google Scholar] [CrossRef] [PubMed]
- Laurberg, P.; Pedersen, K.M. A sensitive radio-immunoassay for serum thyroglobulin--including a correct screening for thyroglobulin autoantibodies. Scand. J. Clin. Lab. Investig. 1987, 47, 685–689. [Google Scholar] [CrossRef]
- Cahoon, E.K.; Rozhko, A.; Hatch, M.; Polyanskaya, O.; Ostroumova, E.; Tang, M.; Nadirov, E.; Yauseyenka, V.; Savasteeva, I.; McConnell, R.J.; et al. Factors associated with serum thyroglobulin levels in a population living in Belarus. Clin. Endocrinol. 2013, 79, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Furlanetto, T.W.; Nguyen, L.Q.; Jameson, J.L. Estradiol increases proliferation and down-regulates the sodium/iodide symporter gene in FRTL-5 cells. Endocrinology 1999, 140, 5705–5711. [Google Scholar] [CrossRef] [PubMed]
- Gietka-Czernel, M. The thyroid gland in postmenopausal women: Physiology and diseases. Prz. Menopauzalny 2017, 16, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Schindler, A.E. Thyroid function and postmenopause. Gynecol. Endocrinol. 2003, 17, 79–85. [Google Scholar] [CrossRef]
- Bordoloi, G.; Jahan, W. A study of thyroid function in premenopausal and postmenopausal women of Dibrugarh town, Assam, India. Int. J. Res. Med. Sci. 2018, 6, 3015–3019. [Google Scholar] [CrossRef]
- Garg, N.; Sodhi, K.; Singh, J.; Badyal, A. Evaluation of subclinical hypothyroidism in women of postmenopausal age group. J. Adv. Res. Biol. Sci. 2014, 4, 20–22. [Google Scholar]
- Kapadia, N.A.; Mehta, N. Comparison of thyroid profile in premenopausal and postmenopausal women. Int. J. Basic Appl. Physiol. 2017, 6, 150–154. [Google Scholar]
- Husdan, H.; Vogi, R.; Oreopoulos, D.; Gryfe, C.; Rapoport, A. Serum Ionic Fluoride: Normal Range and Relationship to Age and Sex. Clin. Chem. 1976, 22, 1884–1888. [Google Scholar]
- Kataraki, P.; Rao, P. Fluoride and oxidative stress in postmenopausal women. IOSR J. Pharm. 2012, 2, 57–59. [Google Scholar] [CrossRef]
- Itai, K.; Onoda, T.; Nohara, M.; Ohsawa, M.; Tanno, K.; Sato, T.; Kuribayashi, T.; Okayama, A. Serum ionic fluoride concentrations are related to renal function and menopause status but not to age in a Japanese general population. Clin. Chim. Acta 2010, 411, 263–266. [Google Scholar] [CrossRef]
- Santisteban, P.; Kohn, L.D.; Di Lauro, R. Thyroglobulin gene expression is regulated by insulin and insulin-like growth factor 1, as well as thyrotropin, in FRTL-5 thyroid cell. J. Biol. Chem. 1987, 262, 4048–4052. [Google Scholar] [PubMed]
- Medina, D.L.; Suzuki, K.; Pietrarelli, M.; Okajima, F.; Kohn, L.D.; Santisteban, P. Role of insulin and serum on thyrotropin regulation of thyroid transcription factor-1 and pax-8 genes expression in FRTL-5 thyroid cells. Thyroid 2000, 10, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, I. Osteocalcin as a hormone regulating glucose metabolism. World J. Diabetes 2015, 6, 1345–1354. [Google Scholar] [CrossRef] [PubMed]
- Rui, X.; Xu, B.; Su, J.; Pan, C.; Zhan, C.; Su, B.; Li, H.; Wang, J.; Sheng, H.; Qu, S. Differential pattern for regulating insulin secretion, insulin resistance, and lipid metabolism by osteocalcin in male and female T2DM patients. Med. Sci. Monit. 2014, 20, 711–719. [Google Scholar]
- Kanazawa, I.; Yamaguchi, T.; Tada, Y.; Yamauchi, M.; Yano, S.; Sugimoto, T. Serum osteocalcin level is positively associated with insulin sensitivity and secretion in patients with type 2 diabetes. Bone 2011, 48, 720–725. [Google Scholar] [CrossRef]
- Aoki, A.; Muneyuki, T.; Yoshida, M.; Munakata, H.; Ishikawa, S.E.; Sugawara, H.; Kawakami, M.; Kakei, M. Circulating osteocalcin is increased in early-stage diabetes. Diabetes Res. Clin. Pract. 2011, 92, 181–186. [Google Scholar] [CrossRef]
- Winhofer, Y.; Handisurya, A.; Tura, A.; Bittighofer, C.; Klein, K.; Schneider, B.; Bieglmayer, C.; Wagner, O.F.; Pacini, G.; Luger, A.; et al. Osteocalcin Is Related to Enhanced Insulin Secretion in Gestational Diabetes Mellitus. Diabetes Care 2010, 33, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Real, J.M.; Izquierdo, M.; Ortega, F.; Gorostiaga, E.; Gómez-Ambrosi, J.; Moreno-Navarrete, J.M.; Frühbeck, G.; Martínez, C.; Idoate, F.; Salvador, J.; et al. The relationship of serum osteocalcin concentration to insulin secretion, sensitivity, and disposal with hypocaloric diet and resistance training. J. Clin. Endocrinol. Metab. 2009, 94, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Ferron, M.; Hinoi, E.; Karsenty, G.; Ducy, P. Osteocalcin differentially regulates β cell and adipocyte gene expression and affects the development of metabolic diseases in wild-type mice. Proc. Natl. Acad. Sci. USA 2008, 105, 5266–5270. [Google Scholar] [CrossRef]
- Mizokami, A.; Yasutake, Y.; Gao, J.; Matsuda, M.; Takahashi, I.; Takeuchi, H.; Hirata, M. Osteocalcin induces release of glucagon-like peptide-1 and thereby stimulates insulin secretion in mice. PLoS ONE 2013, 8, e57375. [Google Scholar] [CrossRef] [PubMed]
- Mizokami, A.; Yasutake, Y.; Higashi, S.; Kawakubo-Yasukochi, T.; Chishaki, S.; Takahashi, I.; Takeuchi, H.; Hirata, M. Oral administration of osteocalcin improves glucose utilization by stimulating glucagon-like peptide-1 secretion. Bone 2014, 69, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.N.; Gill, D.S.; Moudgil, A.; Menon, R.K.; Thomas, M.; Dandona, P. Normal ionized calcium, parathyroid hypersecretion, and elevated osteocalcin in a family with fluorosis. Metabolism 1989, 38, 120–124. [Google Scholar] [CrossRef]
- Bouletreau, P.H.; Bost, M.; Fontanges, E.; Lauverjat, M.; Gutknecht, C.; Ecochard, R.; Delmas, P.D.; Chambrier, C. Fluoride exposure and bone status in patients with chronic intestinal failure who are receiving home parenteral nutrition. Am. J. Clin. Nutr. 2006, 83, 1429–1437. [Google Scholar] [CrossRef]
- Battmann, A.; Resch, H.; Libanati, C.R.; Ludy, D.; Fischer, M.; Farley, S.; Baylink, D.J. Serum Fluoride and Serum Osteocalcin Levels in Response to a Novel Sustained-Release Monofluorophosphate Preparation: Comparison with Plain Monofluorophosphate. Osteoporosis Int. 1997, 7, 48–51. [Google Scholar] [CrossRef]
- Huang, H.; Ba, Y.; Cui, L.; Cheng, X.; Zhu, J.; Zhang, Y.; Yan, P.; Zhu, C.; Kilfoy, B.; Zhang, Y. COLIA2 gene polymorphisms (Pvu II and RSA I), serum calcitropic hormone levels, and dental fluorosis. Community Dent. Oral Epidemiol. 2008, 36, 517–522. [Google Scholar] [CrossRef]
- Dandona, P.; Coumar, A.; Gill, D.S.; Bell, J.; Thomas, M. Sodium Fluoride Stimulates Osteocalcin in Normal Subjects. Clin. Endocrinol. 1988, 29, 437–441. [Google Scholar] [CrossRef]
- Pereira, A.G.; Chiba, F.Y.; de Lima Coutinho Mattera, M.S.; Pereira, R.F.; de Cássia Alves Nunes, R.; Tsosura, T.V.S.; Okamoto, R.; Sumida, D.H. Effects of fluoride on insulin signaling and bone metabolism in ovariectomized rats. J. Trace Elem. Med. Biol. 2017, 39, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Rogalska, A.; Kuter, K.; Żelazko, A.; Glogowska-Gruszka, A.; Świętochowska, E.; Nowak, P. Fluoride Alteration of [3H]Glucose Uptake in Wistar Rat Brain and Peripheral Tissues. Neurotox. Res. 2017, 31, 436–443. [Google Scholar] [CrossRef]
- Validandi, V.; Gourineni, S.R.; Dheeravath, S.; Nagalla, B.; Khandare, A.L. Tamarind Supplementation Ameliorates Fluoride-induced Glucose Intolerance and Insulin Resistance in Rats. Fluoride 2017, 50, 314–323. [Google Scholar]
- Hu, C.Y.; Ren, L.Q.; Li, X.N.; Wu, N.; Li, G.S.; Liu, Q.Y.; Xu, H. Effect of fluoride on insulin level of rats and insulin receptor expression in the MC3T3-E1 cells. Biol. Trace Elem. Res. 2012, 150, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Lupo, M.; Afonso, M.; Buzalaf, R.; Rigalli, A. Effect of Fluoridated Water on Plasma Insulin Levels and Glucose Homeostasis in Rats with Renal Deficiency. Biol. Trace Elem. Res. 2011, 140, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, N.; Mithal, A.; Gupta, S.K.; Godbole, M.M. Reversible impairment of glucose tolerance in patients with endemic fluorosis. Fluoride Collaborative Study Group. Diabetologia 1993, 36, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-González, M.; Jacobo Estrada, T.; Rodríguez-Muñoz, R.; Barrera-Chimal, J.; Bobadilla, N.A.; Barbier, O.C.; Del Razo, L.M. Sub-chronic exposure to fluoride impacts the response to a subsequent nephrotoxic treatment with gentamicin. J. Appl. Toxicol. 2016, 36, 309–319. [Google Scholar] [CrossRef]
- Tojo, A.; Onozato, M.L.; Ha, H.; Kurihara, H.; Sakai, T.; Goto, A.; Fujita, T.; Endou, H. Reduced albumin reabsorption in the proximal tubule of early-stage diabetic rats. Histochem. Cell Biol. 2001, 116, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Peruchetti, D.B.; Silva-Aguiar, R.P.; Siqueira, G.M.; Dias, W.B.; Caruso-Neves, C. High glucose reduces megalin-mediated albumin endocytosis in renal proximal tubule cells through protein kinase B O-GlcNAcylation. J. Biol. Chem. 2018, 293, 11388–11400. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Yang, J.H.; Xu, J.; Jia, J.Y.; Zhang, X.R.; Yue, X.D.; Chen, L.M.; Shan, C.Y.; Zheng, M.Y.; Han, F.; et al. Renal tubular damage may contribute more to acute hyperglycemia induced kidney injury in non-diabetic conscious rats. J. Diabetes Complicat. 2015, 29, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Yang, P.; Yu, N.; Zhuang, Z. An Experimental study of blood biochemical diagnostic indices for chronic fluorosis. Fluoride 1989, 22, 108–111. [Google Scholar]
- Shanthakumari, D.; Subramanian, S. Effect of Fluoride Intoxication on Bone Tissue of Experimental Rats. Res. J. Environ. Sci. 2007, 1, 82–92. [Google Scholar]
- Suketa, Y.; Asao, Y.; Kanamoto, Y.; Shakashita, T.; Okada, S. Changes in adrenal function as a possible mechanism for elevation of serum glucose by single large dose of fluoride. Toxicol. Appl. Pharmacol. 1985, 80, 199–205. [Google Scholar] [CrossRef]
- Rigalli, A.; Ballina, J.C.; Roveri, E.; Puche, R.C. Inhibitory effect of fluoride on the secretion of Insulin. Calcif. Tissue Int. 1990, 46, 333–338. [Google Scholar] [CrossRef]
- Grucka-Mamczar, E.; Birkner, E.; Kasperczyk, S.; Kasperczyk, A.; Chlubek, D.; Samujło, D.; Cegłowska, A. Lipid Balance in rats with fluoride-induced hyperglycemia. Fluoride 2004, 37, 195–200. [Google Scholar]
- McGown, E.L.; Suttie, J.W. Mechanism of fluoride-induced hyperglycemia in the Rat. Toxicol. Appl. Pharmacol. 1977, 40, 83–90. [Google Scholar] [CrossRef]
- Allmann, D.W.; Kleiner, H.S. Effect of NaF on Rat Tissue cAMP levels in vivo. Pharmacol. Ther. Dent. 1980, 5, 73–78. [Google Scholar]
- Grucka-Mamczar, E.; Birkner, E.; Zalejska-Fiolka, J.; Machoy, Z. Disturbances of kidney function in rats with fluoride-induced hyperglycemia after acute poisoning by fluoride. Fluoride 2005, 38, 48–51. [Google Scholar]
- Szymafiska, H.; Mandat, A.; Jaroszewicz-Heigelmann, H.; Szymadski, Z.; Holicki, M.; Neuman, Z.; Ruszkowska, A. The results of assorted investigations carried out in workers exposed to Fluorine compounds. Metabolism of Fluorine. Societas Scientiarum Stetinensis. PWN Warszawa-Poznafi. 1982, 96–102. [Google Scholar]
- Chlubek, D.; Grucka-Mamczar, E.; Birkner, E.; Polaniak, R.; Stawiarska-Pieta, B.; Duliban, H. Activity of pancreatic antioxidative enzymes and malondialdehyde concentrations in rats with hyperglycemia caused by fluoride intoxication. J. Trace Elem. Med. Biol. 2003, 17, 57–60. [Google Scholar] [CrossRef]
- Rupal, A.V.; Dhrutigna, R.K.; Krutika, L.B.; Narasimhacharya, A.V.R.L. Therapeutic benefits of glibenclamide in fluoride intoxicated diabetic rats. Fluoride 2010, 43, 141–149. [Google Scholar]
- García-Montalvo, E.A.; Reyes-Pérez, H.; Del Razo, L.M. Fluoride exposure impairs glucose tolerance via decreased insulin expression and oxidative stress. Toxicology 2009, 263, 75–83. [Google Scholar] [CrossRef]
- Grucka-Mamczar, E.; Birkner, E.; Zalejska-Fiolka, J.; Machoy, Z.; Kasperczyk, S.; Blaszczyk, I. Influence of extended exposure to sodium fluoride and caffeine on the activity of carbohydrate metabolism enzymes in rat blood serum and liver. Fluoride 2007, 40, 62–66. [Google Scholar]
- Sakurai, T.; Suzuki, K.; Taki, T.; Suketa, V.T. The mechanism of changes in metabolism and transport of glucose caused by fluoride administration to rats. Fluoride 1993, 26, 210. [Google Scholar]
- Peters, J.P.; Man, E.B. The relation of albumin to precipitable iodine of serum. J. Clin. Investig. 1948, 27, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Riggs, D.S.; Lavietes, P.H.; Man, E.B. Investigations on the nature of blood iodine. J. Biol. Chem. 1942, 143, 363–372. [Google Scholar]
- Roholm, K. Fluorine Intoxication, a Clinical-Hygienic Study with a Review of the Literature and Some Experimental Investigations; H.K. Lewis & Co.: London, UK, 1937. [Google Scholar]
- Jiménez-Córdova, M.I.; Cárdenas-González, M.; Aguilar-Madrid, G.; Sanchez-Peña, L.C.; Barrera-Hernández, Á.; Domínguez-Guerrero, I.A.; González-Horta, C.; Barbier, O.C.; Del Razo, L.M. Evaluation of kidney injury biomarkers in an adult Mexican population environmentally exposed to fluoride and low arsenic levels. Toxicol. Appl. Pharmacol. 2018, 352, 97–106. [Google Scholar] [CrossRef]
- Kumar, S.P.; Harper, R.A. Fluorosis in Aden. Br. J. Radiol. 1963, 36, 497–502. [Google Scholar] [CrossRef]
- Susheela, A.K.; Jethanandani, P. Serum haptoglobin and C-reactive protein in human skeletal fluorosis. Clin. Biochem. 1994, 27, 463–468. [Google Scholar] [CrossRef]
- Shivashankara, A.R.; Shivarajashankara, Y.M.; Rao, S.H.; Bhat, G.P. A clinical and biochemical study of chronic fluoride toxicity in children of Kheru Thanda of Gulbarga District, Karnataka, India. Fluoride 2000, 33, 66–73. [Google Scholar]
- Subramanian, V.S.; Priya, V.V.; Gayathri, R. Estimation of salivary total protein and albumin in dental fluorosis patient. Drug Invent. Today 2018, 10, 1734–1736. [Google Scholar]
- Awdeh, Z.L.; Islam, M.R.; Samra, S.A. Variations in the level of human serum albumin during glucose tolerance test. Biochem. Biophys. Res. Commun. 1974, 56, 358–362. [Google Scholar] [CrossRef]
- Thrailkill, K.M.; Nimmo, T.; Bunn, R.C.; Cockrell, G.E.; Moreau, C.S.; Mackintosh, S.; Edmondson, R.D.; Fowlkes, J.L. Microalbuminuria in type 1 diabetes mellitus is associated with enhanced excretion of the endocytic, multiligand receptors megalin and cubilin. Diabetes Care 2009, 32, 1266–1268. [Google Scholar] [CrossRef] [PubMed]
- De la Vieja, A.; Santisteban, P. Role of iodide metabolism in physiology and cancer. Endocr. Relat. Cancer 2018, 25, R225–R245. [Google Scholar] [CrossRef]
- Garg, L.C.; Tisher, C.C. Effects of thyroid hormone on Na-K-adenosine triphosphatase activity along the rat nephron. J. Lab. Clin. Med. 1985, 106, 568–572. [Google Scholar]
- Levy, O.; Dai, G.; Riedel, C.; Ginter, C.S.; Paul, E.M.; Lebowitz, A.N.; Carrasco, N. Characterization of the thyroid Na+/I− symporter with an anti-COOH terminus antibody. Proc. Natl. Acad. Sci. USA 1997, 94, 5568–5573. [Google Scholar] [CrossRef]
- Paire, A.; Bernier-Valentin, F.; Selmi-Ruby, S.; Rousset, B. Characterization of the rat thyroid iodide transporter using anti-peptide antibodies. Relationship between its expression and activity. J. Biol. Chem. 1997, 272, 18245–18249. [Google Scholar] [CrossRef]
- Kogai, T.; Endo, T.; Saito, T.; Miyazaki, A.; Kawaguchi, A.; Onaya, T. Regulation by thyroid-stimulating hormone of sodium/iodide symporter gene expression and protein levels in FRTL-5 cells. Endocrinology 1997, 138, 2227–2232. [Google Scholar] [CrossRef]
- Saito, T.; Endo, T.; Kawaguchi, A.; Ikeda, M.; Nakazato, M.; Kogai, T.; Onaya, T. Increased expression of the Na+/I− symporter in cultured human thyroid cells exposed to thyrotropin and in Graves’ thyroid tissue. J. Clin. Endocrinol. Metab. 1997, 82, 3331–3336. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.S.; Santarosa, P.L.; Granja, F.; da Assumpção, L.V.M.; Savoldi, M.; Goldman, G.H. Low expression of sodium iodide symporter identifies aggressive thyroid tumors. Cancer Lett. 2003, 200, 85–91. [Google Scholar] [CrossRef]
- Scipioni, A.; Ferretti, E.; Soda, G.; Tosi, E.; Bruno, R.; Costante, G.; Meringolo, D.; Arturi, F.; Durante, C.; Amorosi, A.; et al. hNIS protein in thyroid: The iodine supply influences its expression and localization. Thyroid 2007, 17, 613–618. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Fluorides and Oral Health; Technical Report Series-846; WHO: Geneva, Switzerland, 1984. [Google Scholar]
- Beltrán-Aguilar, E.D.; Barker, L.; Dye, B.A. Prevalence and severity of dental fluorosis in the United States, 1999–2004. NCHS Data Brief 2010, 1–8. [Google Scholar]
- Marshall, T.A.; Levy, S.M.; Warren, J.J.; Broffitt, B.; Eichenberger-Gilmore, J.M.; Stumbo, P.J. Associations between Intakes of Fluoride from Beverages during Infancy and Dental Fluorosis of Primary Teeth. J. Am. Coll. Nutr. 2004, 23, 108–116. [Google Scholar] [CrossRef]
- Whelton, H.; Crowley, E.; O’Mullane, D.; Donaldson, M.; Kelleher, V.; Cronin, M. Dental caries and enamel fluorosis among the fluoridated and non-fluoridated populations in the Republic of Ireland in 2002. Community Dent. Health 2004, 21, 37–44. [Google Scholar] [PubMed]
- Delange, F. The disorders induced by iodine deficiency. Thyroid 1994, 4, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Instanes, J.T.; Halmøy, A.; Engeland, A.; Haavik, J.; Furu, K.; Klungsøyr, K. Attention-Deficit/Hyperactivity Disorder in Offspring of Mothers with Inflammatory and Immune System Diseases. Biol. Psychiatry 2017, 81, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Abel, M.H.; Ystrom, E.; Caspersen, I.H.; Meltzer, H.M.; Aase, H.; Torheim, L.E.; Askeland, R.B.; Reichborn-Kjennerud, T.; Brantsæter, A.L. Maternal Iodine Intake and Offspring Attention-Deficit/Hyperactivity Disorder: Results from a Large Prospective Cohort Study. Nutrients 2017, 9, 1239. [Google Scholar] [CrossRef]
- Kanık Yüksek, S.; Aycan, Z.; Öner, Ö. Evaluation of Iodine Deficiency in Children with Attention Deficit/Hyperactivity Disorder. J. Clin. Res. Pediatr. Endocrinol. 2016, 8, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Konikowska, K.; Regulska-Ilow, B.; Rózańska, D. The influence of components of diet on the symptoms of ADHD in children. Rocz. Panstw. Zakl. Hig. 2012, 63, 127–134. [Google Scholar] [PubMed]
- Vermiglio, F.; Lo Presti, V.P.; Moleti, M.; Sidoti, M.; Tortorella, G.; Scaffidi, G.; Castagna, M.G.; Mattina, F.; Violi, M.A.; Crisà, A.; et al. Attention deficit and hyperactivity disorders in the offspring of mothers exposed to mild-moderate iodine deficiency: A possible novel iodine deficiency disorder in developed countries. J. Clin. Endocrinol. Metab. 2004, 89, 6054–6060. [Google Scholar] [CrossRef]
- Getahun, D.; Jacobsen, S.J.; Fassett, M.J.; Wing, D.A.; Xiang, A.H.; Chiu, V.Y.; Peltier, M.R. Association between maternal hypothyroidism and autism spectrum disorders in children. Pediatr. Res. 2018, 83, 580–588. [Google Scholar] [CrossRef]
- Andersen, S.L.; Laurberg, P.; Wu, C.S.; Olsen, J. Attention deficit hyperactivity disorder and autism spectrum disorder in children born to mothers with thyroid dysfunction: A Danish nationwide cohort study. BJOG 2014, 121, 1365–1374. [Google Scholar] [CrossRef]
- Błażewicz, A.; Makarewicz, A.; Korona-Glowniak, I.; Dolliver, W.; Kocjan, R. Iodine in autism spectrum disorders. J. Trace Elem. Med. Biol. 2016, 34, 32–37. [Google Scholar] [CrossRef]
- Hamza, R.T.; Hewedi, D.H.; Sallam, M.T. Iodine deficiency in Egyptian autistic children and their mothers: Relation to disease severity. Arch. Med. Res. 2013, 44, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, K.M. Iodine deficiency as a cause of autism. J. Neurol. Sci. 2009, 276, 202. [Google Scholar] [CrossRef] [PubMed]
- Gillberg, I.C.; Gillberg, C.; Kopp, S. Hypothyroidism and autism spectrum disorders. J. Child Psychol. Psychiatry 1992, 33, 531–542. [Google Scholar] [CrossRef]
- Andersson, M.; de Benoist, B.; Darnton-Hill, I. (Eds.) WHO, Unicef. In Iodine Deficiency in Europe: A Continuing Public Health Problem; World Health Organization: Geneva, Switzerland, 2007; Available online: https://www.who.int/nutrition/publications/VMNIS_Iodine_deficiency_in_Europe.pdf (accessed on 19 January 2019).
- Lynn, R.; Vanhanen, T. IQ and the Wealth of Nations; Praeger: Westport, CT, USA, 2012. [Google Scholar]
- Yu, X.; Chen, J.; Li, Y.; Liu, H.; Hou, C.; Zeng, Q.; Cui, Y.; Zhao, L.; Li, P.; Zhou, Z.; et al. Threshold effects of moderately excessive fluoride exposure on children’s health: A potential association between dental fluorosis and loss of excellent intelligence. Environ. Int. 2018, 118, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Razdan, P.; Patthi, B.; Kumar, J.K.; Agnihotri, N.; Chaudhari, P.; Prasad, M. Effect of Fluoride Concentration in Drinking Water on Intelligence Quotient of 12–14-Year-Old Children in Mathura District: A Cross-Sectional Study. J. Int. Soc. Prev. Community Dent. 2017, 7, 252–258. [Google Scholar] [PubMed]
- Khan, S.A.; Singh, R.K.; Navit, S.; Chadha, D.; Johri, N.; Navit, P.; Sharma, A.; Bahuguna, R. Relationship Between Dental Fluorosis and Intelligence Quotient of School Going Children in and Around Lucknow District: A Cross-Sectional Study. J. Clin. Diagn. Res. 2015, 9, ZC10–ZC15. [Google Scholar] [CrossRef] [PubMed]
- Shivaprakash, P.K.; Ohri, K.; Noorani, H. Relation between dental fluorosis and Intelligence quotient in school children of Bagalkot district. J. Indian Soc. Pedod. Prev. Dent. 2011, 29, 117–120. [Google Scholar] [CrossRef]
- Sudhir, K.M.; Chandu, G.N.; Prashant, G.M.; Reddy, S.V.V. Effect of fluoride exposure on Intelligence Quotient (IQ) among 13-15year old school children of known endemic area of fluorosis, Nalgonda District, Andhra Pradesh. J. Indian Assoc. Public Health Dent. 2009, 7, 88–94. [Google Scholar]
- Ding, Y.; Gao, Y.; Sun, H.; Han, H.; Wang, W.; Ji, X.; Liu, X.; Sun, D. The relationships between low levels of urine fluoride on children’s intelligence, dental fluorosis in endemic fluorosis areas in Hulunbuir, Inner Mongolia, China. J. Hazard. Mater. 2011, 186, 1942–1946. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Mondal, N.K. Dental fluorosis and urinary fluoride concentration as a reflection of fluoride exposure and its impact on IQ level and BMI of children of Laxmisagar, Simlapal Block of Bankura District, W.B., India. Environ. Monit. Assess. 2016, 188, 218. [Google Scholar] [CrossRef] [PubMed]
- Bashash, M.; Thomas, D.; Hu, H.; Martinez-Mier, E.A.; Sanchez, B.N.; Basu, N.; Peterson, K.E.; Ettinger, A.S.; Wright, R.; Zhang, Z.; et al. Prenatal Fluoride Exposure and Cognitive Outcomes in Children at 4 and 6–12 Years of Age in Mexico. Environ. Health Perspect. 2017, 125, 097017. [Google Scholar] [CrossRef]
- Bashash, M.; Marchand, M.; Hu, H.; Till, C.; Martinez-Mier, E.A.; Sanchez, B.N.; Basu, N.; Peterson, K.E.; Green, R.; Schnaas, L.; et al. Prenatal fluoride exposure and attention deficit hyperactivity disorder (ADHD) symptoms in children at 6–12 years of age in Mexico City. Environ. Int. 2018, 121, 658–666. [Google Scholar] [CrossRef]
- Dugrillon, A. Iodolactones and iodoaldehydes-mediators of iodine in thyroid autoregulation. Exp Clin. Endocrinol. Diabetes 1996, 104, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Beukelman, C.; van den Berg, A.J.; Hoekstra, M.J.; Uhl, R.; Reimer, K.; Mueller, S. Anti-inflammatory properties of a liposomal hydrogel with povidone-iodine (Repithel) for wound healing in vitro. Burns 2008, 34, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.; Thomas, A.; Harding, K. Iodine released from the wound dressing Iodosorb modulates the secretion of cytokines by human macrophages responding to bacterial lipopolysaccharide. Int. J. Biochem. Cell Biol. 1997, 29, 163–171. [Google Scholar] [CrossRef]
- Nyska, A.; Lomnitski, L.; Maronpot, R.; Moomaw, C.; Brodsky, B.; Sintov, A.; Wormser, U. Effects of iodine on inducible nitric oxide synthase and cyclooxygenase-2 expression in sulfur mustard-induced skin injury in guinea pigs. Arch. Toxicol. 2001, 74, 768–774. [Google Scholar] [CrossRef]
- Venturi, S.; Venturi, M. Iodide, thyroid and stomach carcinogenesis: Evolutionary story of a primitive antioxidant? Eur. J. Endocrinol. 1999, 140, 371–372. [Google Scholar] [CrossRef]
- Winkler, R.; Griebenow, S.; Wonisch, W. Effect of iodide on total antioxidant status of human serum. Cell Biochem. Funct. 2000, 18, 143–146. [Google Scholar] [CrossRef]
- Smyth, P.P. Role of iodine in antioxidant defence in thyroid and breast disease. Biofactors 2003, 19, 121–130. [Google Scholar] [CrossRef]
- Garcia-Solis, P.; Alfaro, Y.; Anguiano, B.; Delgado, G.; Guzman, R.C.; Nandi, S.; Díaz-Muñoz, M.; Vázquez-Martínez, O.; Aceves, C. Inhibition of N-methyl-N-nitrosourea-induced mammary carcinogenesis by molecular iodine (I2) but not by iodide (I−) treatment Evidencethat (I2) prevents cancer promotion. Mol. Cell. Endocrinol. 2005, 236, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, S.J. The iron-H2O2-iodide cytotoxic system. J. Exp. Med. 1982, 156, 1262–1267. [Google Scholar] [CrossRef] [PubMed]
- Majerus, P.M.; Courtois, P.A. Susceptibility of Candida albicans toperoxidase-catalyzed oxidation products of thiocyanate, iodide and bromide. J. Biol. Buccale 1992, 20, 241–245. [Google Scholar] [PubMed]
- Cann, S.A.; van Netten, J.P.; Glover, D.W.; van Netten, C. Iodide accumulation in extrathyroidal tissues. J. Clin. Endocrinol. Metab. 1999, 84, 821–822. [Google Scholar] [CrossRef]
- Bosch, E.H.; van Doorne, H.; de Vries, S. The lactoperoxidase system: The influence of iodide and the chemical and antimicrobial stability over the period of about 18 months. J. Appl. Microb. 2000, 89, 215–224. [Google Scholar] [CrossRef]
- Ihalin, R.; Loimaranta, V.; Tenovuo, J. Origin, structure, and biological activities of peroxidases in human saliva. Arch. Biochem. Biophys. 2006, 445, 261–268. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Choi, H.; Kushida, Y.; Bhayana, B.; Wang, Y.; Hamblin, M.R. Broad-spectrum antimicrobial effects of photocatalysis using titanium dioxide nanoparticles are strongly potentiated by addition of potassium iodide. Antimicrob. Agents Chemother. 2016, 60, 5445–5453. [Google Scholar] [CrossRef]
- Fischer, A.J.; Lennemann, N.J.; Krishnamurthy, S.; Pócza, P.; Durairaj, L.; Launspach, J.L.; Rhein, B.A.; Wohlford-Lenane, C.; Lorentzen, D.; Bánfi, B.; et al. Enhancement of respiratory mucosal anti-viral defenses by iodide oxidation. Am. J. Respir. Cell Mol. Biol. 2011, 45, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Kargar, S.; Shiryazdi, S.M.; Atashi, S.R.; Neamatzadeh, H.; Kamali, M. Urinary Iodine Concentrations in Cancer Patients. Asian Pac. J. Cancer Prev. 2017, 18, 819–821. [Google Scholar]
- Cann, S.A.; van Netten, J.P.; van Netten, C. Iodine, selenium and the development of breast cancer. Cancer Causes Control 2000, 11, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, J. Changes in Dietary Iodine Explains Increasing Incidence of Breast Cancer with Distant Involvement in Young Women. J. Cancer 2017, 8, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Eskin, B.A. Iodine metabolism and breast cancer. Trans. N. Y. Acad. Sci. 1970, 32, 911–947. [Google Scholar] [CrossRef]
- Stadel, B.V. Dietary iodine and risk of breast, endometrial, and ovarian cancer. Lancet 1976, 307, 890–891. [Google Scholar] [CrossRef]
- Franceschi, S. Iodine intake and thyroid carcinoma—A potential risk factor. Exp. Clin. Endocrinol. Diabetes 1998, 106, 38–44. [Google Scholar] [CrossRef]
- Feldt-Rasmussen, U. Iodine and cancer. Thyroid 2001, 11, 483–486. [Google Scholar] [CrossRef]
- Ohshima, M.; Ward, J.M. Dietary iodine deficiency as a tumor promoter and carcinogen in male F344/NCr rats. Cancer Res. 1986, 46, 877–883. [Google Scholar] [PubMed]
- Fang, W.T.; Qao, B.S.; Wang, J.B. Iodine deficiency induces thyroid cancer in rats and mice. Zhonghua Zhong Liu Za Zhi 1994, 16, 341–344. [Google Scholar]
- Zimmermann, M.; Galetti, V. Iodine intake as a risk factor for thyroid cancer: A comprehensive review of animal and human studies. Thyroid Res. 2015, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.M.; Ohshima, M. The role of iodine in carcinogenesis. Adv. Exp. Med. Biol. 1986, 206, 529–542. [Google Scholar]
- Mitro, S.D.; Rozek, L.S.; Vatanasapt, P.; Suwanrungruang, K.; Chitapanarux, I.; Srisukho, S.; Sriplung, H.; Meza, R. Iodine deficiency and thyroid cancer trends in three regions of Thailand, 1990–2009. Cancer Epidemiol. 2016, 43, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Hoption Cann, S.A.; Qiu, Z.; van Netten, C. A prospective study of iodine status, thyroid function, and prostate cancer risk: Follow-up of the First National Health and Nutrition Examination Survey. Nutr. Cancer 2007, 58, 28–34. [Google Scholar] [CrossRef]
- Allen, N.E.; Sauvaget, C.; Roddam, A.W.; Appleby, P.; Nagano, J.; Suzuki, G.; Key, T.J.; Koyama, K. A prospective study of diet and prostate cancer in Japanese men. Cancer Causes Control 2004, 15, 911–920. [Google Scholar] [CrossRef]
- Aranda, N.; Sosa, S.; Delgado, G.; Aceves, C.; Anguiano, B. Uptake and antitumoral effects of iodine and 6-iodolactone in differentiated and undifferentiated human prostate cancer cell lines. Prostate 2013, 73, 31–41. [Google Scholar] [CrossRef]
- Aceves, C.; Anguiano, B. Is Iodine an Anti-oxidant and Antiproliferative Agent for the Mammary and Prostate Glands? In Comprehensive Handbook of Iodine: Nutritional, Endocrine and Pathological Aspects; Preedy, V.R., Burrow, G.N., Watson, R.R., Eds.; Academic Press (Elsevier): London, UK, 2009; pp. 249–257. [Google Scholar]
- Aceves, C.; García-Solís, P.; Arroyo-Helguera, O.; Vega-Riveroll, L.; Delgado, G.; Anguiano, B. Antineoplastic effect of iodine in mammary cancer: Participation of 6-iodolactone (6-IL) and peroxisome proliferator-activated receptors (PPAR). Mol. Cancer 2009, 8, 33. [Google Scholar] [CrossRef]
- Mendieta, I.; Nunez-Anita, E.; Delgado, G.; Aceves, C. Differential effect of iodine on the implantation and metastatic potential of xenografts from two different human breast cancer cell lines. Cancer Res. 2011, 71 (Suppl. 8), 4224. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- World Cancer Research Fund; American Institute for Cancer Research. Global Cancer Data by Country. Available online: https://www.wcrf.org/dietandcancer/cancer-trends/data-cancer-frequency-country (accessed on 22 January 2019).
- Carretero, J.; Sánchez-Robledo, V.; Carretero-Hernández, M.; Catalano-Iniesta, L.; García-Barrado, M.J.; Iglesias-Osma, M.C.; Blanco, E.J. Prolactin system in the hippocampus. Cell Tissue Res. 2019, 375, 193–199. [Google Scholar] [CrossRef]
- Benker, G.; Jaspers, C.; Hausler, G.; Reinwein, D. Control of prolactin secretion. Klinische Wochenschrift. 1990, 68, 1157–1167. [Google Scholar] [CrossRef]
- Larsen, C.M.; Grattan, D.R. Prolactin, neurogenesis, and maternal behaviors. Brain Behav. Immun. 2012, 26, 201–209. [Google Scholar] [CrossRef]
- Ferraris, J.; Bernichtein, S.; Pisera, D.; Goffin, V. Use of prolactin receptor antagonist to better understand prolactin regulation of pituitary homeostasis. Neuroendocrinology 2013, 98, 171–179. [Google Scholar] [CrossRef]
- Patil, M.J.; Henry, M.A.; Akopian, A.N. Prolactin receptor in regulation of neuronal excitability and channels. Channels 2014, 8, 193–202. [Google Scholar] [CrossRef]
- Reyes-Mendoza, J.; Morales, T. Post-treatment with prolactin protects hippocampal CA1 neurons of the ovariectomized female rat against kainic acid-induced neurodegeneration. Neuroscience 2016, 328, 58–68. [Google Scholar] [CrossRef]
- Morales, T. Recent findings on neuroprotection against excitotoxicity in the hippocampus of female rats. J. Neuroendocrinol. 2011, 23, 994–1001. [Google Scholar] [CrossRef]
- Tejadilla, D.; Cerbón, M.; Morales, T. Prolactin reduces the damaging effects of excitotoxicity in the dorsal hippocampus of the female rat independently of ovarian hormones. Neuroscience 2010, 169, 1178–1185. [Google Scholar] [CrossRef]
- Morales, T.; Lorenson, M.; Walker, A.M. Both prolactin (PRL) and a molecular mimic of phosphorylated PRL, S179D-PRL, protect the hippocampus of female rats against excitotoxicity. Neuroscience 2014, 258, 211–217. [Google Scholar] [CrossRef]
- Torner, L.; Karg, S.; Blume, A.; Kandasamy, M.; Kuhn, H.G.; Winkler, J.; Aigner, L.; Neumann, I.D. Prolactin prevents chronic stress-induced decrease of adult hippocampal neurogenesis and promotes neuronal fate. J. Neurosci. 2009, 29, 1826–1833. [Google Scholar] [CrossRef]
- Cabrera, V.; Cantú, D.; Ramos, E.; Vanoye-Carlo, A.; Cerbón, M.; Morales, T. Lactation is a natural model of hippocampus neuroprotection against excitotoxicity. Neurosci. Lett. 2009, 461, 136–139. [Google Scholar] [CrossRef]
- Vergara-Castañeda, E.; Grattan, D.R.; Pasantes-Morales, H.; Pérez-Domínguez, M.; Cabrera-Reyes, E.A.; Morales, T.; Cerbón, M. Prolactin mediates neuroprotection against excitotoxicity in primarycell cultures of hippocampal neurons via its receptor. Brain Res. 2016, 1636, 193–199. [Google Scholar] [CrossRef]
- Walker, T.L.; Vukovic, J.; Koudijs, M.M.; Blackmore, D.G.; Mackay, E.W.; Sykes, A.M.; Overall, R.W.; Hamlin, A.S.; Bartlett, P.F. Prolactin stimulates precursor cells in the adult mouse hippocampus. PLoS ONE 2012, 7, e44371. [Google Scholar] [CrossRef]
- Rivero-Segura, N.A.; Flores-Soto, E.; García de la Cadena, S.; Coronado-Mares, I.; Gómez-Verjan, J.C.; Ferreira, D.G.; Cabrera-Reyes, E.A.; Lopes, L.V.; Massieu, L.; Cerbón, M. Prolactin-induced neuro-protection against glutamate excitotoxicity is mediated by the reduction of [Ca2+]i overload and NF-κB activation. PLoS ONE 2017, 12, e0176910. [Google Scholar] [CrossRef]
- Kinsley, C.H.; Trainer, R.; Stafisso-Sandoz, G.; Quadros, P.; Marcus, L.K.; Hearon, C.; Meyer, E.A.; Hester, N.; Morgan, M.; Kozub, F.J.; et al. Motherhood and the hormones of pregnancy modify concentrations of hippocampal neuronal dendritic spines. Horm. Behav. 2006, 49, 131–142. [Google Scholar] [CrossRef]
- Brusco, J.; Wittmann, R.; de Azevedo, M.; Lcion, A.; Franci, C.; Giovenardi, M.; Rasia-Filho, A.A. Plasma hormonal profiles and dendritic spines density and morphology in the hippocampal CA1 stratum radiatum, evidenced by light microscopy, of virgin and postpartum female rats. Neurosci. Lett. 2008, 438, 346–350. [Google Scholar] [CrossRef]
- Infant and Young Child Feeding: Model Chapter for Textbooks for Medical Students and Allied Health Professionals; SESSION 2, The Physiological Basis of Breastfeeding; World Health Organization: Geneva, Switzerland, 2009. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK148970/ (accessed on 19 January 2019).
- Balbach, L.; Wallaschofski, H.; Volzke, H.; Nauck, M.; Dorr, M.; Haring, R. Serum prolactin concentrations as risk factor of metabolic syndrome or type 2 diabetes? BMC Endocr. Disord. 2013, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, Y.; Xu, M.; Ning, G.; Lu, J.; Dai, M.; Xu, B.; Sun, J.; Sun, W.; Lai, S.; et al. Circulating Prolactin and Risk of Type 2 Diabetes: A Prospective Study. Am. J. Epidemiol. 2015, 184, 295–301. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Corona, G.; Mannucci, E.; Jannini, E.A.; Lotti, F.; Ricca, V.; Monami, M.; Boddi, V.; Bandini, E.; Balercia, G.; Forti, G.; et al. Hypoprolactinemia: A new clinical syndrome in patients with sexual dysfunction. J. Sex. Med. 2009, 6, 1457–1466. [Google Scholar] [CrossRef]
- Rutter, M.K.; Maggi, M.; EMAS Study Group. Low prolactin is associated with sexual dysfunction and psychological or metabolic disturbances in middle aged and elderly men: The European Male Aging Study (EMAS). J. Sex. Med. 2014, 11, 240–253. [Google Scholar]
- Corona, G.; Rastrelli, G.; Boddi, V.; Monami, M.; Melani, C.; Balzi, D.; Sforza, A.; Forti, G.; Mannucci, E.; Maggi, M. Prolactin levels independently predict major cardiovascular events in patients with erectile dysfunction. Int. J. Androl. 2011, 34, 217–224. [Google Scholar] [CrossRef]

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waugh, D.T. Fluoride Exposure Induces Inhibition of Sodium/Iodide Symporter (NIS) Contributing to Impaired Iodine Absorption and Iodine Deficiency: Molecular Mechanisms of Inhibition and Implications for Public Health. Int. J. Environ. Res. Public Health 2019, 16, 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16061086
Waugh DT. Fluoride Exposure Induces Inhibition of Sodium/Iodide Symporter (NIS) Contributing to Impaired Iodine Absorption and Iodine Deficiency: Molecular Mechanisms of Inhibition and Implications for Public Health. International Journal of Environmental Research and Public Health. 2019; 16(6):1086. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16061086
Chicago/Turabian StyleWaugh, Declan Timothy. 2019. "Fluoride Exposure Induces Inhibition of Sodium/Iodide Symporter (NIS) Contributing to Impaired Iodine Absorption and Iodine Deficiency: Molecular Mechanisms of Inhibition and Implications for Public Health" International Journal of Environmental Research and Public Health 16, no. 6: 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16061086