Effects of Lanthanum on the Photosystem II Energy Fluxes and Antioxidant System of Chlorella Vulgaris and Phaeodactylum Tricornutum


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Algae Culture
2.2. Experimental Design
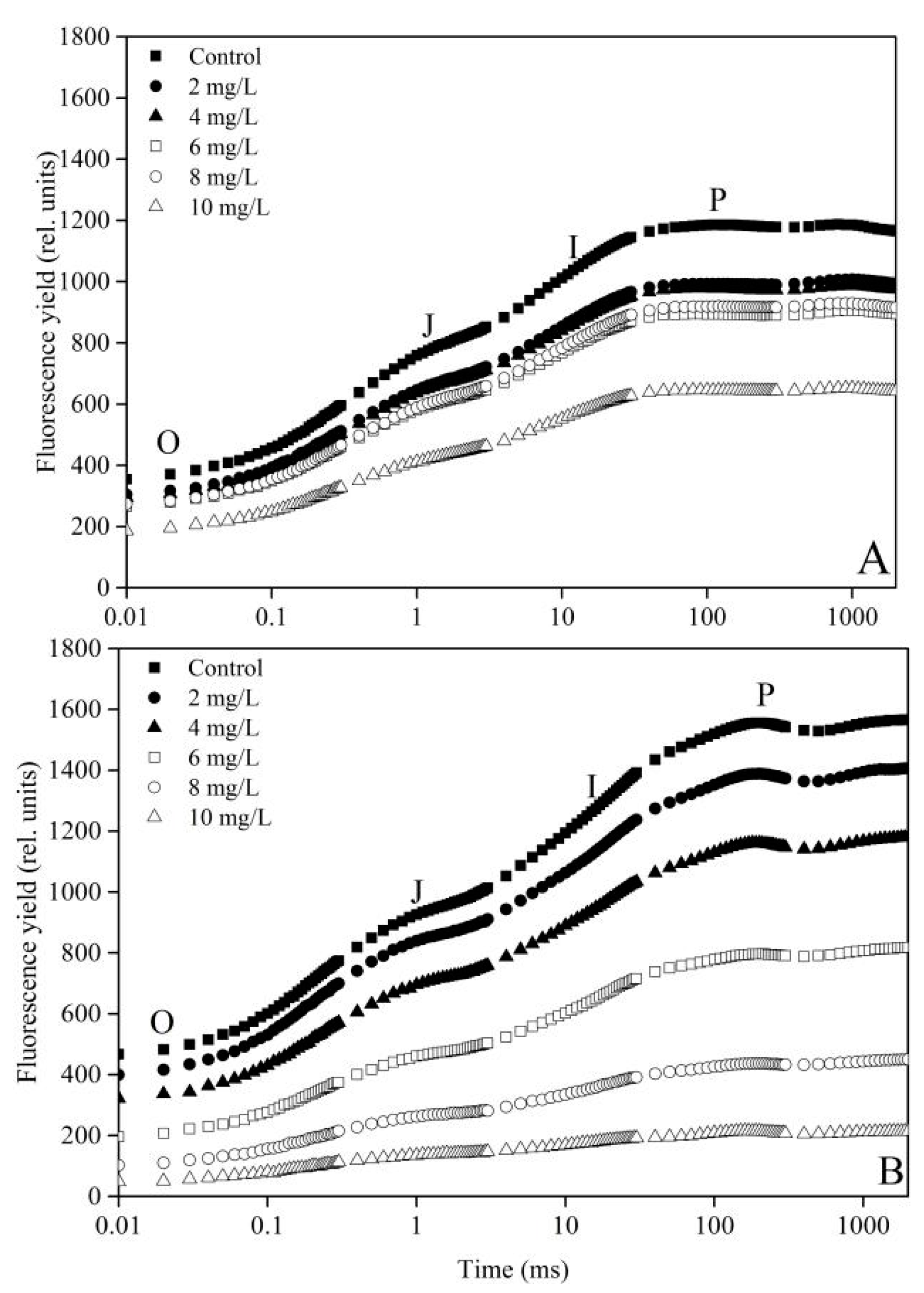
2.3. Measurements of Fluorescence Transient
2.4. Extraction and Determination of Protein and Antioxidase Levels
2.5. Data Analysis and Statistics
3. Results
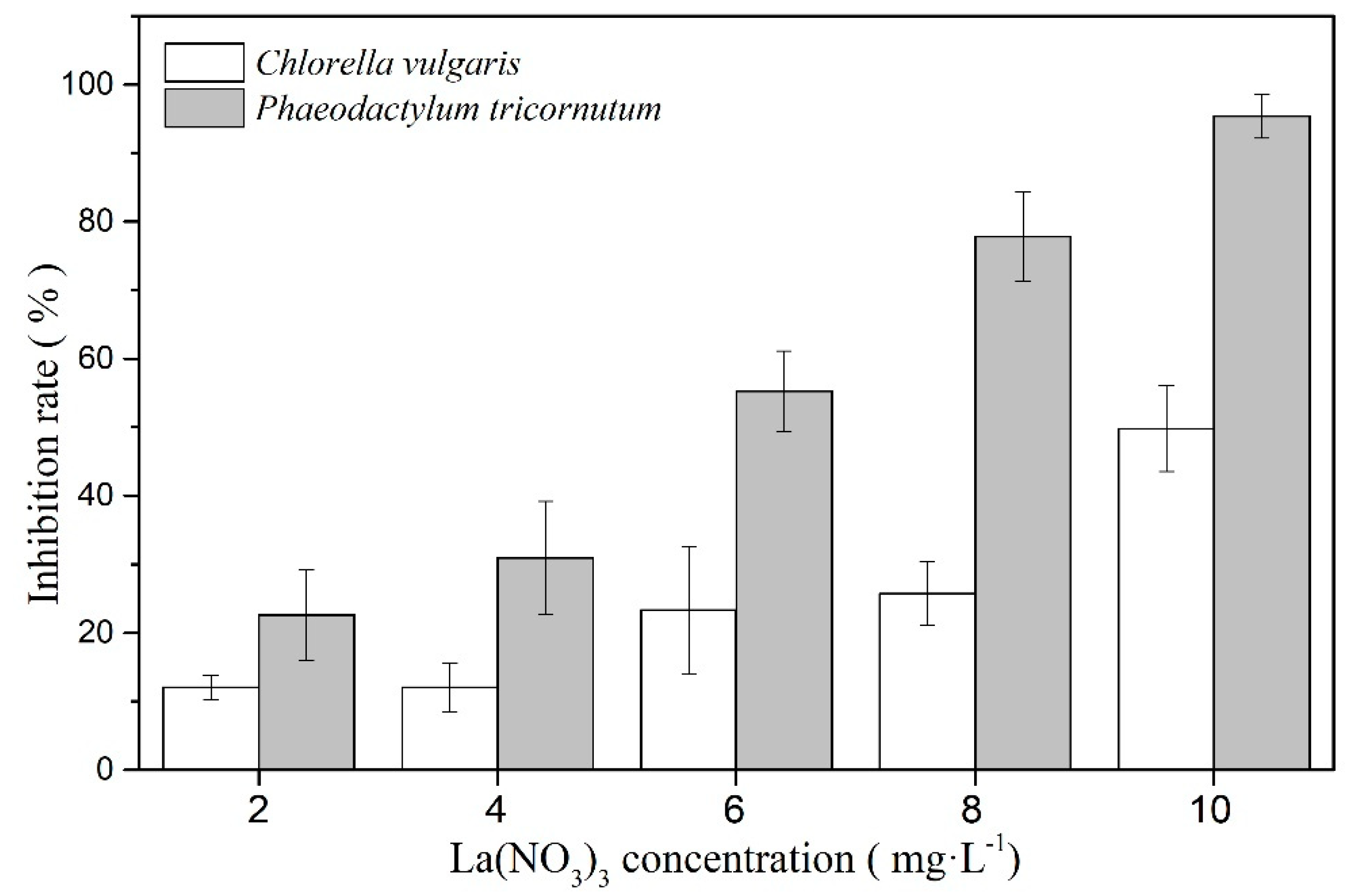
3.1. The Toxicity Effect of La(NO3)3·6H2O on the Two Algae
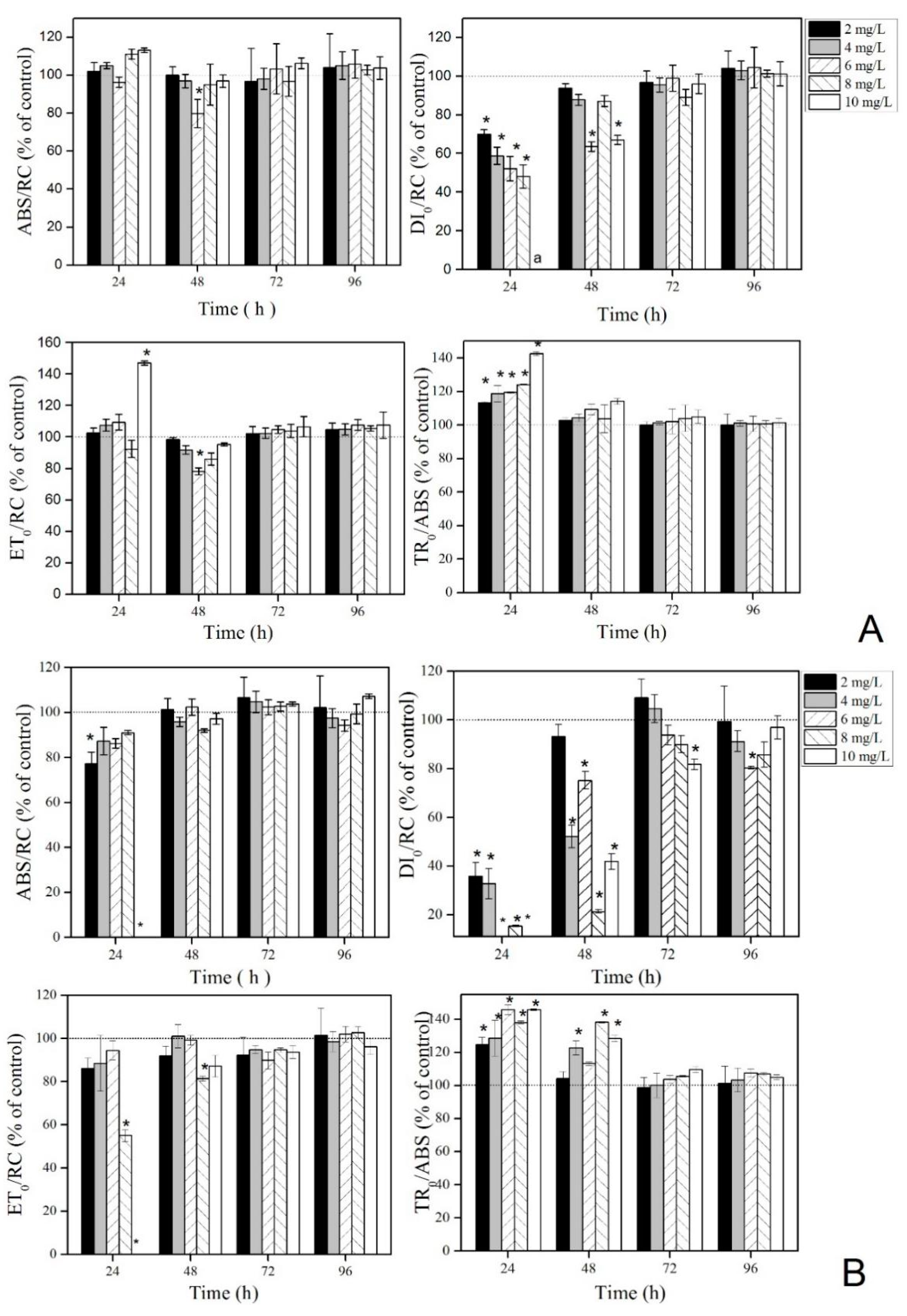
3.2. Effect of La(NO3)3·6H2O on PS II of the Two Microalgae
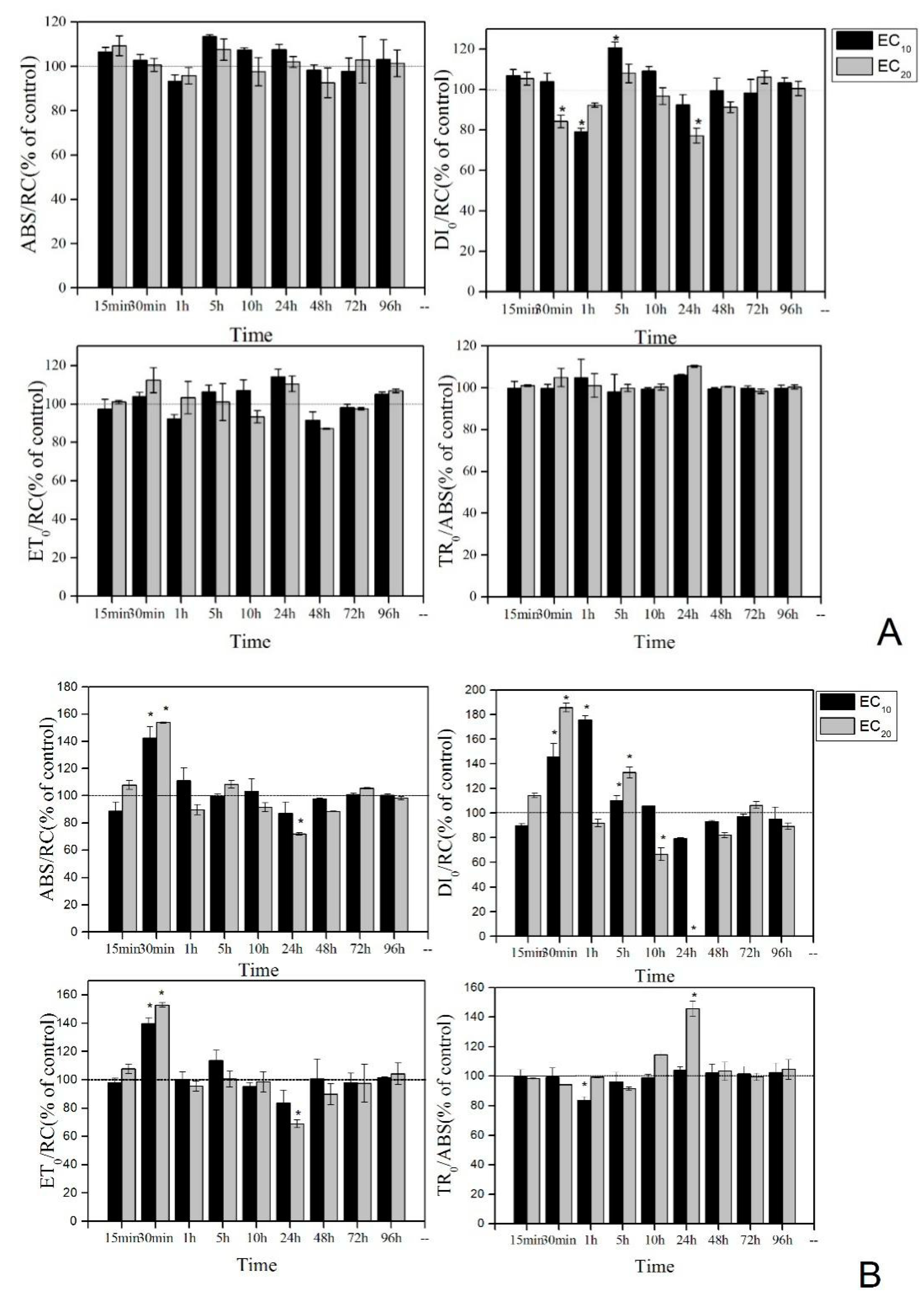
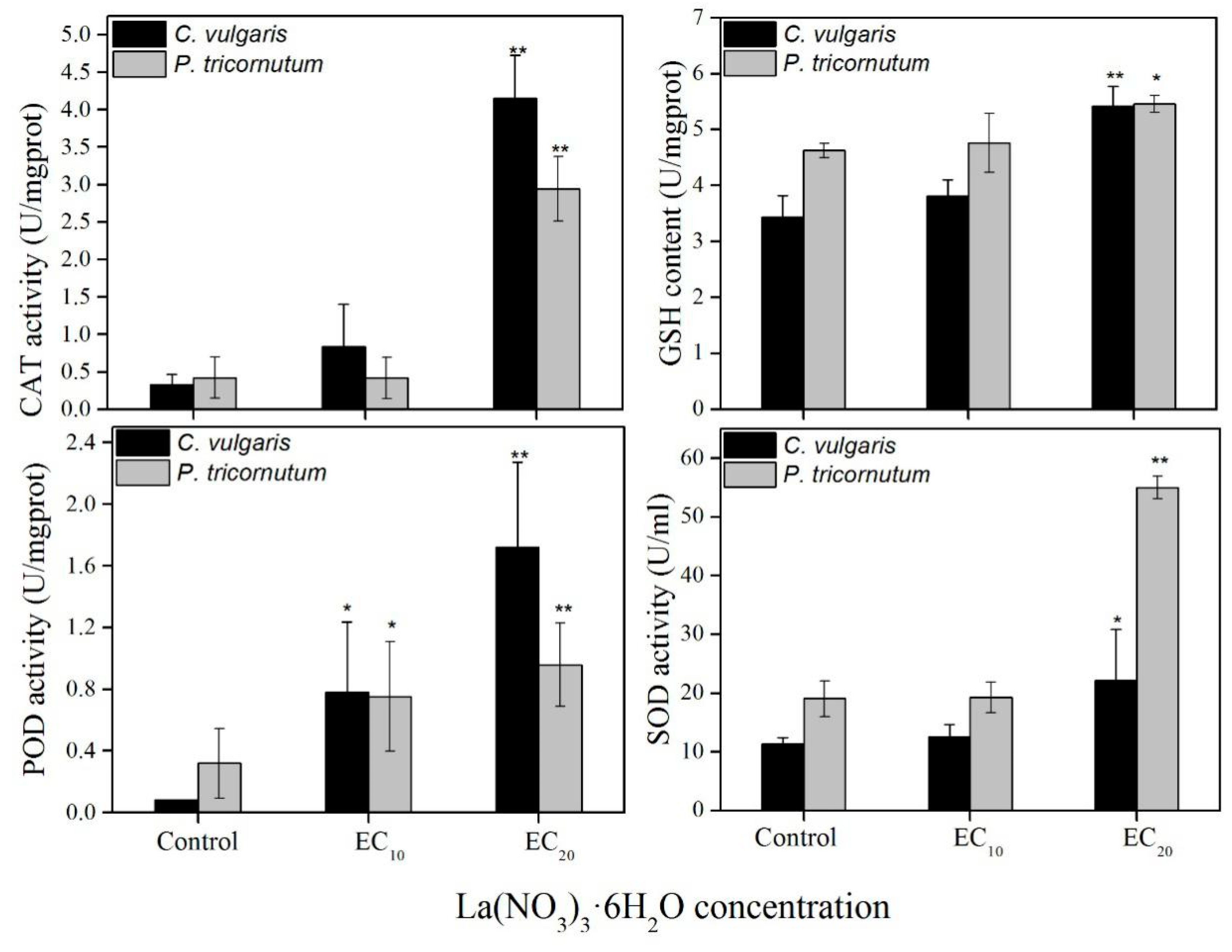
3.3. Effect of Low Concentrations (EC10 and EC20) of La(NO3)3·6H2O on PS Ⅱ and Antioxidant Enzyme System
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yang, Z.; Woolley, A. Carbonatites in China: A review. J. Asian Earth Sci. 2006, 27, 559–575. [Google Scholar] [CrossRef]
- Packey, D.J. Interesting Developments in the Rare Earth Market. Econ. Pap. A J. Appl. Econ. Policy 2013, 32, 454–461. [Google Scholar] [CrossRef]
- Jin, X.; Chu, Z.; Yan, F.; Zeng, Q. Effects of lanthanum(III) and EDTA on the growth and competition of Microcystis aeruginosa and Scenedesmus quadricauda. Limnol. Ecol. Manag. Inland Waters 2009, 39, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.G.; Ni, S.J.; Wang, J.S.; Zuo, R.; Yang, J. A geochemical survey of trace elements in agricultural and non-agricultural topsoil in Dexing area, China. J. Geochem. Explor. 2010, 104, 118–127. [Google Scholar] [CrossRef]
- Wang, L.; Zhong, B.; Liang, T.; Xing, B.; Zhu, Y. Atmospheric thorium pollution and inhalation exposure in the largest rare earth mining and smelting area in China. Sci. Total Environ. 2016, 572, 1–8. [Google Scholar] [CrossRef]
- Wang, L.; Liang, T. Accumulation and fractionation of rare earth elements in atmospheric particulates around a mine tailing in Baotou, China. Atmos. Environ. 2014, 88, 23–29. [Google Scholar] [CrossRef]
- Dai, S.; Xie, P.; Jia, S.; Ward, C.R.; Hower, J.C.; Yan, X.; French, D. Enrichment of U-Re-V-Cr-Se and rare earth elements in the Late Permian coals of the Moxinpo Coalfield, Chongqing, China: Genetic implications from geochemical and mineralogical data. Ore Geol. Rev. 2017, 80, 1–17. [Google Scholar] [CrossRef]
- Li, J.X.; Sun, C.J.; Zheng, L.; Yin, X.F.; Chen, J.H.; Jiang, F.H. Geochemical characteristics of rare earth elements in the surface sediments from the Spratly Islands of China. Mar. Pollut. Bull. 2017, 114, 1103–1109. [Google Scholar] [CrossRef]
- Yuan, J.F.; Mao, X.M.; Wang, Y.X.; Deng, Z.D.; Huang, L.H. Geochemistry of rare-earth elements in shallow groundwater, northeastern Guangdong Province, China. Acta Geochim. 2014, 33, 53–64. [Google Scholar] [CrossRef]
- Zhuang, M.; Zhao, J.; Li, S.; Liu, D.; Wang, K.; Xiao, P.; Yu, L.; Jiang, Y.; Song, J.; Zhou, J.; et al. Concentrations and health risk assessment of rare earth elements in vegetables from mining area in Shandong, China. Chemosphere 2016, 168, 578. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.; Nie, H.; Shao, L.; Wang, G.; Liu, Y. Residual levels of rare earth elements in freshwater and marine fish and their health risk assessment from Shandong, China. Mar. Pollut. Bull. 2016, 107, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yan, L.; Huo, W.; Lu, Q.; Cheng, Z.; Zhang, J.; Li, Z. Rare earth elements and hypertension risk among housewives: A pilot study in Shanxi Province, China. Environ. Pollut. 2016, 220, 837. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Li, Y.; Li, H.; Wei, B.; Liao, X.; Liang, T.; Yu, J. Levels of rare earth elements, heavy metals and uranium in a population living in Baiyun Obo, Inner Mongolia, China: A pilot study. Chemosphere 2015, 128, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Ding, Z.; Chen, Y.; Wang, X.; Dai, L. Bioaccumulation of lanthanum and cerium and their effects on the growth of wheat (Triticum aestivum L) seedlings. Chemosphere 2002, 48, 621–629. [Google Scholar]
- Zeng, Q.; Zhu, J.G.; Cheng, H.L.; Xie, Z.B.; Chu, H.Y. Phytotoxicity of lanthanum in rice in haplic acrisols and cambisols. Ecotoxicol. Environ. Saf. 2006, 64, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Ikka, T.; Kimura, K.; Yasuda, O.; Koyama, H. Characterisation of lanthanum toxicity for root growth of Arabidopsis thaliana from the aspect of natural genetic variation. Funct. Plant Biol. 2007, 34, 984–994. [Google Scholar] [CrossRef]
- Wang, L.H.; Wang, W.; Zhou, Q.; Huang, X.H. Combined effects of lanthanum (III) chloride and acid rain on photosynthetic parameters in rice. Chemosphere 2014, 112, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, M.P.; Fasciano, C.; d’Aquino, L.; Tommasi, F. Responses of antioxidant systems to lanthanum nitrate treatments in tomato plants during drought stress. Plant Biosyst. 2011, 145, 248–252. [Google Scholar] [CrossRef]
- Perl-Treves, R.; Perl, A. Oxidative Stress: An Introduction. In Oxidative Stress in Plants; Inzé, D., Van Montagu, M., Eds.; Taylor & Francis: London, UK, 2002; pp. 1–32. [Google Scholar]
- Li, Q.; Lu, Y.; Shi, Y.; Wang, T.; Ni, K.; Xu, L.; Liu, S.; Wang, L.; Xiong, Q.; Giesy, J.P.; et al. Combined effects of cadmium and fluoranthene on germination, growth and photosynthesis of soybean seedlings. J. Environ. Sci. 2013, 25, 1936–1946. [Google Scholar] [CrossRef]
- Jiang, H.X.; Chen, L.S.; Zheng, J.G.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on Photosystem II photochemistry in Citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 28, 1863–1871. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation & Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 445–483. [Google Scholar]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Lei, A.; Hu, Z.; Wong, Y.; Tam, N.F. Antioxidant responses of microalgal species to pyrene. J. Appl. Phycol. 2006, 18, 67–78. [Google Scholar] [CrossRef]
- Yu, B.; Zhao, C.Y.; Li, J.; Li, J.Y.; Peng, G. Morphological, physiological, and biochemical responses of Populus euphratica to soil flooding. Photosynthetica 2015, 53, 110–117. [Google Scholar] [CrossRef]
- Aderemi, A.O.; Novais, S.C.; Mfl, L.; Alves, L.M.; Hunter, C.; Pahl, O. Oxidative stress responses and cellular energy allocation changes in microalgae following exposure to widely used human antibiotics. Aquat. Toxicol. 2018, 203, 130. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.T.; Wong, Y.S.; Tam, N.F.Y. Antioxidant responses of different microalgal species to nonylphenol-induced oxidative stress. J. Appl. Phycol. 2017, 29, 1317–1329. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, Z.; Zhi, W. Oxidative stress responses and toxin accumulation in the freshwater snail Radix swinhoei (Gastropoda, Pulmonata) exposed to microcystin-LR. Environ. Sci. Pollut. Res. 2016, 23, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Tai, P.; Zhao, Q.; Su, D.; Li, P.; Stagnitti, F. Biological toxicity of lanthanide elements on algae. Chemosphere 2010, 80, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Rackova, L.; Paganova, V.; Swoczyna, T.; Rusinowski, S.; Sitko, K. Can chlorophyll-a fluorescence parameters be used as bio-indicators to distinguish between drought and salinity stress in Tilia cordata Mill? Environ. Exp. Bot. 2018, 152, 149–157. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Montavon, P.; Bortlik, K. Evolution of Robusta Green Coffee Redox Enzymatic Activities with Maturation. J. Agric. Food Chem. 2004, 52, 3590–3594. [Google Scholar] [CrossRef] [PubMed]
- Peskin, A.V.; Winterbourn, C.C. A microtiter plate assay for superoxide dismutase using a water-soluble tetrazolium salt (WST-1). Clin. Chim. Acta 2000, 293, 157–166. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, M.; Lin, K.F.; Sun, W.F.; Xiong, B.; Guo, M.J.; Cui, X.H.; Fu, R.B. Eco-toxicological effect of Carbamazepine on Scenedesmus obliquus and Chlorella pyrenoidosa. Environ. Toxicol. Pharmacol. 2012, 33, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Góth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol. 1985, 113, 548–555. [Google Scholar] [PubMed]
- Joonas, E.; Aruoja, V.; Olli, K.; Syvertsen-Wiig, G.; Vija, H.; Kahru, A. Potency of (doped) rare earth oxide particles and their constituent metals to inhibit algal growth and induce direct toxic effects. Sci. Total Environ. 2017, 593, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Singh-Tomar, R.; Jajoo, A. Alteration in PS II heterogeneity under the influence of polycyclic aromatic hydrocarbon (fluoranthene) in wheat leaves (Triticum aestivum). Plant Sci. 2013, 209, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Bascik-Remisiewicz, A.; Aksmann, A.; Zak, A.; Kowalska, M.; Tukaj, Z. Toxicity of Cadmium, Anthracene, and Their Mixture to Desmodesmus subspicatus Estimated by Algal Growth-Inhibition ISO Standard Test. Arch. Environ. Contam. Toxicol. 2011, 60, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Increases in the fluorescence Fo level and reversible inhibition of Photosystem II reaction center by high-temperature treatments in higher plants. Photosynth. Res. 1997, 52, 57–64. [Google Scholar] [CrossRef]
- Perron, M.C.; Juneau, P. Effect of endocrine disrupters on photosystem II energy fluxes of green algae and cyanobacteria. Environ. Res. 2011, 111, 520–529. [Google Scholar] [CrossRef]
- Song, Y.G.; Liu, B.; Wang, L.F.; Li, M.H.; Liu, Y. Damage to the oxygen-evolving complex by superoxide anion, hydrogen peroxide, and hydroxyl radical in photoinhibition of photosystem II. Photosynth. Res. 2006, 90, 67–78. [Google Scholar] [CrossRef]
- Zhuang, L.E.; Zhao, L.; Yin, P.H. Combined algicidal effect of urocanic acid, N-acetylhistamine and l-histidine to harmful alga Phaeocystis globosa. RSC Adv. 2018, 8, 12760–12766. [Google Scholar] [CrossRef]
- Tan, S.; Hu, X.L.; Yin, P.H.; Zhao, L. Photosynthetic inhibition and oxidative stress to the toxic Phaeocystis globosa caused by a diketopiperazine isolated from products of algicidal bacterium metabolism. J. Microbiol. 2016, 54, 364–375. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Terms | Illustration |
|---|---|
| F0 | Minimal recorded fluorescence intensity |
| Fm | Maximal recorded fluorescence intensity |
| F50 μs,F2 ms | Fluorescence intensities at 50 μs and 2 ms |
| OJIP | O phase (F20 μs/F0), J phase (F2 ms), I phase (F30 ms) and P phase (Fm) |
| Vj | Relative variable fluorescence intensity at the J-step |
| M0 | Approximated initial slope of the fluorescence transient |
| RC | Reaction center of photosystem II |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, D.; He, N.; Chen, Q.; Duan, S. Effects of Lanthanum on the Photosystem II Energy Fluxes and Antioxidant System of Chlorella Vulgaris and Phaeodactylum Tricornutum . Int. J. Environ. Res. Public Health 2019, 16, 2242. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16122242
Sun D, He N, Chen Q, Duan S. Effects of Lanthanum on the Photosystem II Energy Fluxes and Antioxidant System of Chlorella Vulgaris and Phaeodactylum Tricornutum . International Journal of Environmental Research and Public Health. 2019; 16(12):2242. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16122242
Chicago/Turabian StyleSun, Dong, Ning He, Qi Chen, and Shunshan Duan. 2019. "Effects of Lanthanum on the Photosystem II Energy Fluxes and Antioxidant System of Chlorella Vulgaris and Phaeodactylum Tricornutum " International Journal of Environmental Research and Public Health 16, no. 12: 2242. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16122242