Implicit Motor Imagery and the Lateral Occipitotemporal Cortex: Hints for Tailoring Non-Invasive Brain Stimulation

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
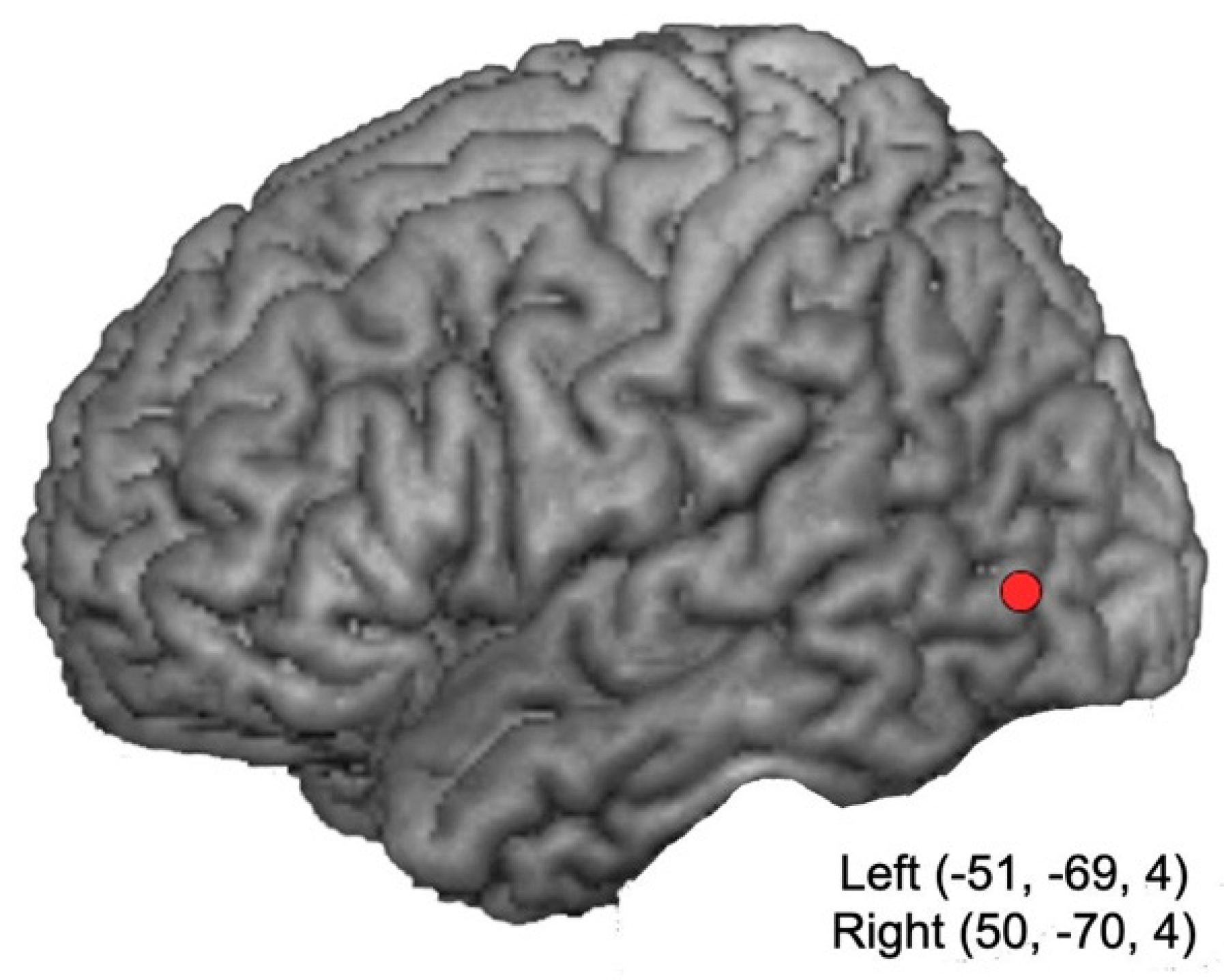
2.2. Neuronavigation and TMS
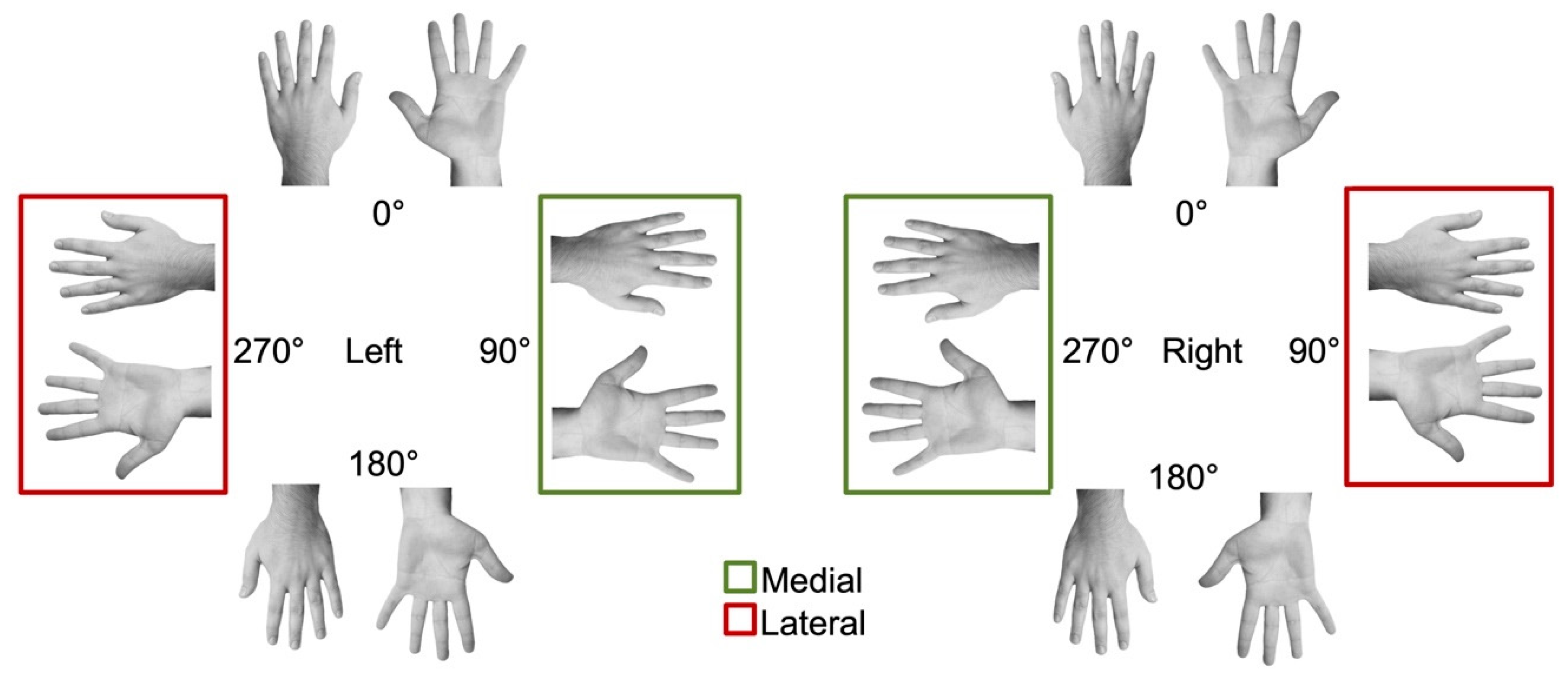
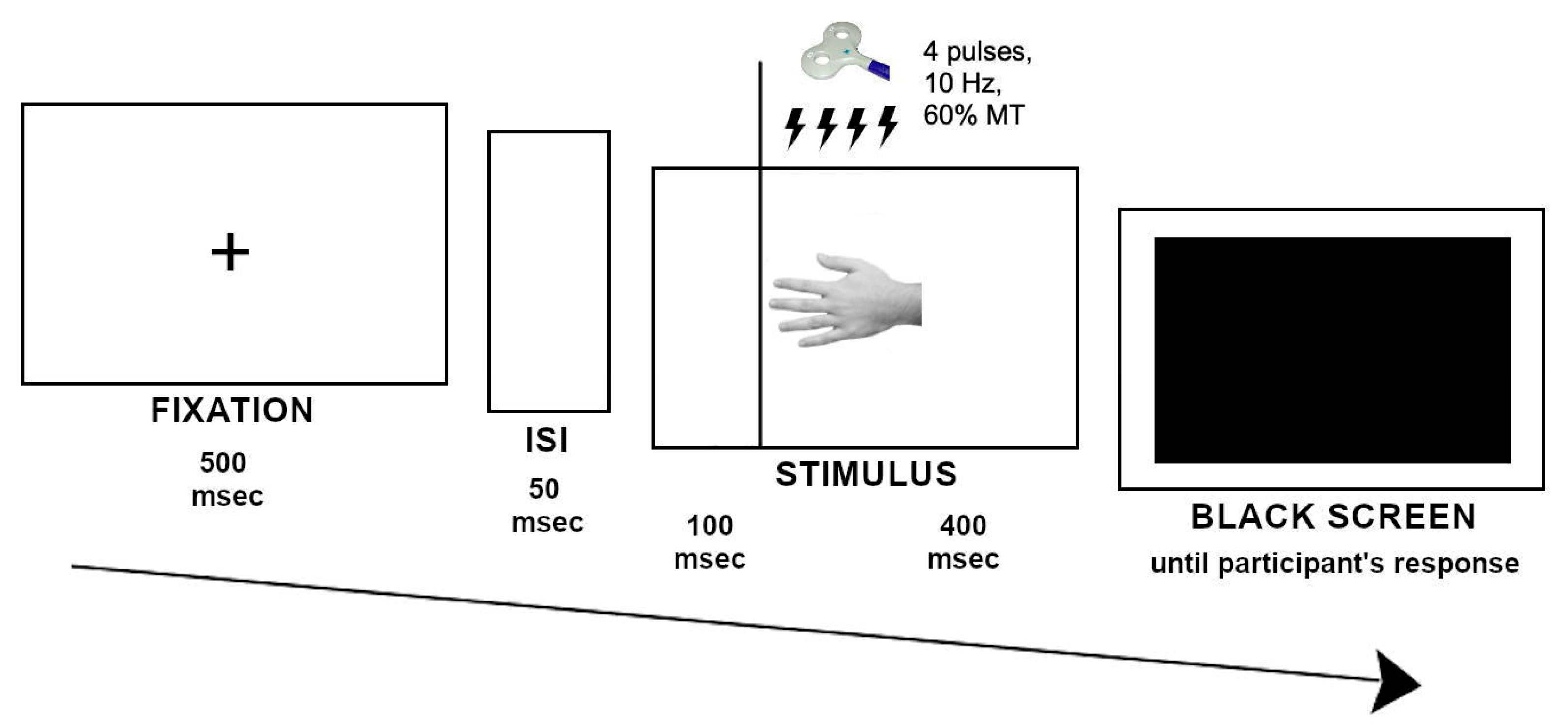
2.3. Experimental Task and Procedure
2.4. Statistical Analysis
3. Results
3.1. Hand Laterality and Spatial Orientation
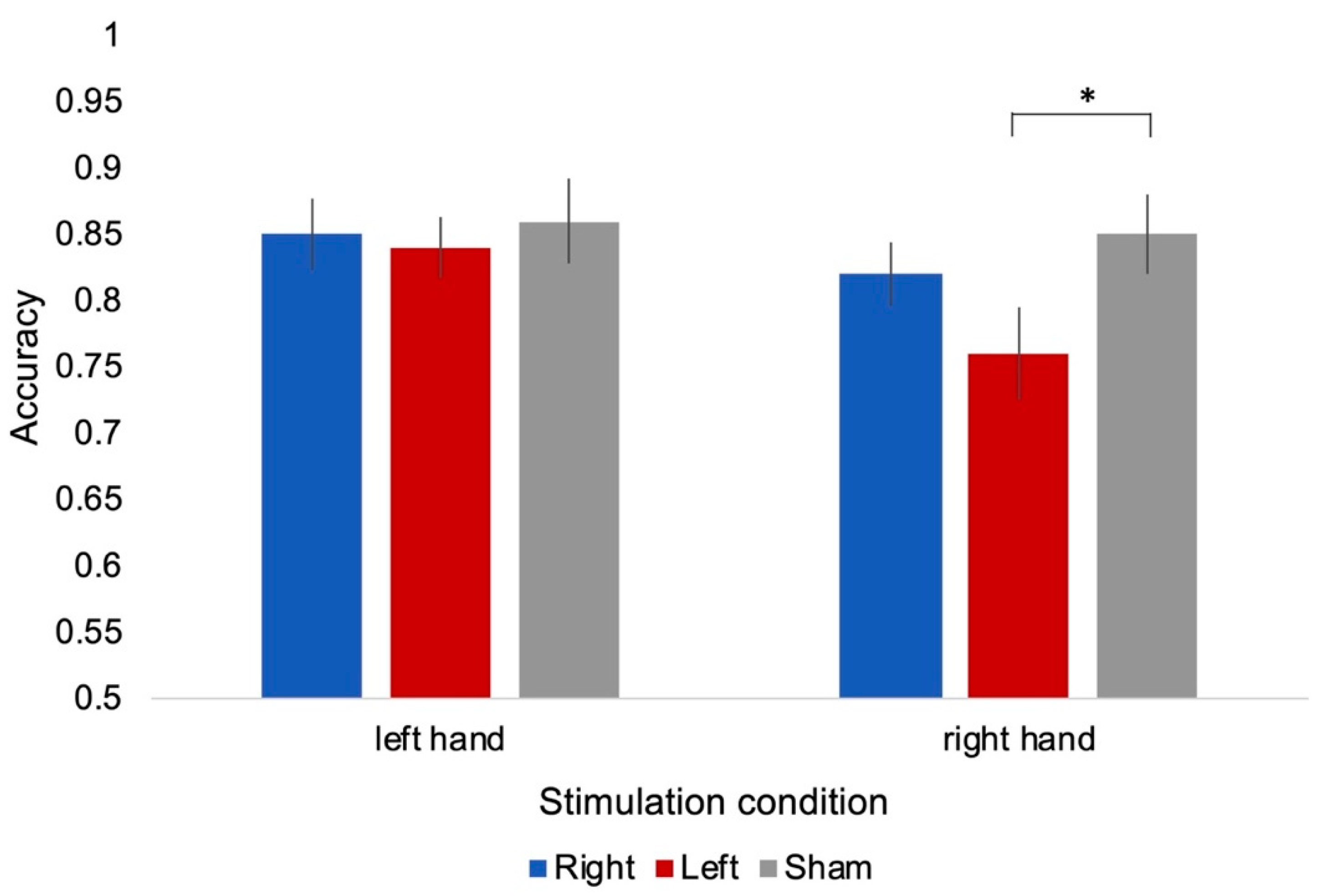
3.1.1. Accuracy
3.1.2. RTs
3.2. Biomechanical Effect
3.2.1. Accuracy
3.2.2. RTs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Conson, M.; Pistoia, F.; Sarà, M.; Grossi, D.; Trojano, L. Recognition and mental manipulation of body parts dissociate in locked-in syndrome. Brain Cogn. 2010, 73, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Conson, M.; Sacco, S.; Sarà, M.; Pistoia, F.; Grossi, D.; Trojano, L. Selective motor imagery defect in patients with locked- in syndrome. Neuropsychologia 2008, 46, 2622–2628. [Google Scholar] [CrossRef] [PubMed]
- Fiori, F.; Sedda, A.; Ferrè, E.R.; Toraldo, A.; Querzola, M.; Pasotti, F.; Ovadia, D.; Piroddi, C.; Dell’Aquila, R.; Lunetta, C.; et al. Exploring motor and visual imagery in amyotrophic lateral sclerosis. Exp. Brain Res. 2013, 226, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Fiori, F.; Sedda, A.; Ferrè, E.R.; Toraldo, A.; Querzola, M.; Pasotti, F.; Ovadia, D.; Piroddi, C.; Dell’Aquila, R.; Redaelli, T.; et al. Motor imagery in spinal cord injury patients: Moving makes the difference. J. Neuropsychol. 2014, 8, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Amick, M.M.; Schendan, H.E.; Ganis, G.; Cronin-Golomb, A. Frontostriatal circuits are necessary for visuomotor transformation: Mental rotation in Parkinson’s disease. Neuropsychologia 2006, 44, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Dominey, P.; Decety, J.; Broussolle, E.; Chazot, G.; Jeannerod, M. Motor imagery of a lateralized sequential task is asymmetrically slowed in hemi-Parkinson’s patients. Neuropsychologia 1995, 33, 727–741. [Google Scholar] [CrossRef]
- Helmich, R.C.; de Lange, F.P.; Bloem, B.R.; Toni, I. Cerebral compensation during motor imagery in Parkinson’s disease. Neuropsychologia 2007, 45, 2201–2215. [Google Scholar] [CrossRef]
- Van Nuenen, B.F.; Helmich, R.C.; Buenen, N.; van de Warrenburg, B.P.; Bloem, B.R.; Toni, I. Compensatory activity in the extrastriate body area of Parkinson’s disease patients. J. Neurosci. 2012, 32, 9546–9553. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, M.; Takahashi, T.; Hirosawa, T.; Oboshi, Y.; Yoshikawa, E.; Minabe, Y.; Ouchi, Y. The Lateral occipito-temporal cortex is involved in the mental manipulation of body part imagery. Front. Hum. Neurosci. 2017, 11, 181. [Google Scholar] [CrossRef] [Green Version]
- Paizis, C.; Skoura, X.; Personnier, P.; Papaxanthis, C. Motor Asymmetry Attenuation in Older Adults during Imagined Arm Movements. Front. Aging Neurosci. 2014, 6, 49. [Google Scholar] [CrossRef]
- Personnier, P.; Ballay, Y.; Papaxanthis, C. Mentally represented motor actions in normal aging: III. Electromyographic features of imagined arm movements. Behav. Brain Res. 2010, 206, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Personnier, P.; Paizis, C.; Ballay, Y.; Papaxanthis, C. Mentally represented motor actions in normal aging II. The influence of the gravito-inertial context on the duration of overt and covert arm movements. Behav. Brain Res. 2008, 186, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Zapparoli, L.; Saetta, G.; De Santis, C.; Gandola, M.; Zerbi, A.; Banfi, G.; Paulesu, E. When I am (almost) 64: The effect of normal ageing on implicit motor imagery in young elderlies. Behav. Brain Res. 2016, 303, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Zapparoli, L.; Invernizzi, P.; Gandola, M.; Verardi, M.; Berlingeri, M.; Sberna, M.; De Santis, A.; Zerbi, A.; Banfi, G.; Bottini, G.; et al. Mental images across the adult lifespan: A behavioural and fMRI investigation of motor execution and motor imagery. Exp. Brain Res. 2013, 224, 519–540. [Google Scholar] [CrossRef]
- Downing, P.E.; Peelen, M.V. Body selectivity in occipitotemporal cortex: Causal evidence. Neuropsychologia 2015, 83, 138–148. [Google Scholar] [CrossRef]
- Downing, P.E.; Jiang, Y.; Shuman, M.; Kanwisher, N. A cortical area selective for visual processing for the human body. Science 2001, 293, 2470–2473. [Google Scholar] [CrossRef]
- Collet, C.; Guillot, A.; Lebon, F.; MacIntyre, T.; Moran, A. Measuring motor imagery using psychometric, behavioral, and psychophysiological tools. Exerc. Sport Sci. Rev. 2011, 39, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Hétu, S.; Grégoire, M.; Saimpont, A.; Coll, M.P.; Eugène, F.; Michon, P.E.; Jackson, P.L. The neural network of motor imagery: An ALE meta-analysis. Neurosci. Biobehav. Rev. 2013, 37, 930–949. [Google Scholar] [CrossRef]
- Parsons, L.M. Temporal and kinematic properties of motor behavior reflected in mentally simulated action. J. Exp. Psychol. Hum. Percept. Perform. 1994, 20, 709–730. [Google Scholar] [CrossRef]
- Sekiyama, K. Kinesthetic aspects of mental representations in the identification of left and right hands. Percept. Psychophys. 1982, 32, 89–95. [Google Scholar] [CrossRef]
- Tomasino, B.; Gremese, M. Effects of stimulus type and strategy on mental rotation network: An activation likelihood estimation meta-analysis. Front. Hum. Neurosci. 2016, 7, 693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brady, N.; Maguinness, C.; Ní Choisdealbha, A. My hand or yours? Markedly different sensitivity to egocentric and allocentric views in the hand laterality task. PLoS ONE 2011, 6, e23316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conson, M.; De Bellis, F.; Baiano, C.; Zappullo, I.; Raimo, G.; Finelli, C.; Ruggiero, I.; Positano, M.; Trojano, L. Sex differences in implicit motor imagery: Evidence from the hand laterality task. Acta Psychol. 2020, 203, 103010. [Google Scholar] [CrossRef] [PubMed]
- Gentilucci, M.; Daprati, E.; Gangitano, M. Implicit visual analysis in handedness recognition. Conscious. Cogn. 1998, 7, 478–493. [Google Scholar] [CrossRef] [PubMed]
- Ní Choisdealbha, Á.; Brady, N.; Maguinness, C. Differing roles for the dominant and non-dominant hands in the hand laterality task. Exp. Brain Res. 2011, 211, 73–85. [Google Scholar] [CrossRef]
- Zapparoli, L.; Invernizzi, P.; Gandola, M.; Berlingeri, M.; De Santis, A.; Zerbi, A.; Banfi, G.; Paulesu, E. Like the back of the (right) hand? A new fMRI look on the hand laterality task. Exp. Brain Res. 2014, 232, 3873–3895. [Google Scholar] [CrossRef]
- Ferri, F.; Frassinetti, F.; Ardizzi, M.; Costantini, M.; Gallese, V. A sensorimotor network for the bodily self. J. Cogn. Neurosci. 2012, 24, 1584–1595. [Google Scholar] [CrossRef]
- Chen, R.; Classen, J.; Gerloff, C.; Celnik, P.; Wassermann, E.M.; Hallett, M.; Cohen, L.G. Depression of motor cortex excitability by low-frequency transcranial magnetic stimulation. Neurology 1997, 48, 1398–1403. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Valls-Sole, J.; Wassermann, E.M.; Hallett, M. Responses to rapid-rate transcranial magnetic stimulation of the human motor cortex. Brain 1994, 117, 847–858. [Google Scholar] [CrossRef]
- Luber, B.; Lisanby, S.H. Enhancement of human cognitive performance using transcranial magnetic stimulation (TMS). NeuroImage 2014, 85, 961–970. [Google Scholar] [CrossRef] [Green Version]
- Urgesi, C.; Candidi, M.; Ionta, S.; Aglioti, S.M. Representation of body identity and body actions in extrastriate body area and ventral premotor cortex. Nat. Neurosci. 2007, 10, 30–31. [Google Scholar] [CrossRef] [Green Version]
- Wassermann, E.M. Risk and safety of repetitive transcranial magnetic stimulation: Report and suggested guidelines from the International Workshop on the Safety of Repetitive Transcranial Magnetic Stimulation, June 5–7, 1996. Electroencephalogr. Clin. Neurophysiol. 1998, 108, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Safety of TMS Consensus Group. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engell, A.D.; McCarthy, G. Selective attention modulates face-specific induced gamma oscillations recorded from ventral occipitotemporal cortex. J. Neurosci. 2010, 30, 8780–8786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thierry, G.; Pegna, A.J.; Dodds, C.; Roberts, M.; Basan, S.; Downing, P. An event-related potential component sensitive to images of the human body. NeuroImage 2006, 32, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Bläsing, B.; Brugger, P.; Weigelt, M.; Schack, T. Does thumb posture influence the mental rotation of hands? Neurosci. Lett. 2013, 534, 139–144. [Google Scholar] [CrossRef]
- Ionta, S.; Fourkas, A.D.; Fiorio, M.; Aglioti, S.M. The influence of hands posture on mental rotation of hands and feet. Exp. Brain Res. 2007, 183, 1–7. [Google Scholar] [CrossRef] [Green Version]
- De Bellis, F.; Trojano, L.; Errico, D.; Grossi, D.; Conson, M. Whose hand is this? Differential responses of right and left extrastriate body areas to visual images of self and others hands. Cogn. Affect. Behav. Neurosci. 2017, 17, 826–837. [Google Scholar] [CrossRef]
- Conson, M.; Aromino, A.R.; Trojano, L. Whose hand is this? Handedness and visual perspective modulate self/other discrimination. Exp. Brain Res. 2010, 206, 449–453. [Google Scholar] [CrossRef]
- Kanwisher, N.; McDermott, J.; Chun, M.M. The fusiform face area: A module in human extrastriate cortex specialized for face perception. J. Neurosci. 1997, 17, 4302–4311. [Google Scholar] [CrossRef]
- Bracci, S.; Ietswaart, M.; Peelen, M.V.; Cavina-Pratesi, C. Dissociable neural responses to hands and non-hand body parts in human left extrastriate visual cortex. J. Neurophysiol. 2010, 103, 3389–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebner, H.R.; Hartwigsen, G.; Kassuba, T.; Rothwell, J.C. How does transcranial magnetic stimulation modify neuronal activity in the brain? Implications for studies of cognition. Cortex 2009, 45, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conson, M.; Errico, D.; Mazzarella, E.; De Bellis, F.; Grossi, D.; Trojano, L. Impact of body posture on laterality judgement and explicit recognition tasks performed on self and others’ hands. Exp. Brain Res. 2015, 233, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Conson, M.; Mazzarella, E.; Trojano, L. Self-touch affects motor imagery: A study on posture interference effect. Exp. Brain Res. 2011, 215, 115–122. [Google Scholar] [CrossRef]
- Limanowski, J.; Blankenburg, F. Integration of visual and proprioceptive limb position information in human posterior parietal, premotor, and extrastriate cortex. J. Neurosci. 2016, 36, 2582–2589. [Google Scholar] [CrossRef] [Green Version]
- Limanowski, J.B.; Blankenburg, F. Network activity underlying the illusory self-attribution of a dummy arm. Hum. Brain Mapp. 2015, 36, 2284–2304. [Google Scholar] [CrossRef]
- Wold, A.; Limanowski, J.; Walter, H.; Blankenburg, F. Proprioceptive drift in the rubber hand illusion is intensified following 1 Hz TMS of the left EBA. Front. Hum. Neurosci. 2014, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Astafiev, S.V.; Stanley, C.M.; Shulman, G.L.; Corbetta, M. Extrastriate body area in human occipital cortex responds to the performance of motor actions. Nat. Neurosci. 2004, 7, 542–548. [Google Scholar] [CrossRef]
- Gentilucci, M.; Daprati, E.; Gangitano, M. Right-handers and left-handers have different representations of their own hand. Cogn. Brain Res. 1998, 6, 185–192. [Google Scholar] [CrossRef]
- Ionta, S.; Blanke, O. Differential influence of hands posture on mental rotation of hands and feet in left and right handers. Exp. Brain Res. 2009, 195, 207–217. [Google Scholar] [CrossRef]
- Machii, K.; Cohen, D.; Ramos-Estebanez, C.; Pascual-Leone, A. Safety of rTMS to non-motor cortical areas in healthy participants and patients. Clin. Neurophysiol. 2006, 117, 455–471. [Google Scholar] [CrossRef] [PubMed]
- Schuster, C.; Butler, J.; Andrews, B.; Kischka, U.; Ettlin, T. Comparison of embedded and added motor imagery training in patients after stroke: Results of a randomised controlled pilot trial. Trials 2012, 13, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heremans, E.; Nieuwboer, A.; Feys, P.; Vercruysse, S.; Vandenberghe, W.; Sharma, N.; Helsen, W.F. External cueing improves motor imagery quality in patients with Parkinson disease. Neurorehab. Neural. Repair 2012, 26, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Heremans, E.; Nieuwboer, A.; Spildooren, J.; De Bondt, S.; D’hooge, A.M.; Helsen, W.; Feys, P. Cued motor imagery in patients with multiple sclerosis. Neuroscience 2012, 206, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; Verhagen, L.; de Lange, F.P.; Toni, I. The Extrastriate Body Area Computes Desired Goal States during Action Planning. eNeuro 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Zapparoli, L.; Seghezzi, S.; Sacheli, L.M.; Verga, C.; Banfi, G.; Paulesu, E. Eyes wide shut: How visual cues affect brain patterns of simulated gait. Hum. Brain Mapp. 2020. [Google Scholar] [CrossRef]
- Callow, N.; Jiang, D.; Roberts, R.; Edwards, M.G. Kinesthetic imagery provides additive benefits to internal visual imagery on slalom task performance. J. Sport Exerc. Psychol. 2017, 39, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Girón, E.C.; McIsaac, T.; Nilsen, D. Effects of kinesthetic versus visual imagery practice on two technical dance movements: A pilot study. J. Danc. Med. Sci. 2012, 16, 36–38. [Google Scholar]
- Guillot, A.; Hoyek, N.; Louis, M.; Collet, C. Understanding the timing of motor imagery: Recent findings and future directions. Int. Rev. Sport Exerc. Psychol. 2012, 5, 3–22. [Google Scholar] [CrossRef]
- Olsson, C.-J.; Nyberg, L. Motor imagery: If you can’t do it, you won’t think it. Scand. J. Med. Sci. Sports 2010, 20, 711–715. [Google Scholar] [CrossRef]
- Paris-Alemany, A.; La Touche, R.; Gadea-Mateos, L.; Cuenca-Martínez, F.; Suso-Martí, L. Familiarity and complexity of a movement influences motor imagery in dancers: A cross-sectional study. Scand. J. Med. Sci. Sports 2019, 29, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Jun, S.C. Performance variation in motor imagery brain-computer interface: A brief review. J. Neurosci. Methods 2015, 243, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Blankertz, B.; Sannelli, C.; Halder, S.; Hammer, E.M.; Kübler, A.; Müller, K.R.; Curio, G.; Dickhaus, T. Neurophysiological predictor of SMR-based BCI performance. Neuroimage 2010, 51, 1303–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dorsum | Palm | |||||||
|---|---|---|---|---|---|---|---|---|
| 0° | 90° | 180° | 270° | 0° | 90° | 180° | 270° | |
| Right hand | ||||||||
| Left rTMS | 0.89 | 0.78 | 0.65 | 0.88 | 0.74 | 0.74 | 0.65 | 0.74 |
| 0.22 | 0.29 | 0.28 | 0.22 | 0.26 | 0.30 | 0.30 | 0.32 | |
| Right rTMS | 0.87 | 0.89 | 0.59 | 0.89 | 0.81 | 0.85 | 0.81 | 0.85 |
| 0.19 | 0.21 | 0.35 | 0.26 | 0.27 | 0.24 | 0.22 | 0.22 | |
| Sham | 0.93 | 0.96 | 0.78 | 0.90 | 0.83 | 0.80 | 0.71 | 0.85 |
| 0.14 | 0.09 | 0.24 | 0.19 | 0.33 | 0.29 | 0.30 | 0.22 | |
| Left hand | ||||||||
| Left rTMS | 0.91 | 0.90 | 0.68 | 0.86 | 0.89 | 0.90 | 0.73 | 0.86 |
| 0.17 | 0.22 | 0.37 | 0.19 | 0.22 | 0.13 | 0.24 | 0.21 | |
| Right rTMS | 0.87 | 0.90 | 0.65 | 0.95 | 0.94 | 0.93 | 0.73 | 0.84 |
| 0.18 | 0.25 | 0.32 | 0.11 | 0.16 | 0.12 | 0.33 | 0.19 | |
| Sham | 0.89 | 0.90 | 0.73 | 0.91 | 0.90 | 0.89 | 0.80 | 0.83 |
| 0.19 | 0.26 | 0.27 | 0.19 | 0.19 | 0.24 | 0.26 | 0.28 | |
| Dorsum | Palm | |||||||
|---|---|---|---|---|---|---|---|---|
| 0° | 90° | 180° | 270° | 0° | 90° | 180° | 270° | |
| Right hand | ||||||||
| Left rTMS | 1040 | 1205 | 1564 | 1099 | 1329 | 1458 | 1408 | 1326 |
| 439 | 455 | 596 | 438 | 385 | 519 | 421 | 477 | |
| Right rTMS | 1006 | 1192 | 1533 | 1097 | 1290 | 1425 | 1488 | 1181 |
| 226 | 413 | 667 | 320 | 313 | 512 | 724 | 435 | |
| Sham | 946 | 1230 | 1498 | 1046 | 1302 | 1348 | 1340 | 1095 |
| 390 | 371 | 595 | 403 | 373 | 396 | 455 | 287 | |
| Left hand | ||||||||
| Left rTMS | 1159 | 1091 | 1504 | 1361 | 1255 | 1156 | 1483 | 1337 |
| 456 | 467 | 439 | 564 | 446 | 380 | 596 | 458 | |
| Right rTMS | 1121 | 1050 | 1553 | 1275 | 1239 | 1214 | 1443 | 1311 |
| 391 | 270 | 578 | 443 | 341 | 233 | 419 | 295 | |
| Sham | 1118 | 1076 | 1464 | 1196 | 1238 | 1181 | 1414 | 1202 |
| 319 | 261 | 483 | 366 | 399 | 368 | 437 | 242 | |
| Dorsum | Palm | |||
|---|---|---|---|---|
| Lateral | Medial | Lateral | Medial | |
| Left rTMS | 0.82 | 0.89 | 0.80 | 0.82 |
| 0.22 | 0.22 | 0.20 | 0.20 | |
| Right rTMS | 0.92 | 0.89 | 0.85 | 0.89 |
| 0.13 | 0.22 | 0.16 | 0.14 | |
| Sham | 0.94 | 0.90 | 0.81 | 0.87 |
| 0.11 | 0.18 | 0.24 | 0.21 | |
| Dorsum | Palm | |||
|---|---|---|---|---|
| Lateral | Medial | Lateral | Medial | |
| Left rTMS | 1283 | 1095 | 1397 | 1241 |
| 474 | 445 | 451 | 385 | |
| Right rTMS | 1233 | 1074 | 1368 | 1198 |
| 403 | 279 | 365 | 288 | |
| Sham | 1213 | 1061 | 1275 | 1138 |
| 325 | 301 | 284 | 295 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conson, M.; Cecere, R.; Baiano, C.; De Bellis, F.; Forgione, G.; Zappullo, I.; Trojano, L. Implicit Motor Imagery and the Lateral Occipitotemporal Cortex: Hints for Tailoring Non-Invasive Brain Stimulation. Int. J. Environ. Res. Public Health 2020, 17, 5851. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17165851
Conson M, Cecere R, Baiano C, De Bellis F, Forgione G, Zappullo I, Trojano L. Implicit Motor Imagery and the Lateral Occipitotemporal Cortex: Hints for Tailoring Non-Invasive Brain Stimulation. International Journal of Environmental Research and Public Health. 2020; 17(16):5851. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17165851
Chicago/Turabian StyleConson, Massimiliano, Roberta Cecere, Chiara Baiano, Francesco De Bellis, Gabriela Forgione, Isa Zappullo, and Luigi Trojano. 2020. "Implicit Motor Imagery and the Lateral Occipitotemporal Cortex: Hints for Tailoring Non-Invasive Brain Stimulation" International Journal of Environmental Research and Public Health 17, no. 16: 5851. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17165851