Responses of Periphyton Microbial Growth, Activity, and Pollutant Removal Efficiency to Cu Exposure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Periphyton Culture
2.2. Experimental Design
2.3. Samples Analysis
3. Results and Discussion
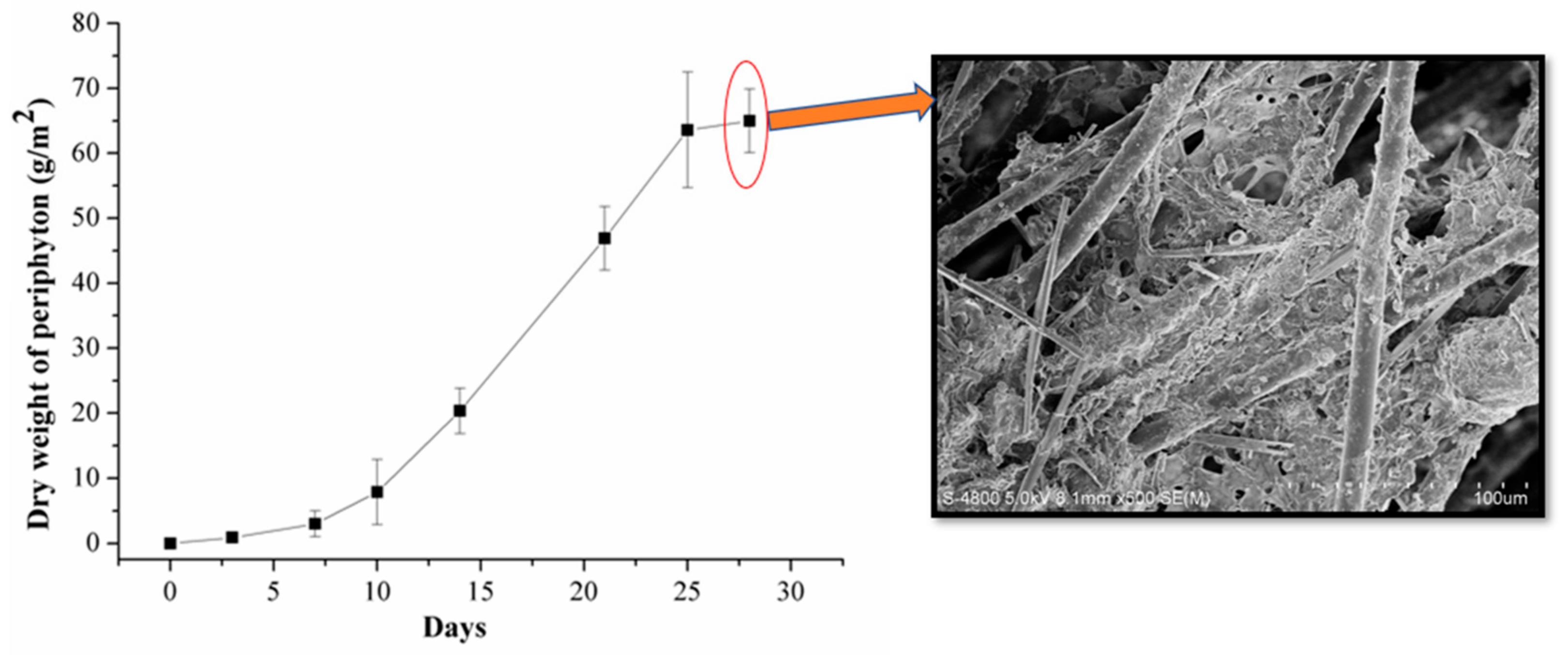
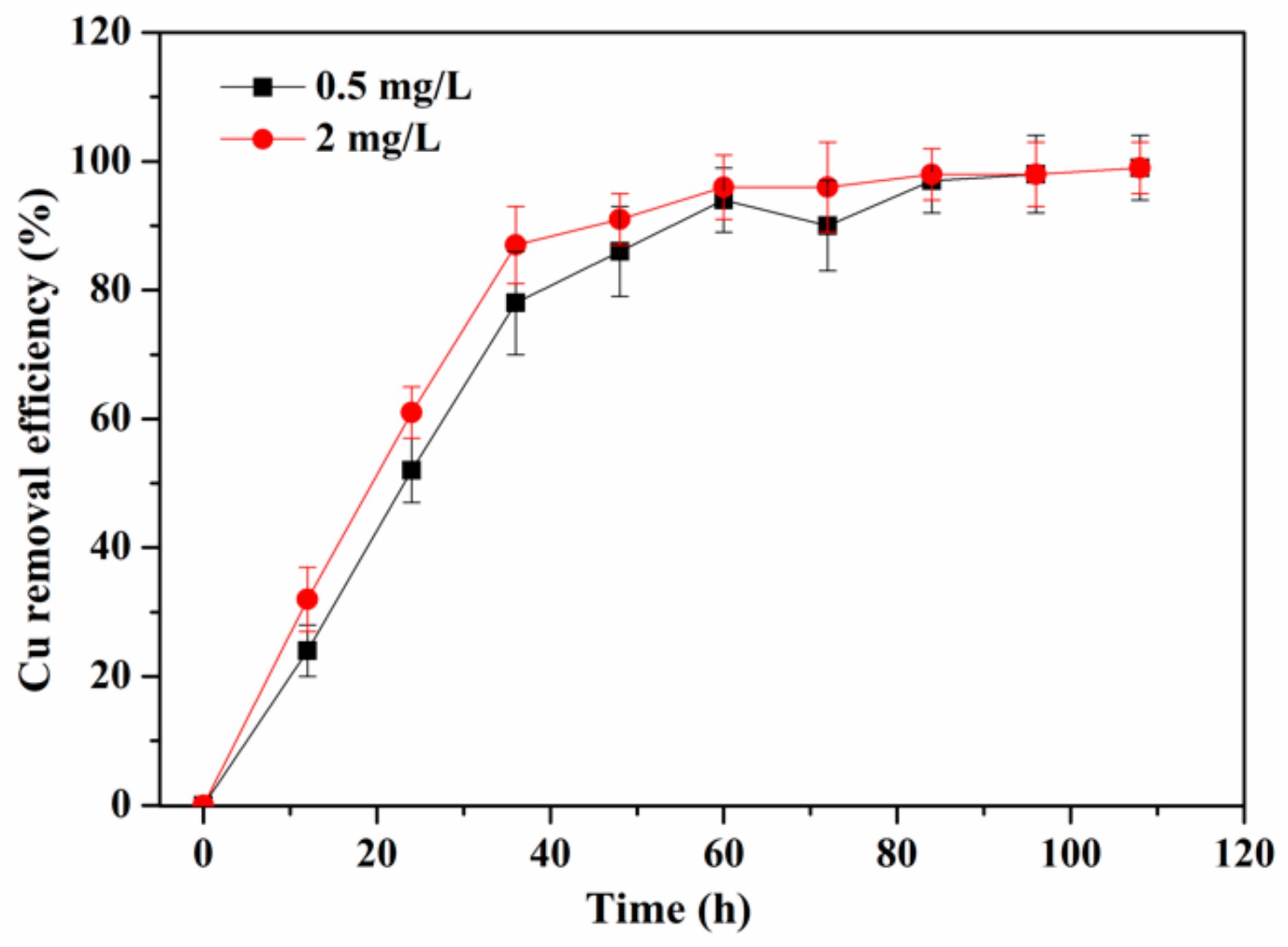
3.1. Periphyton Characterizaiton and Cu2+ Removal Efficiency
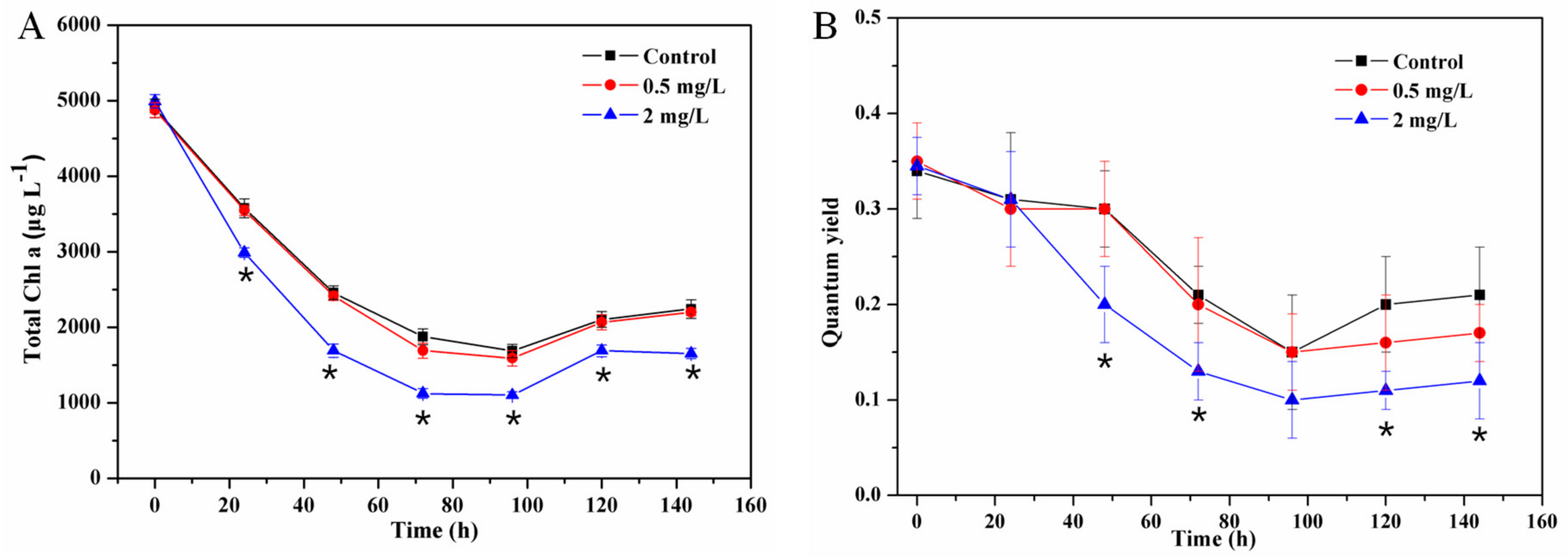
3.2. Effect of Cu2+ on Microbial Growth of Periphyton
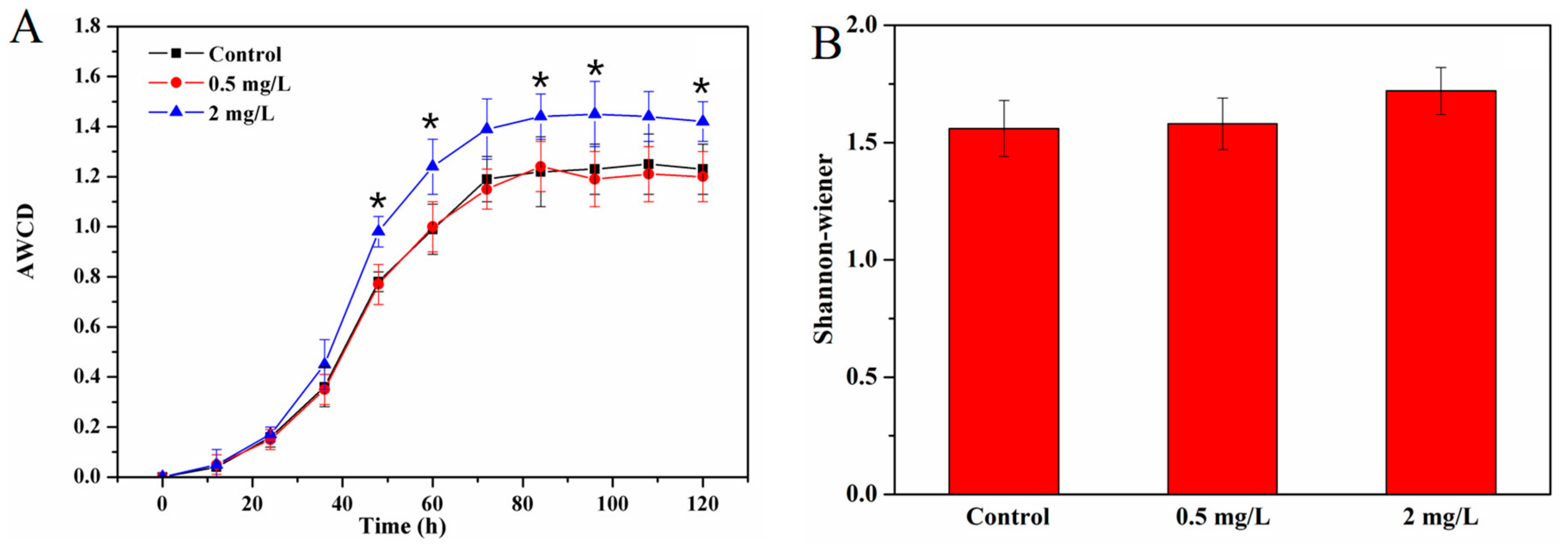
3.3. Effect of Cu2+ on Carbon Metabolism of Periphyton
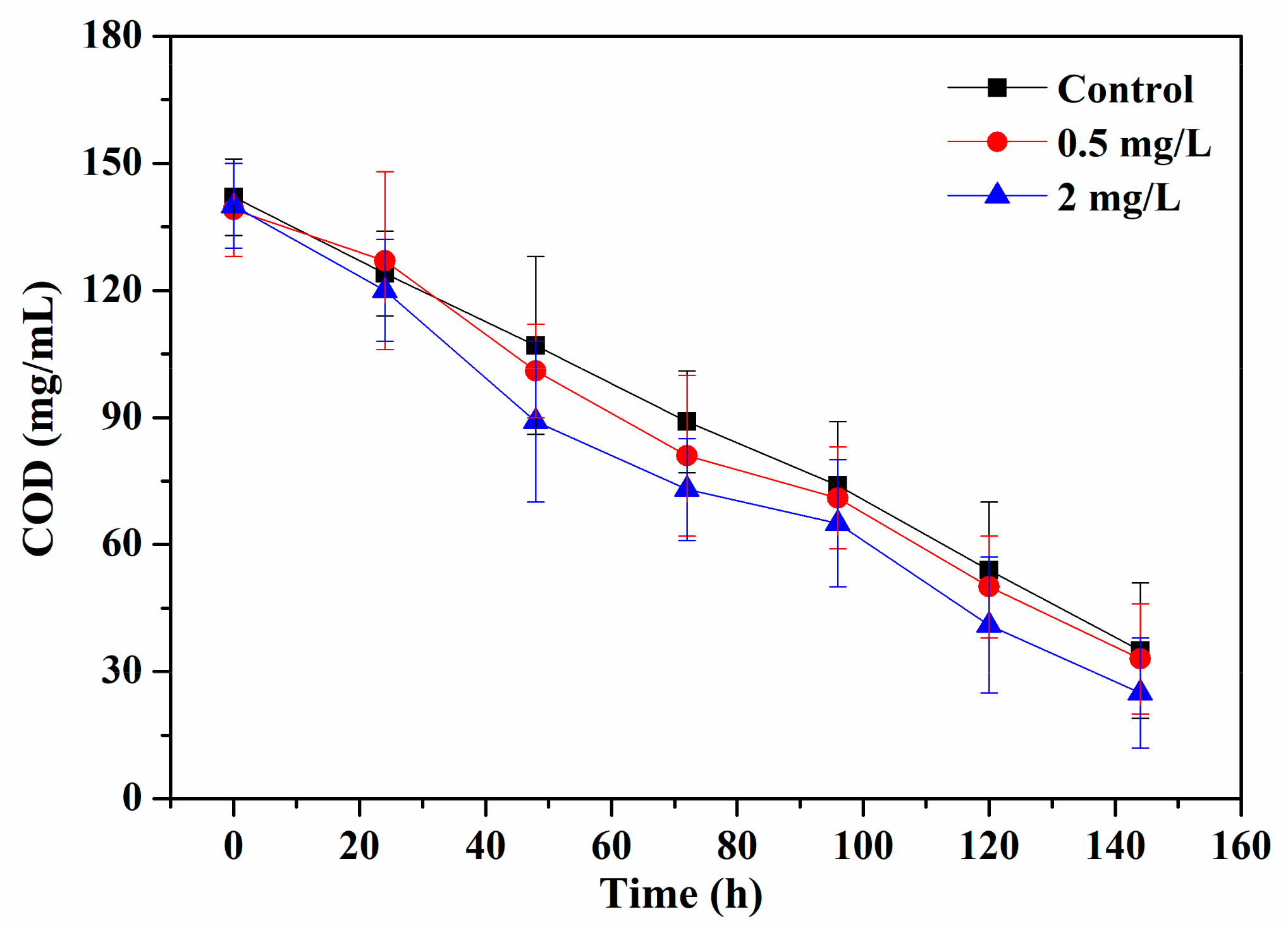
3.4. Pollutants Removal of Periphyton after Exposed to Cu2+
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Boopathy, R. Factors limiting bioremediation technologies. Bioresour. Technol. 2000, 74, 63–67. [Google Scholar] [CrossRef]
- De Lorenzo, V. Systems biology approaches to bioremediation. Curr. Opin. Biotech. 2008, 19, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.L.; Zhang, J.; Zhu, L.Z. Evaluation of the application potential of bentonites in phenanthrene bioremediation by characterizing the biofilm community. Bioresour. Technol. 2013, 134, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, G.; Xu, J.F.; Thompson, R.W.; Yang, Y.; Randol-Smith, P.; Weathers, P.J. Integrated green algal technology for bioremediation and biofuel. Bioresour. Technol. 2012, 107, 1–9. [Google Scholar] [CrossRef]
- Yang, J.L.; Liu, J.Z.; Wu, C.X.; Kerr, P.G.; Wong, P.K.; Wu, Y.H. Bioremediation of agricultural solid waste leachates with diverse species of Cu (II) and Cd (II) by periphyton. Bioresour. Technol. 2016, 221, 214–221. [Google Scholar] [CrossRef]
- Bradac, P.; Wagner, B.; Kistler, D.; Traber, J.; Behra, R.; Sigg, L. Cadmium speciation and accumulation in periphyton in a small stream with dynamic concentration variations. Environ. Pollut. 2010, 158, 641–648. [Google Scholar] [CrossRef]
- Larned, S.T. A prospectus for periphyton: Recent and future ecological research. J. N. Am. Benthol. Soc. 2010, 29, 182–206. [Google Scholar] [CrossRef] [Green Version]
- Kanavillil, N.; Thorn, M.; Kurissery, S. Characterization of natural biofilms in temperate inland waters. J. Great Lakes Res. 2012, 38, 429–438. [Google Scholar] [CrossRef]
- Wu, Y.H.; Xia, L.Z.; Yu, Z.Q.; Shabbir, S.; Kerr, P.G. In situ bioremediation of surface waters by periphytons. Bioresour. Technol. 2014, 151, 367–372. [Google Scholar] [CrossRef]
- Liu, J.Z.; Wang, F.W.; Wu, W.T.; Wan, J.J.; Yang, J.L.; Xiang, S.L.; Wu, Y.H. Biosorption of high-concentration Cu (II) by periphytic biofilms and the development of a fiber periphyton bioreactor (FPBR). Bioresour. Technol. 2018, 248, 127–134. [Google Scholar] [CrossRef]
- Chonova, T.; Labanowski, J.; Cournoyer, B.; Chardon, C.; Keck, F.; Laurent, E.; Mondamert, L.; Vasselon, V.; Wiest, L.; Bouchez, A. River biofilm community changes related to pharmaceutical loads emitted by a wastewater treatment plant. Environ. Sci. Pollut. R. 2018, 25, 9254–9264. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hua, X.; Zhang, L.; Song, N.; Dong, D.; Guo, Z. Influence of organic carbon fractions of freshwater biofilms on the sorption for phenanthrene and ofloxacin: The important role of aliphatic carbons. Sci. Total Environ. 2019, 685, 818–826. [Google Scholar] [CrossRef]
- Arnon, S.; Packman, A.I.; Peterson, C.G.; Gray, K.A. Effects of overlying velocity on periphyton structure and denitrification. J. Geophys. Res. Biogeo. 2007, G1. [Google Scholar] [CrossRef] [Green Version]
- Battin, T.J.; Besemer, K.; Bengtsson, M.M.; Romani, A.M.; Packmann, A.I. The ecology and biogeochemistry of stream biofilms. Nat. Rev. Microbiol. 2016, 14, 251–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, J.A.; Bunn, A.; McKinstry, C.; Peacock, A.; Miracle, A.L. Investigating freshwater periphyton community response to uranium with phospholipid fatty acid and denaturing gradient gel electrophoresis analyses. J. Environ. Radioactiv. 2008, 99, 730–738. [Google Scholar] [CrossRef]
- De Godos, I.; Gonzalez, C.; Becares, E.; Garcia-Encina, P.A.; Munoz, R. Simultaneous nutrients and carbon removal during pretreated swine slurry degradation in a tubular biofilm photobioreactor. Appl. Microbiol. Biotechnol. 2009, 82, 187–194. [Google Scholar] [CrossRef]
- Liu, J.Z.; Wu, Y.H.; Wu, C.X.; Muylaert, K.; Vyverman, W.; Yu, H.Q.; Munoz, R.; Rittmann, B. Advanced nutrient removal from surface water by a consortium of attached microalgae and bacteria: A review. Bioresour. Technol. 2017, 241, 1127–1137. [Google Scholar] [CrossRef]
- Shabbir, S.; Faheem, M.; Ali, N.; Kerr, P.G.; Wu, Y.H. Evaluating role of immobilized periphyton in bioremediation of azo dye amaranth. Bioresour. Technol. 2017, 225, 395–401. [Google Scholar] [CrossRef]
- Tang, J.; Wu, Y.H.; Esquivel-Elizondo, S.; Sorensen, S.J.; Rittmann, B.E. How microbial aggregates protect against nanoparticle Toxicity. Trends Biotechnol. 2018, 36, 1171–1182. [Google Scholar] [CrossRef]
- Meylan, S.; Behra, R.; Sigg, L. Accumulation of copper and zinc in periphyton in response to dynamic variations of metal speciation in freshwater. Environ. Sci. Technol. 2003, 37, 5204–5212. [Google Scholar] [CrossRef]
- Wu, Y.H.; Zhang, S.Q.; Zhao, H.J.; Yang, L.Z. Environmentally benign periphyton bioreactors for controlling cyanobacterial growth. Bioresour. Technol. 2010, 101, 9681–9687. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Dong, D.; Zhang, L.; Song, Z.; Hua, X.; Guo, Z. Response of freshwater biofilms to antibiotic florfenicol and ofloxacin stress: Role of extracellular polymeric substances. Int. J. Environ. Res. Public Health 2019, 16, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, L.K.; Bergey, E.A. Metal toxicity and recovery response of riverine periphytic algae. Sci. Total Environ. 2018, 642, 1020–1031. [Google Scholar] [CrossRef]
- Sousa, M.L.; Chow, F.; Pompeo, M.L.M. Community-level changes in periphytic biofilm caused by copper contamination. J. Appl. Phycol. 2019, 31, 2401–2410. [Google Scholar] [CrossRef]
- Ma, L.; Wang, F.W.; Yu, Y.C.; Liu, J.Z.; Wu, Y.H. Cu removal and response mechanisms of periphytic biofilms in a tubular bioreactor. Bioresour. Technol. 2018, 248, 61–67. [Google Scholar] [CrossRef]
- Liu, J.Z.; Tang, J.; Wan, J.J.; Wu, C.X.; Graham, B.; Kerr, P.G.; Wu, Y.H. Functional sustainability of periphytic biofilms in organic matter and Cu2+ removal during prolonged exposure to TiO2 nanoparticles. J. Hazard. Mater. 2019, 370, 4–12. [Google Scholar] [CrossRef]
- Hou, J.; Li, T.F.; Miao, L.Z.; You, G.X.; Xu, Y.; Liu, S.Q. Effects of titanium dioxide nanoparticles on algal and bacterial communities in periphytic biofilms. Environ. Pollut. 2019, 251, 407–414. [Google Scholar] [CrossRef]
- Honeywill, C.; Paterson, D.N.; Hagerthey, S.E. Determination of microphytobenthic biomass using pulse-amplitude modulated minimum fluorescence. Eur. J. Phycol. 2002, 37, 485–492. [Google Scholar] [CrossRef]
- Ge, Z.W.; Du, H.J.; Gao, Y.L.; Qiu, W.F. Analysis on metabolic functions of stored rice microbial communities by BIOLOG ECO microplates. Front. Microbiol. 2018, 9, 1375. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lyu, T.; Zhang, Y.; Button, M.; Arias, C.A.; Weber, K.P.; Brix, H.; Carvalho, P.N. Impacts of design configuration and plants on the functionality of the microbial community of mesocosm-scale constructed wetlands treating ibuprofen. Water Res. 2018, 131, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Li, G.Z.; Park, S.; Rittmann, B.E. Developing an efficient TiO2-coated biofilm carrier for intimate coupling of photocatalysis and biodegradation. Water Res. 2012, 46, 6489–6496. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.L.; Sun, P.F.; Yang, J.L.; Huang, Y.P.; Wu, Y.H. Kinetics simulation of Cu and Cd removal and the microbial community adaptation in a periphytic biofilm reactor. Bioresour. Technol. 2019, 276, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Xu, Y.; Wu, Y.H.; Tang, J.; Esquivel-Elizondo, S.; Kerr, P.G.; Staddon, P.L.; Liu, J.Z. Functional sustainability of nutrient accumulation by periphytic biofilm under temperature fluctuations. Environ. Technol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhu, N.Y.; Zhu, Y.; Zamir, S.M.; Wu, Y.H. Sustainable pollutant removal by periphytic biofilm via microbial composition shifts induced by uneven distribution of CeO2 nanoparticles. Bioresour. Technol. 2018, 248, 75–81. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, W.; Zhao, W.; Song, J. Responses of Periphyton Microbial Growth, Activity, and Pollutant Removal Efficiency to Cu Exposure. Int. J. Environ. Res. Public Health 2020, 17, 941. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17030941
Zhong W, Zhao W, Song J. Responses of Periphyton Microbial Growth, Activity, and Pollutant Removal Efficiency to Cu Exposure. International Journal of Environmental Research and Public Health. 2020; 17(3):941. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17030941
Chicago/Turabian StyleZhong, Wei, Weiqun Zhao, and Jianhui Song. 2020. "Responses of Periphyton Microbial Growth, Activity, and Pollutant Removal Efficiency to Cu Exposure" International Journal of Environmental Research and Public Health 17, no. 3: 941. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17030941