Fenton Reaction-Induced Oxidative Damage to Membrane Lipids and Protective Effects of 17β-Estradiol in Porcine Ovary and Thyroid Homogenates


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Assay of Lipid Peroxidation
2.4. Measurement of Lipid Peroxidation Products
2.5. Statistical Analyses
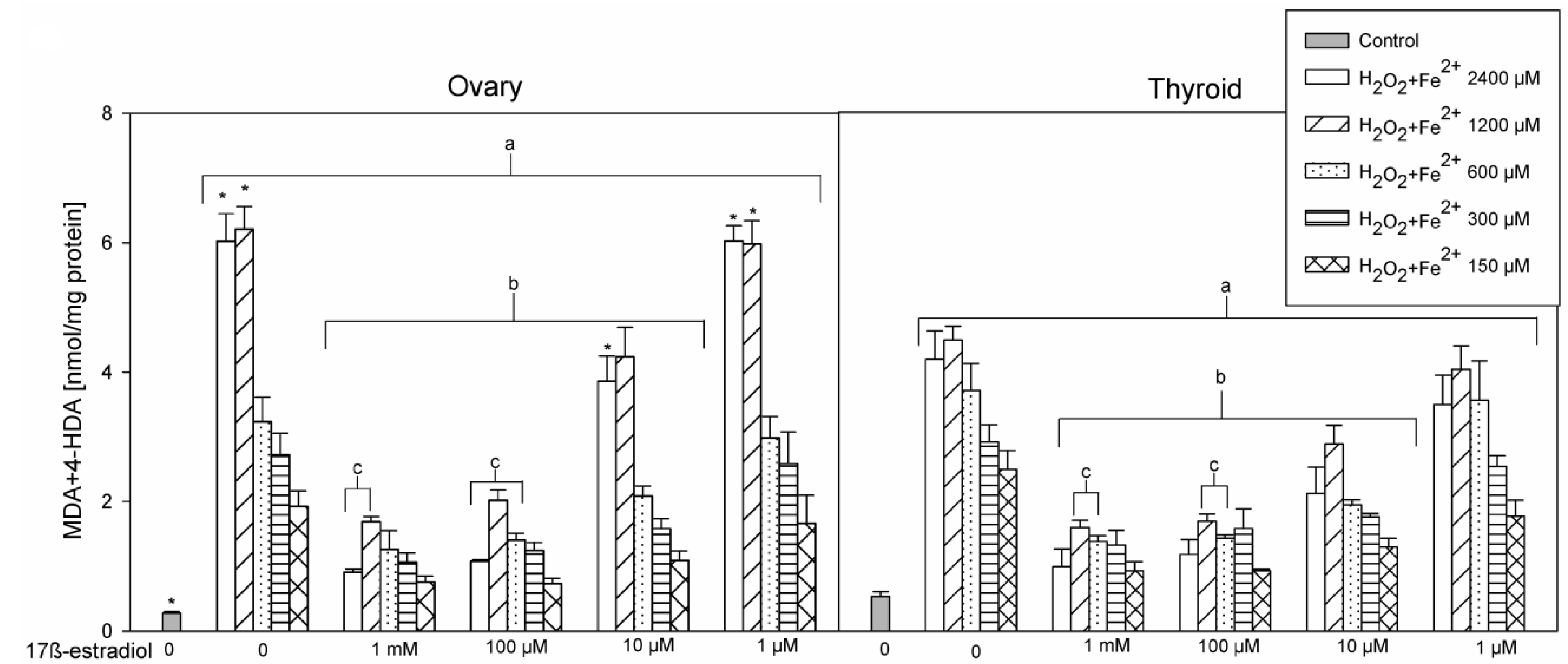
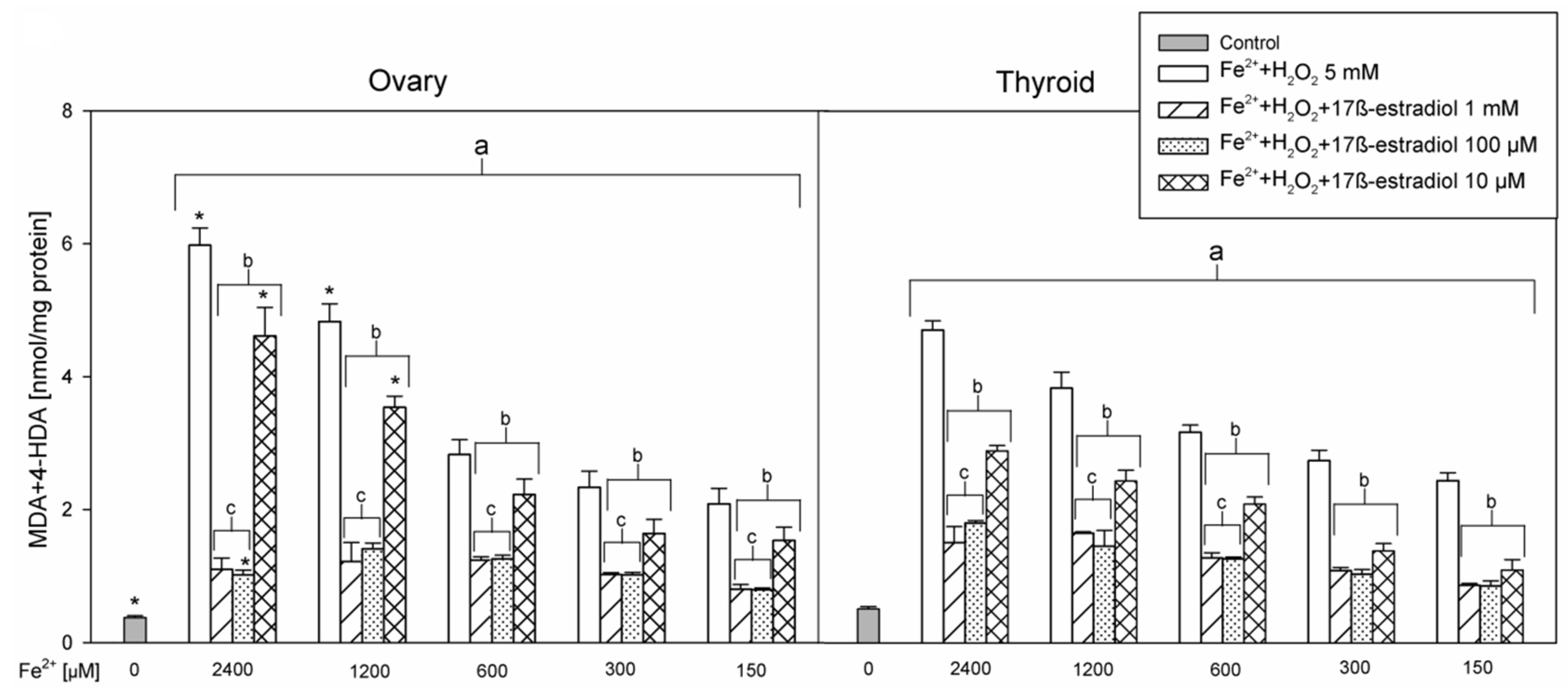
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Halliwell, B. Free radicals and antioxidants: A personal view. Nutr. Rev. 1994, 52, 253–265. [Google Scholar] [CrossRef]
- Koppenol, W.H.; Hider, R.H. Iron and redox cycling. Do’s and don’ts. Free Radic. Biol. Med. 2019, 133, 3–10. [Google Scholar] [CrossRef]
- Fenton, H.J.H. LXXIII. Oxidation of tartaric acid in presence of iron. J. Chem. Soc. Trans. 1894, 65, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, B. Hydroxyl radical and its scavengers in health and disease. Oxid. Med. Cell. Longev. 2011, 809696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, M.; Sugawara, Y.; Wen, K.; Giulivi, C. Generation of oxygen free radicals in thyroid cells and inhibition of thyroid peroxidase. Exp. Biol. Med. 2002, 227, 141–146. [Google Scholar] [CrossRef]
- Chen, M.J.; Chou, C.H.; Shun, C.T.; Mao, T.L.; Wen, W.F.; Chen, C.D.; Chen, S.U.; Yang, Y.S.; Ho, H.N. Iron suppresses ovarian granulosa cell proliferation and arrests cell cycle through regulating p38 mitogen-activated protein kinase/p53/p21 pathway. Biol. Reprod. 2017, 97, 438–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbownik, M.; Lewiński, A. Melatonin reduces Fenton reaction-induced lipid peroxidation in porcine thyroid tissue. J. Cell. Biochem. 2003, 90, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Karbownik-Lewińska, M.; Kokoszko-Bilska, A. Oxidative damage to macromolecules in the thyroid–Experimental evidence. Thyroid Res. 2012, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Karbownik-Lewińska, M.; Stępniak, J.; Krawczyk, J.; Zasada, K.; Szosland, J.; Gesing, A.; Lewiński, A. External hydrogen peroxide is not indispensable for experimental induction of lipid peroxidation via Fenton reaction in porcine ovary homogenates. Neuro Endocrinol. Lett. 2010, 31, 343–347. [Google Scholar]
- Stępniak, J.; Karbownik-Lewińska, M. 17β-estradiol prevents experimentally-induced oxidative damage to membrane lipids and nuclear DNA in porcine ovary. Syst. Biol. Reprod. Med. 2016, 62, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Stępniak, J.; Lewiński, A.; Karbownik-Lewińska, M. Membrane lipids and nuclear DNA are differently susceptive to Fenton reaction substrates in porcine thyroid. Toxicol. In Vitro. 2013, 27, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Stepniak, J.; Lewinski, A.; Karbownik-Lewinska, M. Oxidative damage to membrane lipids in the thyroid -no differences between sexes. Drug Chem. Toxicol. 2019, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Behl, C.; Skutella, T.; Lezoualc’h, F.; Post, A.; Widmann, M.; Newton, C.J.; Holsboer, F. Neuroprotection against Oxidative Stress by Estrogens: Structure-Activity Relationship. Mol. Pharmacol. 1997, 51, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Prokai, L.; Prokai-Tatrai, K.; Perjesi, P.; Zharikova, A.D.; Perez, E.J.; Liu, R.; Simpkins, J.W. Quinol-based cyclic antioxidant mechanism in estrogen neuroprotection. Proc. Natl. Acad. Sci. USA 2003, 100, 11741–11746. [Google Scholar] [CrossRef] [Green Version]
- Stepniak, J.; Lewinski, A.; Karbownik-Lewinska, M. Sexual dimorphism of NADPH oxidase/H₂O₂ system in rat thyroid cells; effect of exogenous 17β-estradiol. Int. J. Mol Sci. 2018, 19, 4063. [Google Scholar] [CrossRef] [Green Version]
- Dahlman-Wright, K.; Cavailles, V.; Fuqua, S.A.; Jordan, V.C.; Katzenellenbogen, J.A.; Korach, K.S.; Maggi, A.; Muramatsu, M.; Parker, M.G.; Gustafsson, J.A. International Union of Pharmacology. LXIV. Estrogen Receptors. Pharmacol. Rev. 2006, 58, 773–781. [Google Scholar] [CrossRef] [Green Version]
- Santin, A.P.; Furlanetto, T.W. Role of Estrogen in Thyroid Function and Growth Regulation. J. Thyroid Res. 2011, 875125. [Google Scholar] [CrossRef] [Green Version]
- Culmsee, C.; Vedder, H.; Ravati, A.; Junker, V.; Otto, D.; Ahlemeyer, B.; Krieg, J.-C.; Krieglstein, J. Neuroprotection by estrogens in a mouse model of focal cerebral ischemia and in cultured neurons: Evidence for a receptor-independent antioxidative mechanism. J. Cereb. Blood Flow Metab. 1999, 19, 1263–1269. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bacon, B.R.; Adams, P.C.; Kowdley, K.V.; Powell, L.W.; Tavill, A.S. Diagnosis and management of hemochromatosis: 2011 Practice Guideline by the American Association for the Study of Liver Diseases. Hepatology 2011, 54, 328–343. [Google Scholar] [CrossRef] [Green Version]
- Bresgen, N.; Eckl, P.M. Oxidative stress and the homeodynamics of iron metabolism. Biomolecules 2015, 5, 808–847. [Google Scholar] [CrossRef] [PubMed]
- Abbassi-Ghanavati, M.; Greer, L.G.; Cunningham, F.G. Pregnancy and laboratory studies: A reference table for clinicians. Obstet. Gynecol. 2009, 114, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Moosmann, B.; Behl, C. The antioxidant neuroprotective effects of estrogens and phenolic compounds are independent from their estrogenic properties. Proc. Natl. Acad. Sci. USA 1999, 96, 8867–8872. [Google Scholar] [CrossRef] [Green Version]
- Vedder, H.; Anthes, N.; Stumm, G.; Würz, C.; Behl, C.; Krieg, J.C. Estrogen hormones reduce lipid peroxidation in cells and tissues of the central nervous system. J. Neurochem. 1999, 72, 2531–2538. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.; Pelle, E.; Huang, X. Iron and menopause: Does increased iron affect the health of postmenopausal women? Antioxidants Redox Signal. 2009, 11, 2939–2943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoya-Estrada, A.; Velázquez-Yescas, K.G.; Veruete-Bedolla, D.B.; Ruiz-Herrera, J.D.; Villarreal-Barranca, A.; Romo-Yañez, J.; Ortiz-Luna, G.F.; Arellano-Eguiluz, A.; Solis-Paredes, M.; Flores-Pliego, A.; et al. Parameters of oxidative stress in reproductive and postmenopausal Mexican women. Int. J. Environ. Res. Public Health 2020, 17, 1492. [Google Scholar] [CrossRef] [Green Version]
- Barton, J.C.; Edwards, C.Q. HFE Hemochromatosis. Synonyms: Hemochromatosis Type 1, HFE-Associated Hemochromatosis, HFE-HH. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK1440/ (accessed on 30 January 2019).
- Edwards, C.Q.; Kelly, T.M.; Ellwein, G.; Kushner, J.P. Thyroid disease in hemochromatosis. Increased incidence in homozygous men. Arch. Intern. Med. 1983, 143, 1890–1893. [Google Scholar] [CrossRef]
- Gannon, P.O.; Medelci, S.; Le Page, C.; Beaulieu, M.; Provencher, D.M.; Mes-Masson, A.M.; Santos, M.M. Impact of hemochromatosis gene (HFE) mutations on epithelial ovarian cancer risk and prognosis. Int. J. Cancer 2011, 128, 2326–2334. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, M.S.; Grace, N.D. Folic acid deficiency and iron overload. Arch. Intern. Med. 1970, 125, 140–144. [Google Scholar] [CrossRef]
- Brinton, L.A.; Trabert, B.; Anderson, G.L.; Falk, R.T.; Felix, A.S.; Fuhrman, B.J.; Gass, M.L.; Kuller, L.H.; Pfeiffer, R.M.; Rohan, T.E.; et al. Serum estrogens and estrogen metabolites and endometrial cancer risk among postmenopausal women. Cancer Epidemiol. Biomarkers Prev. 2016, 25, 1081–1089. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, J.; Yin, C.; Zhao, D.; Zhao, Y.; Li, Y.; Yin, L. Role of β-estradiol in MCF-7 breast cancer cell line based on the bioinformatics analysis. Gynecol. Obstet. Investig. 2018, 84, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, M.; Reiter, R.J.; Burkhardt, S.; Gitto, E.; Tan, D.X.; Lewiński, A. Melatonin attenuates estradiol-induced oxidative damage to DNA: Relevance for cancer prevention. Exp. Biol. Med. 2001, 226, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.R.; Zhang, R.; Lian, Z.X.; Deng, S.L.; Yu, K. Estrogen-receptor expression and function in female reproductive disease. Cells 2019, 8, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rynkowska, A.; Stępniak, J.; Karbownik-Lewińska, M. Fenton Reaction-Induced Oxidative Damage to Membrane Lipids and Protective Effects of 17β-Estradiol in Porcine Ovary and Thyroid Homogenates. Int. J. Environ. Res. Public Health 2020, 17, 6841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17186841
Rynkowska A, Stępniak J, Karbownik-Lewińska M. Fenton Reaction-Induced Oxidative Damage to Membrane Lipids and Protective Effects of 17β-Estradiol in Porcine Ovary and Thyroid Homogenates. International Journal of Environmental Research and Public Health. 2020; 17(18):6841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17186841
Chicago/Turabian StyleRynkowska, Aleksandra, Jan Stępniak, and Małgorzata Karbownik-Lewińska. 2020. "Fenton Reaction-Induced Oxidative Damage to Membrane Lipids and Protective Effects of 17β-Estradiol in Porcine Ovary and Thyroid Homogenates" International Journal of Environmental Research and Public Health 17, no. 18: 6841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17186841