Effects of Long-Term Endurance Exercise and Lithium Treatment on Neuroprotective Factors in Hippocampus of Obese Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Subject and Method
2.2. Lithium Treatment
2.3. Exercise Protocol
2.4. Tissue and Blood Sampling
2.5. Analysis
2.5.1. H&E Staining
2.5.2. Blood Factor Analysis
2.5.3. Western Blotting
2.6. Statistical Analysis
3. Results
3.1. Body Composition
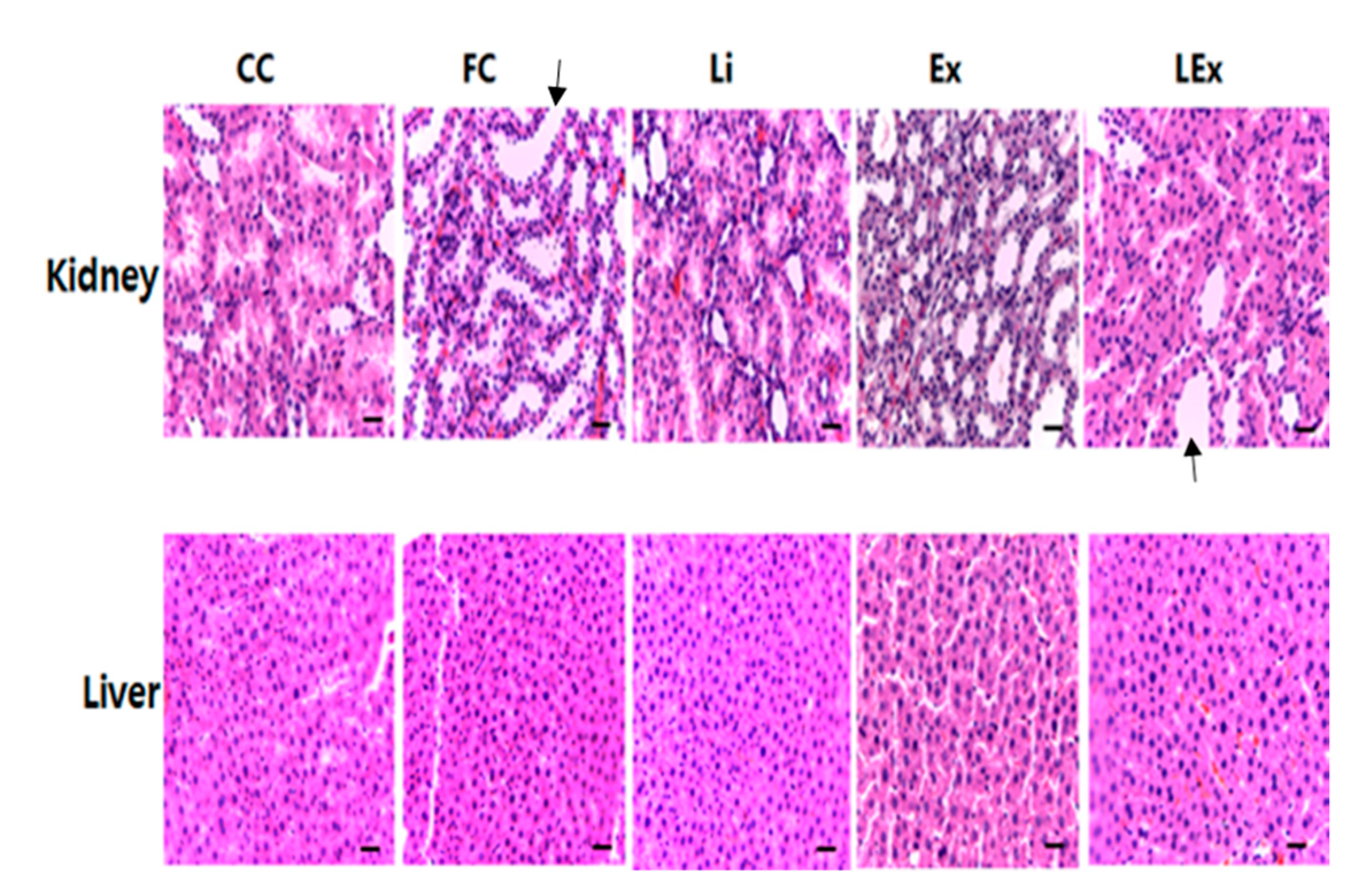
3.2. Toxicity Test
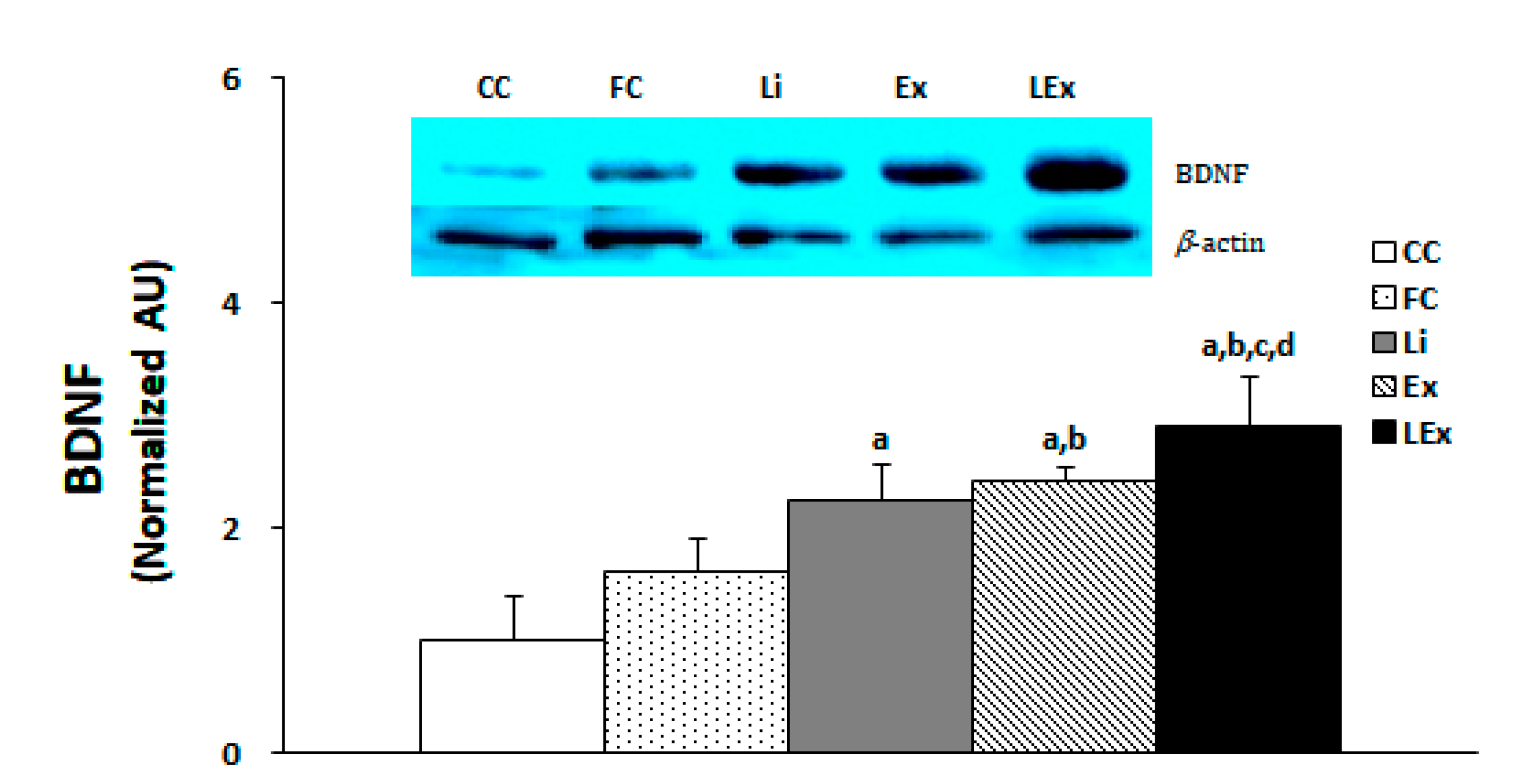
3.3. BDNF Production
3.4. GSK3β/Phospho-GSK3β Expression Ratio
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanson, C.D.; Clarke, C. Is expressed emotion related to estimates of abilitymade by older people with cognitive impairments and their partners? Aging Ment. Health 2013, 17, 535–543. [Google Scholar] [CrossRef]
- Alves, G.; Forsaa, E.B.; Pedersen, K.F.; Dreetz, G.M.; Larsen, J.P. Epidemiology of Parkinson’s disease. J. Neurol. 2008, 255, 18–32. [Google Scholar] [CrossRef]
- Wang, G.D.; Lai, D.J.; Burau, K.D.; Du, X.L. Potential gains in life expectancy from reducing heart disease, cancer, Alzheimer’s disease, kidney disease or HIV/AIDS as major causes of death in the USA. Public Health 2013, 127, 348–356. [Google Scholar] [CrossRef]
- Abbott, R.D.; Ross, G.W.; White, L.R.; Nelson, J.S.; Masaki, K.H.; Tanner, C.M.; Curb, J.D.; Blanchette, P.L.; Popper, J.S.; Petrovitch, H. Midlife adiposity and the future risk of Parkinson’s disease. Neurology 2002, 59, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Hassing, L.B.; Dahl, A.K.; Thorvaldsson, V.; Berg, S.; Gatz, M.; Pedersen, N.L.; Johansson, B. Overweight in midlife and risk of dementia: A 40-year follow-up study. Int. J. Obes. 2009, 33, 893–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivipelto, M.; Ngandu, T.; Fratiglioni, L.; Viitanen, M.; Kåreholt, I.; Winblad, B.; Helkala, E.L.; Tuomilehto, J.; Soininen, H.; Nissinen, A. Obesity and vascular risk factors at midlife and the risk of dementia and Alzheimer disease. Arch. Neurol. 2005, 62, 1556–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlskog, J.E. Cheaper, simpler, and better: Tips for treating seniors with Parkinson disease. Mayo Clin. Proc. 2011, 86, 1211–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arida, R.M.; Scorza, F.A.; Scorza, C.A.; Cavalheiro, E.A. Is physical activity beneficial for recovery in temporal lobe epilepsy? Evidences from animal studies. Neurosci. Biobehav. Rev. 2009, 33, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Buchman, A.S.; Boyle, P.A.; Yu, L.; Shah, R.C.; Wilson, R.S.; Bennett, D.A. Total daily physical activity and the risk of AD and cognitive decline in older adults. Neurology 2012, 78, 1323–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Long, W.; Liu, G.; Zhang, X.; Yang, X. Effect of seabuckthorn (Hippophae rhamnoides ssp. sinensis) leaf extract on the swimming endurance and exhaustive exercise-induced oxidative stress of rats. J. Sci. Food Agric. 2012, 92, 736–742. [Google Scholar] [CrossRef]
- Greenberg, M.E.; Xu, B.; Lu, B.; Hempstead, B.L. New insights in the biology of BDNF synthesis and release: Implications in CNS function. J. Neurosci. 2009, 29, 12764–12767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, S.D.; Bramham, C.R. Brain-derived neurotrophic factor mechanisms and function in adult synaptic plasticity: New insights and implications for therapy. Curr. Opin. Drug Discov. Devel. 2006, 9, 580–586. [Google Scholar] [PubMed]
- Hariri, A.R.; Goldberg, T.E.; Mattay, V.S.; Kolachana, B.S.; Callicott, J.H.; Egan, M.F.; Weinberger, D.R. Brain-derived neurotrophic factor val66met polymorphism affects human memory-related hippocampal activity and predicts memory performance. J. Neurosci. 2003, 23, 6690–6694. [Google Scholar] [CrossRef] [Green Version]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Ferris, L.T.; Williams, J.S.; Shen, C.L. The effect of acute exercise on serum brain-derived neurotrophic factor levels and cognitive function. Med. Sci. Sports Exerc. 2007, 39, 728–734. [Google Scholar] [CrossRef]
- Griffin, É.W.; Mullally, S.; Foley, C.; Warmington, S.A.; O’Mara, S.M.; Kelly, A.M. Aerobic exercise improves hippocampal function and increases BDNF in the serum of young adult males. Physiol. Behav. 2011, 104, 934–941. [Google Scholar] [CrossRef]
- Skriver, K.; Roig, M.; Lundbye-Jensen, J.; Pingel, J.; Helge, J.W.; Kiens, B.; Nielsen, J.B. Acute exercise improves motor memory: Exploring potential biomarkers. Neurobiol. Learn Mem. 2014, 116, 46–58. [Google Scholar] [CrossRef]
- Tsai, C.L.; Chen, F.C.; Pan, C.Y.; Wang, C.H.; Huang, T.H.; Chen, T.C. Impact of acute aerobic exercise and cardiorespiratory fitness on visuospatial attention performance and serum BDNF levels. Psychoneuroendocrinology 2014, 41, 121–131. [Google Scholar] [CrossRef]
- Boyko, M.; Nassar, A.; Kaplanski, J.; Zlotnik, A.; Sharon-Granit, Y.; Azab, A.N. Effects of acute lithium treatment on brain levels of inflammatory mediators in poststroke rats. Biomed. Res. Int. 2015, 2015, 916234. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, D.; Singh, A.; Singh, B.; Bandhu, H.K.; Chand, B.; Singh, N. Effect of lithium augmentation on the trace elemental profile in diabetic rats. Biometals 1999, 12, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Frey, B.N.; Andreazza, A.C.; Rosa, A.R.; Martins, M.R.; Valvassori, S.S.; Réus, G.Z.; Hatch, J.P.; Quevedo, J.; Kapczinski, F. Lithium increases nerve growth factor levels in the rat hippocampus in an animal model of mania. Behav. Pharmacol. 2006, 17, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Hajek, T.; Weiner, M.W. Neuroprotective effects of lithium in human brain? food for thought. Curr. Alzheimer Res. 2016, 13, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Khairova, R.; Pawar, R.; Salvadore, G.; Juruena, M.F.; de Sousa, R.T.; Soeiro-de-Souza, M.G.; Salvador, M.; Zarate, C.A.; Gattaz, W.F.; Machado-Vieira, R. Effects of lithium on oxidative stress parameters in healthy subjects. Mol. Med. Rep. 2012, 5, 680–682. [Google Scholar]
- Machado-Vieira, R.; Manji, H.K.; Zarate, C.A., Jr. The role of lithium in the treatment of bipolar disorder: Convergent evidence for neurotrophic effects as a unifying hypothesis. Bipolar Disord. 2009, 11, 92–109. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, H.R.; Dehpour, A.R.; Mehr, S.E.; Sharifzadeh, M.; Ghahremani, M.H.; Razmi, A.; Ostad, S.N. Lithium attenuates cannabinoid-induced dependence in the animal model: Involvement of phosphorylated ERK1/2 and GSK-3β signaling pathways. Acta Med. Iran 2014, 52, 656–663. [Google Scholar]
- Rahimi-Balaei, M.; Momeny, M.; Babaeikelishomi, R.; Ejtemaei-Mehr, S.; Tavangar, S.M.; Dehpour, A.R. The modulatory effect of lithium on doxorubicin-induced cardiotoxicity in rat. Eur. J. Pharmacol. 2010, 641, 193–198. [Google Scholar] [CrossRef]
- Lenox, R.H.; Hahn, C.G. Overview of the mechanism of action of lithium in the brain: Fifty-year update. J. Clin. Psychiatry 2000, 61, 5–15. [Google Scholar]
- Phiel, C.J.; Klein, P.S. Molecular targets of lithium action. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 789–813. [Google Scholar] [CrossRef]
- Leeds, P.R.; Yu, F.; Wang, Z.; Chiu, C.T.; Zhang, Y.; Leng, Y.; Linares, G.R.; Chuang, D.M. A new avenue for lithium: Intervention in traumatic brain injury. ACS Chem. Neurosci. 2014, 5, 422–433. [Google Scholar] [CrossRef]
- Hancock, C.R.; Han, D.H.; Chen, M.; Terada, S.; Yasuda, T.; Wright, D.C.; Holloszy, J.O. High-fat diets cause insulin resistance despite an increase in muscle mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 7815–7820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanak, A.S.; Chevillard, L.; El Balkhi, S.; Risède, P.; Peoc’h, K.; Mégarbane, B. Study of blood and brain lithium pharmacokinetics in the rat according to three different modalities of poisoning. Toxicol. Sci. 2015, 143, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, J.M., Jr.; Pritchard, H.D.; Braude, M.C.; D’Aguanno, W. Plasma and brain lithium levels after lithium carbonate and lithium chloride administration by different routes in rats. Proc. Soc. Exp. Biol. Med. 1971, 137, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Koltai, E.; Hart, N.; Taylor, A.W.; Goto, S.; Ngo, J.K.; Davies, K.J.; Radak, Z. Age-associated declines in mitochondrial biogenesis and protein quality control factors are minimized by exercise training. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R127–R134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.Y.; Hong, H.S.; Chen, L.L.; Lin, X.H.; Lin, J.H.; Lin, Z. Effects of exercise of different intensities on the angiogenesis, infarct healing, and function of the left ventricle in postmyocardial infarction rats. Coron. Artery Dis. 2011, 22, 497–506. [Google Scholar] [CrossRef]
- Margolis, B.; Ziberstein, A.; Franks, C.; Felder, S.; Kremer, S.; Ullrich, A.; Rhee, S.G.; Skorecki, K.; Schlessinger, J. Effect of phospholipase C-gamma overexpression on PDGF induced second messengers and mitogenesis. Science 1990, 248, 607–610. [Google Scholar] [CrossRef]
- Lowry, E.C.; Blumber, J.M.; Rhea, R.L.; Ranson, J.P. Serum levels of orally administered penicillin. US Armed Forces Med. J. 1951, 2, 265–270. [Google Scholar]
- Chiu, C.T.; Chuang, D.M. Molecular actions and therapeutic potential of lithium in preclinical and clinical studies of CNS disorders. Pharmacol. Ther. 2010, 128, 281–304. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Klein, P.S. Multiple roles for glycogen synthase kinase-3 as a drug target in Alzheimer’s disease. Curr. Drug Targets. 2006, 7, 1389–1397. [Google Scholar] [CrossRef]
- Jope, R.S.; Yuskaitis, C.J.; Beurel, E. Glycogen synthase kinase-3(GSK3): Inflammation, diseases, and therapeutics. Neurochem. Res. 2007, 32, 577–595. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jope, R.S. Is glycogen synthase kinase-3 a central modulator in mood regulation? Neuropsychopharmacology 2010, 35, 2143–2154. [Google Scholar] [CrossRef] [Green Version]
- Meijer, L.; Flajolet, M.; Greengard, P. Pharmacological inhibitors of glycogen synthase kinase 3. Trends Pharmacol. Sci. 2004, 25, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Michalek, S.M.; Jope, R.S. Innate and adaptive immune responses regulated by glycogen synthase kinase-3 (GSK3). Trends Immunol. 2010, 31, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, M.K.; Chuang, D.M. Lithium neuroprotection: Molecular mechanisms and clinical implications. Expert Rev. Mol. Med. 2004, 6, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Rowe, M.K.; Wiest, C.; Chuang, D.M. GSK-3 is a viable potential target for therapeutic intervention in bipolar disorder. Neurosci. Biobehav. Rev. 2007, 31, 920–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, M.H.; Wendland, J.R.; Chuang, D.M. Lithium inhibits Smad3/4 transactivation via increased CREB activity induced by enhanced PKA and AKT signaling. Mol. Cell Neurosci. 2008, 37, 440–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Q.; Shi, T.; Chuang, D.M.; Qian, Y. Lithium reduces ischemia-induced hippocampal CA1 damage and behavioral deficits in gerbils. Brain Res. 2007, 1184, 270–276. [Google Scholar] [CrossRef]
- Tao, X.; West, A.E.; Chen, W.G.; Corfas, G.; Greenberg, M.E. A calcium-responsive transcription factor, CaRF, that regulates neuronal activity-dependent expression of BDNF. Neuron 2002, 33, 383–395. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, S.; Liang, M.H.; Marinova, Z.; Yahyavi, A.; Chuang, D.M. The mood stabilizers lithium and valproate selectively activate the promoter IV of brain-derived neurotrophic factor in neurons. Mol. Psychiatry 2009, 14, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Chuang, D.M.; Wang, Z.; Chiu, C.T. GSK-3 as a target for lithium-induced neuroprotection against excitotoxicity in neuronal cultures and animal models of ischemic stroke. Front. Mol. Neurosci. 2011, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Gould, T.D.; Chen, G.; Manji, H.K. In vivo evidence in the brain for lithium inhibition of glycogen synthase kinase-3. Neuropsychopharmacology 2004, 29, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Kaidanovich-Beilin, O.; Milman, A.; Weizman, A.; Pick, C.G.; Eldar-Finkelman, H. Rapid antidepressive-like activity of specific glycogen synthase kinase-3 inhibitor and its effect on beta-catenin in mouse hippocampus. Biol. Psychiatry 2004, 55, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Tso, P.; Woods, S.C. Receptor CD36 links a risk-associated allele to obesity and metabolic disorders. J. Biol. Chem. 2018, 293, 13349–13350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, K.I.; Miller, D.L.; Roecklein, K.A. The aging hippocampus: Interactions between exercise, depression, and BDNF. Neuroscientist 2012, 18, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Leem, Y.H.; Kato, M.; Chang, H. Regular exercise and creatine supplementation prevent chronic mild stress-induced decrease in hippocampal neurogenesis via Wnt/GSK3β/β-catenin pathway. J. Exerc. Nutrition Biochem. 2018, 30, 1–6. [Google Scholar] [CrossRef]
- Wang, L.R.; Kim, S.H.; Baek, S.S. Effects of treadmill exercise on the anxiety-like behavior through modulation of GSK3β/β-catenin signaling in the maternal separation rat pup. J. Exerc. Rehabil. 2019, 26, 206–212. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, W.W.; Ma, P.; Ma, Z.X.; Hao, M.; Adelusi, T.I.; Du, L.; Yin, X.X.; Lu, Q. Swimming training alleviated insulin resistance through Wnt3a/β-catenin signaling in type 2 diabetic rats. Iran. J. Basic Med. Sci. 2017, 20, 1220–1226. [Google Scholar]
- Pena, G.S.; Paez, H.G.; Johnson, T.K.; Halle, J.L.; Carzoli, J.P.; Visavadiya, N.P.; Zourdos, M.C.; Whitehurst, M.A.; Khamoui, A.V. Hippocampal growth factor and myokine cathepsin B expression following aerobic and resistance training in 3xTg-AD mice. Int. J. Chronic Dis. 2020, 2020, 5919501. [Google Scholar] [CrossRef] [Green Version]
- Lista, I.; Sorrentino, G. Biological mechanisms of physical activity in preventing cognitive decline. Cell Mol. Neurobiol. 2010, 30, 493–503. [Google Scholar] [CrossRef]
- Tang, S.W.; Chu, E.; Hui, T.; Helmeste, D.; Law, C. Influence of exercise on serum brain-derived neurotrophic factor concentrations in healthy human subjects. Neurosci. Lett. 2008, 431, 62–65. [Google Scholar] [CrossRef]
- Neeper, S.A.; Gómez-Pinilla, F.; Choi, J.; Cotman, C. Exercise and brain neurotrophins. Nature 1995, 373, 109. [Google Scholar] [CrossRef] [PubMed]
- Russo-Neustadt, A.; Beard, R.C.; Cotman, C.W. Exercise, antidepressant medications, and enhanced brain derived neurotrophic factor expression. Neuropsychopharmacology 1999, 21, 679–682. [Google Scholar] [CrossRef] [Green Version]
- Zoladz, J.A.; Pilc, A.; Majerczak, J.; Grandys, M.; Zapart-Bukowska, J.; Duda, K. Endurance training increases plasma brain-derived neurotrophic factor concentration in young healthy men. J. Physiol. Pharmacol. 2008, 59, 119–132. [Google Scholar] [PubMed]
- Adlard, P.A.; Cotman, C.W. Voluntary exercise protects against stress-induced decreases in brain-derived neurotrophic factor protein expression. Neuroscience 2004, 124, 985–992. [Google Scholar] [CrossRef]
- Pizzorusso, T.; Ratto, G.M.; Putignano, E.; Maffei, L. Brain-derived neurotrophic factor causes cAMP response element-binding protein phosphorylation in absence of calcium increases in slices and cultured neurons from rat visual cortex. J. Neurosci. 2000, 20, 2809–2816. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, E.E.; Theibert, A.B. Functions of PI 3-kinase in development of the nervous system. Int. J. Dev. Neurosci. 2002, 20, 187–197. [Google Scholar] [CrossRef]
- Chen, M.J.; Russo-Neustadt, A.A. Exercise activates the phosphatidylinositol 3-kinase pathway. Brain Res. Mol. Brain Res. 2005, 135, 181–193. [Google Scholar] [CrossRef]
- Duman, R.S.; Voleti, B. Signaling pathways underlying the pathophysiology and treatment of depression: Novel mechanisms for rapid-acting agents. Trends Neurosci. 2012, 35, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Graham, L.C.; Grabowska, W.A.; Chun, Y.; Risacher, S.L.; Philip, V.M.; Saykin, A.J.; Alzheimer’s Disease Neuroimaging Initiative (ADNI); Sukoff Rizzo, S.J.; Howell, G.R. Exercise prevents obesity-induced cognitive decline and white matter damage in mice. Neurobiol. Aging 2019, 80, 154–172. [Google Scholar] [CrossRef]
- Park, H.S.; Park, S.S.; Kim, C.J.; Shin, M.S.; Kim, T.W. Exercise Alleviates Cognitive Functions by Enhancing Hippocampal Insulin Signaling and Neuroplasticity in High-Fat Diet-Induced Obesity. Nutrients 2019, 11, 1603. [Google Scholar] [CrossRef] [Green Version]
- Bichet, D.G. Lithium, cyclic AMP signaling, A-kinase anchoring proteins, and aquaporin-2. J. Am. Soc. Nephrol. 2006, 7, 920–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robben, J.H.; Knoers, N.V.; Deen, P.M. Cell biological aspects of the vasopressin type-2 receptor and aquaporin 2 water channel in nephrogenic diabetes insipidus. Am. J. Physiol. Renal Physiol. 2006, 291, F257–F270. [Google Scholar] [CrossRef] [PubMed]
- Trepiccione, F.; Christensen, B.M. Lithium-induced nephrogenic diabetes insipidus: New clinical and experimental findings. J. Nephrol. 2010, 23, S43–S48. [Google Scholar] [PubMed]
- Presne, C.; Fakhouri, F.; Noël, L.H.; Stengel, B.; Even, C.; Kreis, H.; Mignon, F.; Grünfeld, J.P. Lithium-induced nephropathy: Rate of progression and prognostic factors. Kidney Int. 2003, 64, 585–592. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Group | 1 Week | 3 Week | 6 Week | 9 Week | 12 Week |
|---|---|---|---|---|---|
| CC | 39.9 ± 4.5 | 48.7 ± 1.8 | 52.4 ± 1.8 | 56.9 ± 3.8 | 57.5 ± 4.2 |
| FC | 53.0 ± 3.3 a | 64.0 ± 0.8 a | 68.5 ± 2.3 a | 73.0 ± 0.8a | 72.0 ± 0.9 a |
| Li | 50.1 ± 2.6 a | 63.0 ± 1.9 a | 65.3 ± 4.6 a | 82.4 ± 2.1 a | 77.3 ± 0.5 a |
| Ex | 49.3 ± 4.4 a | 57.3 ± 0.9 a | 69.8 ± 2.3 a | 67.3 ± 2.4 a | 70.9 ± 1.3 a |
| Lex | 50.0 ± 4.4a | 67.2 ± 4.3a | 76.0 ± 1.9 a | 75.9 ± 0.2 a | 72.9 ± 0.3 a |
| Group | 0 Week | 8 Week | 12 Week |
|---|---|---|---|
| CC | 301.0 ± 3.0 | 403.0 ± 13.0 | 462.0 ± 9.5 |
| FC | 300.0 ± 2.7 | 500.4 ± 1.2 a | 590.2 ± 20.0 a |
| Li | 299.8 ± 2.0 | 503.8 ± 1.5 a | 536.5 ± 8.4 a,b |
| Ex | 302.1 ± 1.5 | 505.0 ± 6.4 a | 555.8 ± 16.0 a,b |
| Lex | 300.8 ± 0.9 | 502.0 ± 3.5 a | 527.3 ± 24.2 a,b |
| Group | Retroperitoneal | Epididymal | Mesenteric | Visceral |
|---|---|---|---|---|
| CC | 16.39 ± 1.63 | 15.88 ± 0.14 | 10.97 ± 1.12 | 43.24 ± 5.02 |
| FC | 28.46 ± 0.72 a | 23.12 ± 0.66 a | 18.64 ± 1.12 a | 70.23 ± 1.02 a |
| Li | 23.00 ± 2.00 a,b | 20.15 ± 1.63 a | 14.65 ± 0.97 a,b | 56.04 ± 3.01 a,b |
| Ex | 22.05 ± 2.61 a,b | 15.42 ± 0.86 b,c | 14.13 ± 1.23 a,b | 51.61 ± 4.35 a,b |
| Lex | 24.75 ± 1.50 a,b | 16.64 ± 0.80 b | 15.89 ± 1.33 a | 59.49 ± 4.10 a,b |
| CC | FC | Li | Ex | Lex | |
|---|---|---|---|---|---|
| ALT | 2.58 ± 3.0 | 2.53 ± 0.43 | 2.11 ± 0.21 | 2.92 ± 0.42 | 2.15 ± 0.39 |
| AST | 0.35 ± 0.02 | 0.39 ± 0.02 | 0.36 ± 0.01 | 0.37 ± 0.02 | 0.35 ± 0.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Cheon, W.; Kim, K. Effects of Long-Term Endurance Exercise and Lithium Treatment on Neuroprotective Factors in Hippocampus of Obese Rats. Int. J. Environ. Res. Public Health 2020, 17, 3317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17093317
Park J, Cheon W, Kim K. Effects of Long-Term Endurance Exercise and Lithium Treatment on Neuroprotective Factors in Hippocampus of Obese Rats. International Journal of Environmental Research and Public Health. 2020; 17(9):3317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17093317
Chicago/Turabian StylePark, Jusik, Wookwang Cheon, and Kijin Kim. 2020. "Effects of Long-Term Endurance Exercise and Lithium Treatment on Neuroprotective Factors in Hippocampus of Obese Rats" International Journal of Environmental Research and Public Health 17, no. 9: 3317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17093317