Assessing the Spatial Distribution of Biodiversity in a Changing Temperature Pattern: The Case of Catalonia, Spain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Setting
2.2. Variables and Information Sources
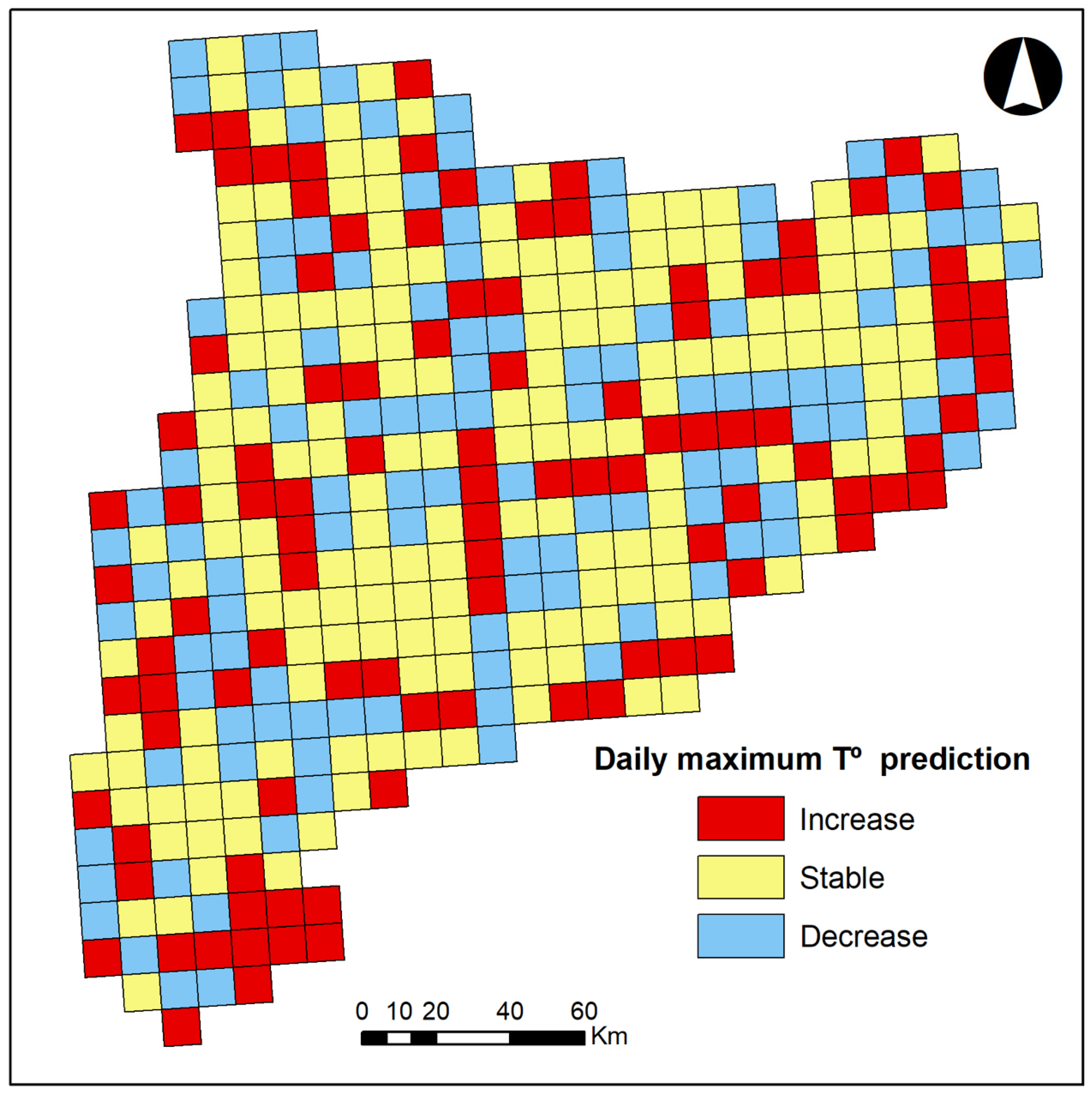
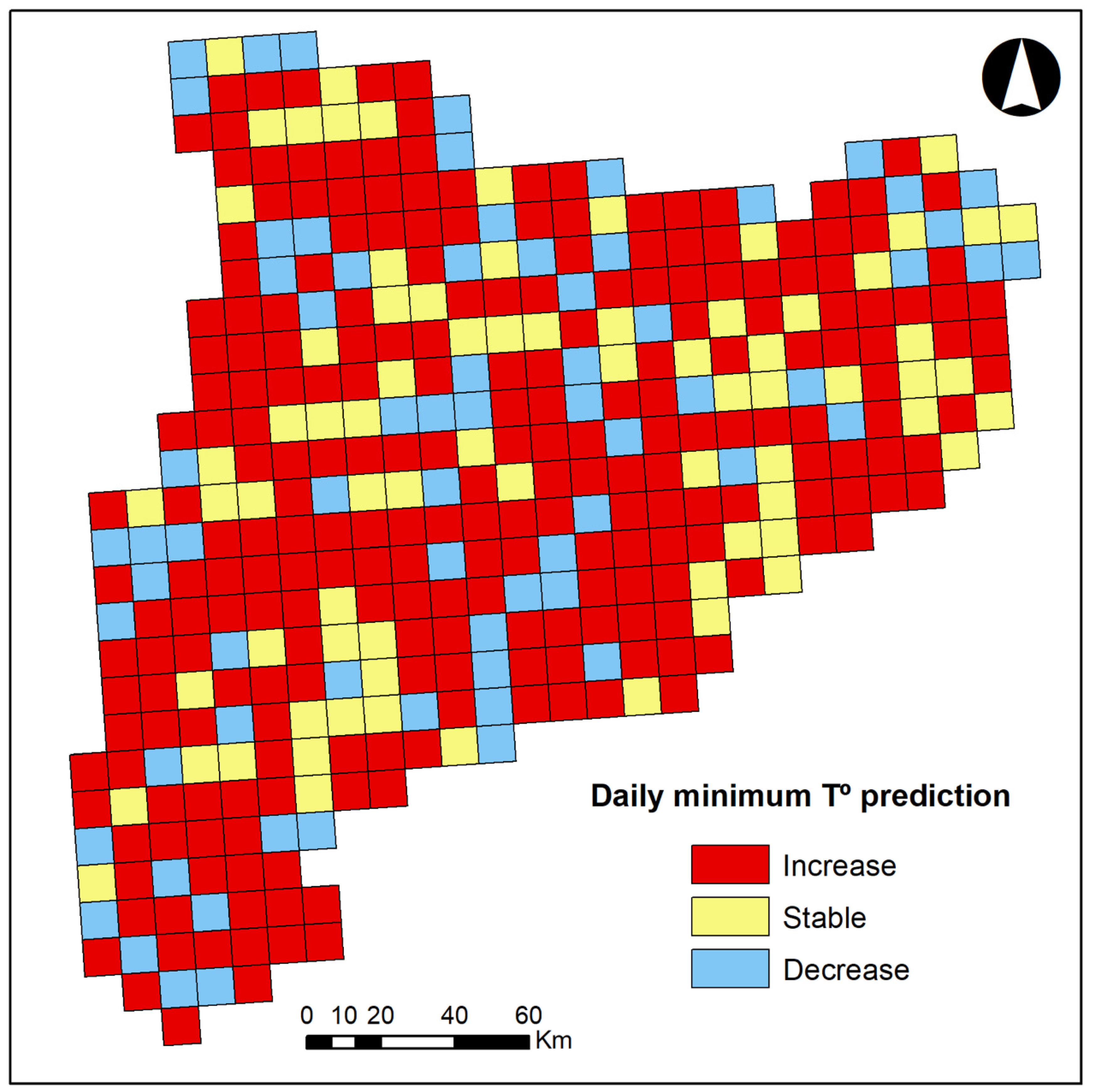
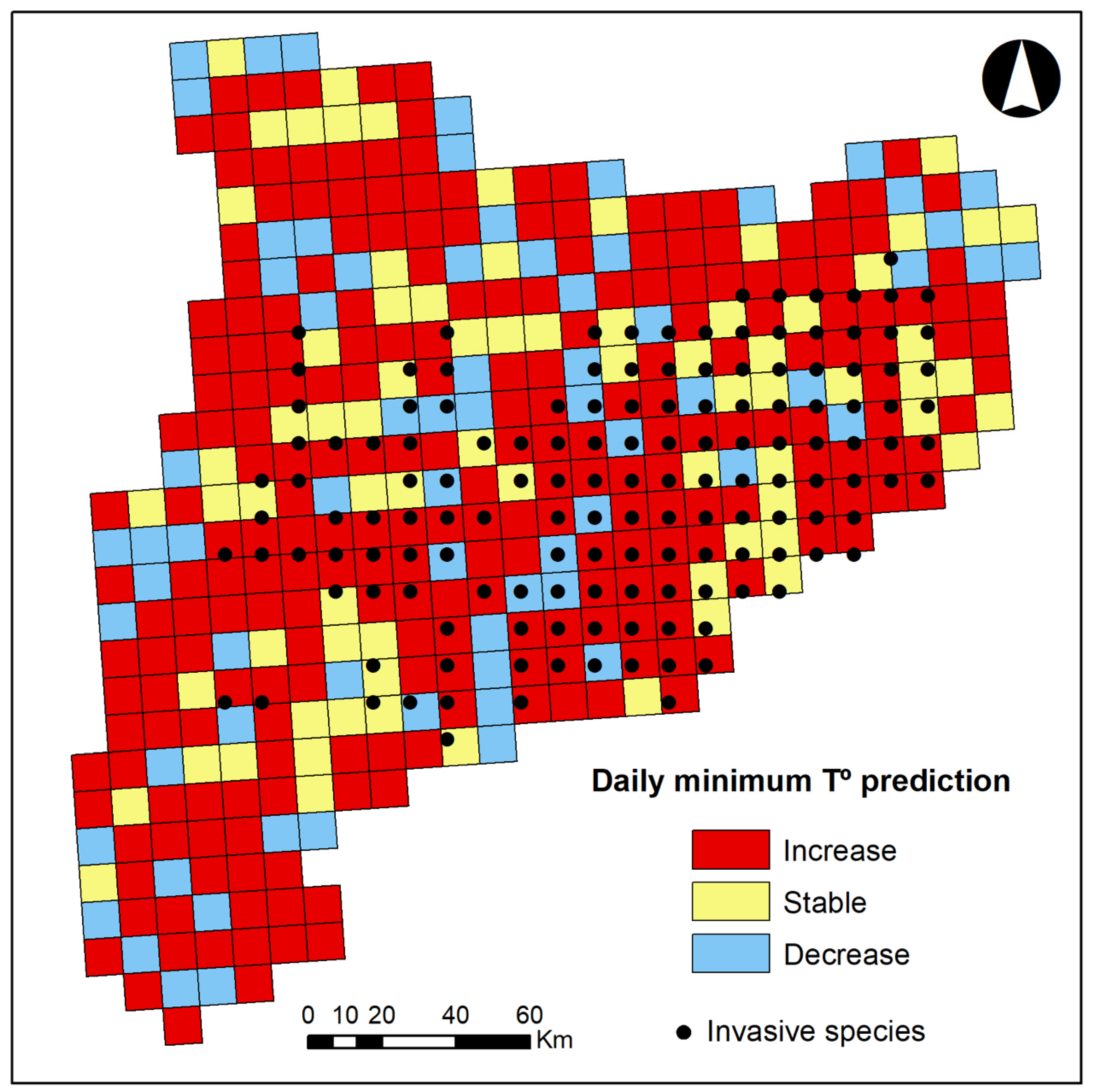
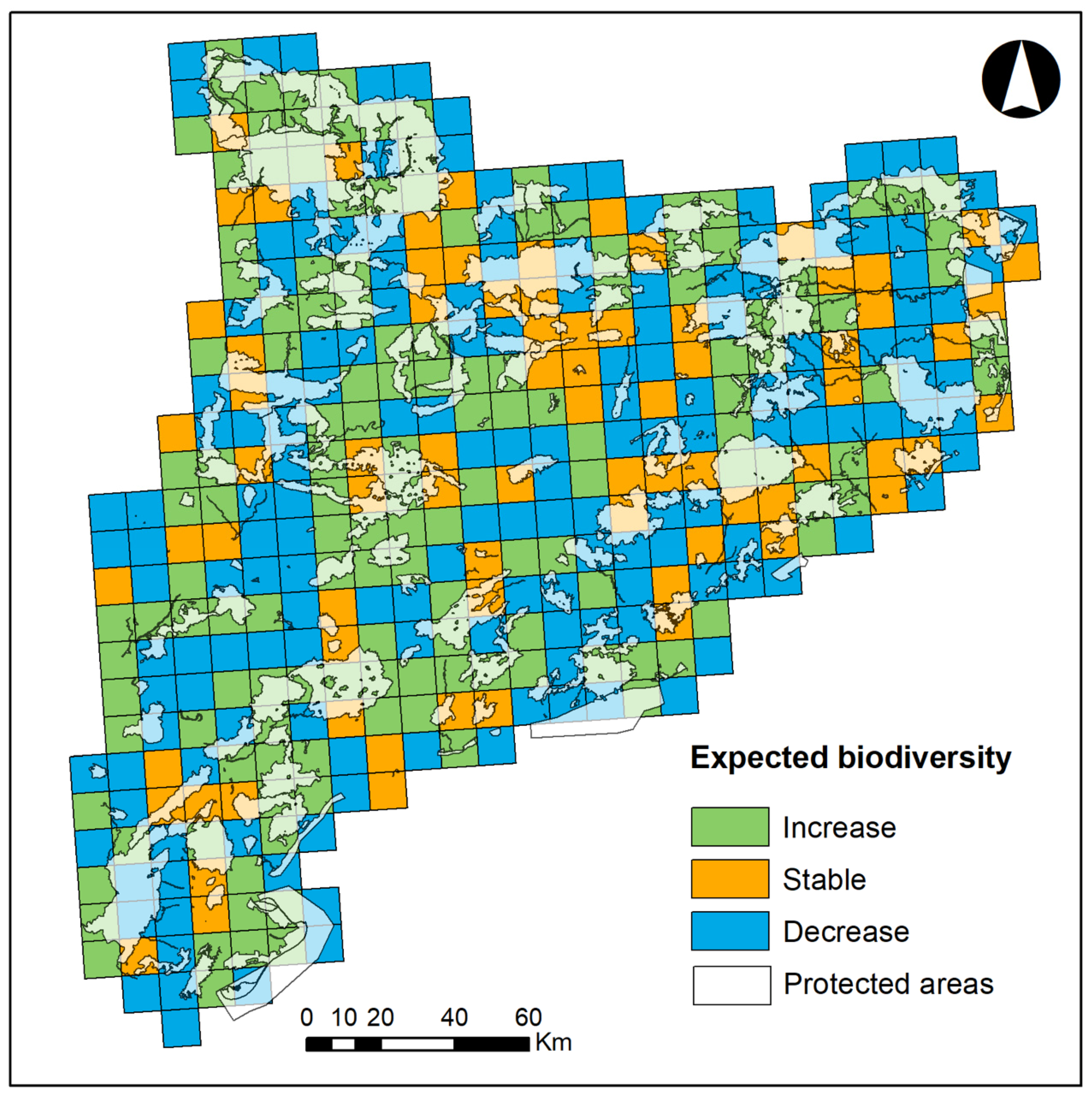
2.2.1. Response Variables
2.2.2. Explanatory Variables
2.3. Statistical Analysis
Spatial Adjustment
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Declaration
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Relative Risk | 95% Credibility Interval | |
|---|---|---|---|
| Variations in the temperature (°C) | |||
| Maximum temperature | 1.275 | 0.698 | 2.322 |
| Minimum temperature | 0.933 | 0.493 | 1.767 |
| Hill shade and elevation [1st Quintile < 161 m] | |||
| 2nd Quintile 161–334 m | 0.967 | 0.869 | 1.074 |
| 3rd Quintile 335–608 m | 0.945 | 0.826 | 1.079 |
| 4th Quintile 609–1069 m | 0.968 | 0.825 | 1.132 |
| 5th Quintile 1070–2541 m | 0.968 | 0.801 | 1.166 |
| Distance to anthropic areas [1st Quintile < 180 m] | |||
| 2nd Quintile 180–539 m | 0.948 | 0.867 | 1.036 |
| 3rd Quintile 540–905 m | 0.991 | 0.907 | 1.082 |
| 4th Quintile 906–1664 m | 0.976 | 0.891 | 1.068 |
| 5th Quintile 1665–10170 m | 0.905 | 0.822 | 0.997 |
| Slope [1st Quintile < 0.37%] | |||
| 2nd Quintile | 1.147 | 1.047 | 1.255 |
| 3rd Quintile | 1.056 | 0.947 | 1.178 |
| 4th Quintile 7.4%–11.0% | 1.118 | 1.004 | 1.245 |
| 5th Quintile 11.0%–24.4% | 1.129 | 1.001 | 1.276 |
| Aspect [1st Quartile < 93.0] | |||
| 2nd Quartile 93.0–174.0 | 1.025 | 0.941 | 1.115 |
| 3rd Quartile 174.1–257.0 | 0.993 | 0.919 | 1.072 |
| 4th Quartile 257.1–357.0 | 0.936 | 0.848 | 1.032 |
| Land use [Other] | |||
| Coniferous forests | |||
| 1997 Cover map | 0.948 | 0.000 | 51,87,048.404 |
| 2002 Cover map | 1.075 | 0.000 | 5,887,435.884 |
| 2010 Cover map | 1.486 | 0.881 | 2.512 |
| Dense forest | |||
| 1997 Cover map | 1.077 | 0.000 | 5,897,531.078 |
| 2002 Cover map | 0.914 | 0.000 | 5,005,564.082 |
| 2010 Cover map | 1.384 | 0.824 | 2.330 |
| Fruit trees and berries | |||
| 1997 Cover map | 1.032 | 0.000 | 5,647,790.248 |
| 2002 Cover map | 1.000 | 0.000 | 5,474,699.312 |
| 2010 Cover map | 1.420 | 0.853 | 2.366 |
| Artificial, non-agricultural vegetated | |||
| 1997 Cover map | 0.911 | 0.000 | 4,993,598.489 |
| 2002 Cover map | 1.214 | 0.000 | 6,652,178.514 |
| 2010 Cover map | 1.266 | 0,000 | 6,928,848.938 |
| Transitional woodland scrub | |||
| 1997 Cover map | 0.988 | 0.000 | 5,404,510.383 |
| 2002 Cover map | 1.061 | 0.000 | 5,808,358.914 |
| 2010 Cover map | 1.448 | 0.863 | 2.433 |
| Natural grassland | |||
| 1997 Cover map | 1.256 | 0.000 | 6,905,193.758 |
| 2002 Cover map | 0.977 | 0.000 | 5,373,630.141 |
| 2010 Cover map | 1.276 | 0.757 | 2.152 |
| Mixed forest | |||
| 1997 Cover map | 1.418 | 0.000 | 7,762,645.290 |
| 2002 Cover map | 1.004 | 0.000 | 5,502,696.848 |
| 2010 Cover map | 1.522 | 0.905 | 2.567 |
References
- Vicente-Serrano, S.M.; López-Moreno, J.I.; Beguería, S.; Lorenzo-Lacruz, J.; Sánchez-Lorenzo, A.; García-Ruíz, J.M.; Azorín-Molina, C.; Morán-Tejeda, E.; Revuelto, J.; Trigo, R.; et al. Evidence of increasing drought severity caused by temperature rise in southern Europe. Environ. Res. Lett. 2014, 9, 044001. [Google Scholar] [CrossRef]
- Field, C.B.; Barros, V.R.; Dokken, D.J.; Mach, K.J.; Mastrandrea, M.D.; Bilir, T.E.; Chatterjee, M.; Ebi, K.L.; Estrada, Y.O.; Genova, R.C.; et al. (Eds.) IPCC, 2014: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- McMahon, S.M.; Harrison, S.P.; Armbruster, W.S.; Bartlein, P.J.; Beale, C.M.; Edwards, M.E.; Kattge, J.; Midgley, G.; Morin, X.; Prentice, I.C. Improving assessment and modelling of climate change impacts on global terrestrial biodiversity. Trends Ecol. Evol. 2011, 26, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Iwamura, T.; Guisan, A.; Wilson, K.A.; Possingham, H.P. How robust are global conservation priorities to climate change? Glob. Environ. Chang. 2013, 23, 1277–1284. [Google Scholar] [CrossRef]
- Kittel, T.G.F. The Vulnerability of Biodiversity to Rapid Climate Change. In Vulnerability of Ecosystems to Climate; Seastedt, T.R., Suding, K., Eds.; Elsevier Inc.: Oxford, UK; Academic Press: Cambridge, UK, 2013; Volume 4, Chapter 15; pp. 185–201. [Google Scholar]
- Perrings, C.; Mäler, K.G.; Folke, C.; Holling, C.; Jansson, B.O. (Eds.) Biodiversity Loss: Economic and Ecological Issues; Cambridge University Press: Cambridge, UK, 1995; p. 332. [Google Scholar]
- Díaz, S.; Fargione, J.; Chapin, F.S.; Tilman, D. Biodiversity loss threatens human well-being. PLoS Biol. 2006, 4, e277. [Google Scholar] [CrossRef]
- Pimm, S.L.; Raven, P. Biodiversity: Extinction by numbers. Nature 2000, 403, 843–845. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pysek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef]
- Kovats, S.; Valentini, R.; Bouwer, L.M.; Georgopoulou, E.; Jacob, D.; Martin, E.; Rounsevell, M.; Soussana, J.F. Chapter 23. In IPCC, 2014: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Wright, S. Correlation and causation. J. Agric. Res. 1921, 20, 557–585. [Google Scholar]
- Veldkamp, A.; Lambin, E.F. Predicting land-use change. Agric. Ecosyst. Environ. 2001, 85, 1–6. [Google Scholar] [CrossRef]
- Anderson, J.T.; Panetta, A.M.; Mitchell-Olds, T. Evolutionary and ecological responses to anthropogenic climate change: Update on anthropogenic climate change. Plant Physiol. 2012, 160, 1728–1740. [Google Scholar] [CrossRef]
- Klausmeyer, K.R.; Shaw, M.R. Climate change, habitat loss, protected areas and the climate adaptation potential of species in Mediterranean ecosystems worldwide. PLoS ONE 2009, 4, e6392. [Google Scholar] [CrossRef] [PubMed]
- Ahola, M.P.; Laaksonen, T.; Eeva, T. Climate change can alter competitive relationships between resident and migratory birds. J. Anim. Ecol. 2007, 76, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-Mclane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Sala, O.; Chapin, F.; Armesto, J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Hansen, A.J.; Neilson, R.P.; Dale, V.H.; Flather, C.H.; Iverson, L.R.; Currie, D.J.; Shafer, S.; Cook, R.; Bartlein, P.J. Global change in forests: Responses of species, communities, and biomes interactions between climate change and land use are projected to cause large shifts in biodiversity. Bioscience 2001, 51, 765–779. [Google Scholar] [CrossRef]
- Brooker, R.W.; Travis, J.M.; Clark, E.J.; Dytham, C. Modelling species’ range shifts in a changing climate: The impacts of biotic interactions, dispersal distance and the rate of climate change. J. Theor. Biol. 2007, 245, 59–65. [Google Scholar] [CrossRef]
- Mawdsley, J.R.; O’Malley, R.; Ojima, D.S. A Review of Climate-Change Adaptation Strategies for Wildlife Management and Biodiversity Conservation. Conserv. Biol. 2009, 23, 1080–1089. [Google Scholar] [CrossRef]
- Perrings, C.; Duraiappah, A.; Larigauderie, A.; Mooney, H. The biodiversity and ecosystem services science-policy interface. Science 2011, 331, 1139–1140. [Google Scholar] [CrossRef]
- Lepers, E.; Lambin, E.F.; Janetos, A.C.; Defries, R.; Achard, F.; Ramankutty, N.; Scholes, R.J. A synthesis of information on rapid land-cover change for the period 1081–2000. Bioscience 2005, 55, 115–124. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2011, 14, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, L08707. [Google Scholar] [CrossRef]
- Maiorano, L.; Falcucci, A.; Zimmermann, N.E.; Psomas, A.; Pottier, J.; Baisero, D.; Rondinini, C.; Guisan, A.; Boitani, L. The future of terrestrial mammals in the Mediterranean basin under climate change. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2011, 366, 2681–2692. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, P.; Neilson, R.P.; Lenihan, J.M.; Drapek, R.J. Global patterns in the vulnerability of ecosystems to vegetation shifts due to climate change. Glob. Ecol. Biogeogr. 2010, 19, 755–768. [Google Scholar] [CrossRef]
- Serra, L.; Juan, P.; Varga, D.; Mateu, J.; Saez, M. Spatial pattern modelling of wildfires in Catalonia, Spain 2004–2008. Environ. Model. Softw. 2012, 40, 235–244. [Google Scholar] [CrossRef]
- European Parliament Resolution of 20 April 2012. Our Life Insurance, Our Natural Capital: An EU Biodiversity Strategy to 2020. Available online: http://ec.europa.eu/environment/nature/biodiversity/comm2006/pdf/EP_resolution_april2012.pdf (accessed on 3 February 2018).
- Torras, O.; Martín-Queller, E.; Saura, S. Relating landscape structure, environment and management to biodiversity indicators estimated from forest inventory data in Catalonia (NE Spain). For. Syst. 2009, 18, 322–337. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Fisheries, Food and Environment. Spanish Inventory of Natural Heritage and Biodiversity [in Spanish]. Ministry of Agriculture and Fisheries, Food and Environment, Government of Spain: Madrid, Spain, 2015. Available online: http://www.mapama.gob.es/es/biodiversidad/temas/inventarios-nacionales/inventario-espanol-patrimonio-natural-biodiv (accessed on 3 February 2018).
- Ministry of Agriculture and Fisheries, Food and Environment. Open Data. Spanish Inventory of Natural Heritage and Biodiversity [in Spanish]. Ministry of Agriculture and Fisheries, Food and Environment, Government of Spain: Madrid, Spain, 2015. Available online: http://www.mapama.gob.es/en/biodiversidad/temas/inventarios-nacionales/inventario-espanol-patrimonio-natural-biodiv/sistema-indicadores/descarga-datos-abiertos.aspx (accessed on 3 February 2018).
- Saez, M.; Barceló, M.A.; Tobías, A.; Varga, D.; Ocaña-Riola, R.; Juan, P.; Mateu, J. Space-time interpolation of daily air temperatures. J. Environ. Stat. 2012, 3. Available online: http://www.jenvstat.org/v03/i05/paper (accessed on 3 February 2018).
- Coblentz David, D.; Riitters Kurt, H. Topographic controls on the regional-scale biodiversity of the south-western USA. J. Biogeogr. 2004, 31, 1125–1138. [Google Scholar] [CrossRef]
- Badgley, C.; Smiley, T.M.; Terry, R.; Davis, E.B.; DeSantis, L.R.; Fox, D.L.; Hopkins, S.S.; Jezkova, T.; Matocq, M.D.; Matzke, N.; et al. Biodiversity and topographic complexity: Modern and geohistorical perspectives. Trends Ecol. Evol. 2017, 32, 211–226. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45. [Google Scholar] [CrossRef] [PubMed]
- Cartographic and Geological Institute of Catalonia. Available online: http://www.icgc.cat/en/ (accessed on 3 February 2018).
- Röder, A.; Udelhoven, T.; Hill, J.; del Barrio, G.; Tsiourlis, G. Trend analysis of Landsat-TM and -ETM+ imagery to monitor grazing impact in a rangeland ecosystem in Northern Greece. Remote Sens. Environ. 2008, 112, 2863–2875. [Google Scholar] [CrossRef]
- Chuvieco, E.; Aguado, I.; Yebra, Y.; Nieto, H.; Salas, J.; Martín, M.P.; Vilar, L.; Martínez, J.; Martín, S.; Ibarra, P.; et al. Development of a framework for fire risk assessment using remote sensing and geographic information system technologies. Ecol. Model. 2010, 221, 46–58. [Google Scholar] [CrossRef]
- Vidal-Macua, J.J.; Zabala, A.; Ninyerol, M.; Pons, X. Developing spatially and thematically detailed back dated maps for land cover studies. Int. J. Digit. Earth 2017, 10, 175–206. [Google Scholar] [CrossRef]
- Lindgren, F.; Rue, H.; Lindström, J. An explicit link between Gaussian fields and Gaussian Markov random fields: The stochastic partial differential equation approach (with discussion). J. R. Stat. Soc. Ser. B 2011, 73, 423–498. [Google Scholar]
- Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian inference for latent Gaussian models by using integrated nested Laplace approximations (with discussion). J. R. Stat. Soc. Ser. B 2009, 71, 319–392. [Google Scholar] [CrossRef]
- Simpson, D.P.; Rue, H.; Martins, T.G.; Riebler, A.; Sørbye, S.H. Penalising model component complexity: A principled, practical approach to constructing priors (with discussion). Stat. Sci. 2017, 32, 1–46. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. Available online: https://www.R-project.org/ (accessed on 25 November 2017).
- R INLA Project 2018. Available online: http://www.r-inla.org/home (accessed on 3 February 2018).
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varga, D.; Roigé, M.; Pintó, J.; Saez, M. Assessing the Spatial Distribution of Biodiversity in a Changing Temperature Pattern: The Case of Catalonia, Spain. Int. J. Environ. Res. Public Health 2019, 16, 4026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16204026
Varga D, Roigé M, Pintó J, Saez M. Assessing the Spatial Distribution of Biodiversity in a Changing Temperature Pattern: The Case of Catalonia, Spain. International Journal of Environmental Research and Public Health. 2019; 16(20):4026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16204026
Chicago/Turabian StyleVarga, Diego, Mariona Roigé, Josep Pintó, and Marc Saez. 2019. "Assessing the Spatial Distribution of Biodiversity in a Changing Temperature Pattern: The Case of Catalonia, Spain" International Journal of Environmental Research and Public Health 16, no. 20: 4026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16204026