Dietary Thiols: A Potential Supporting Strategy against Oxidative Stress in Heart Failure and Muscular Damage during Sports Activity

 , , ,
, , ,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
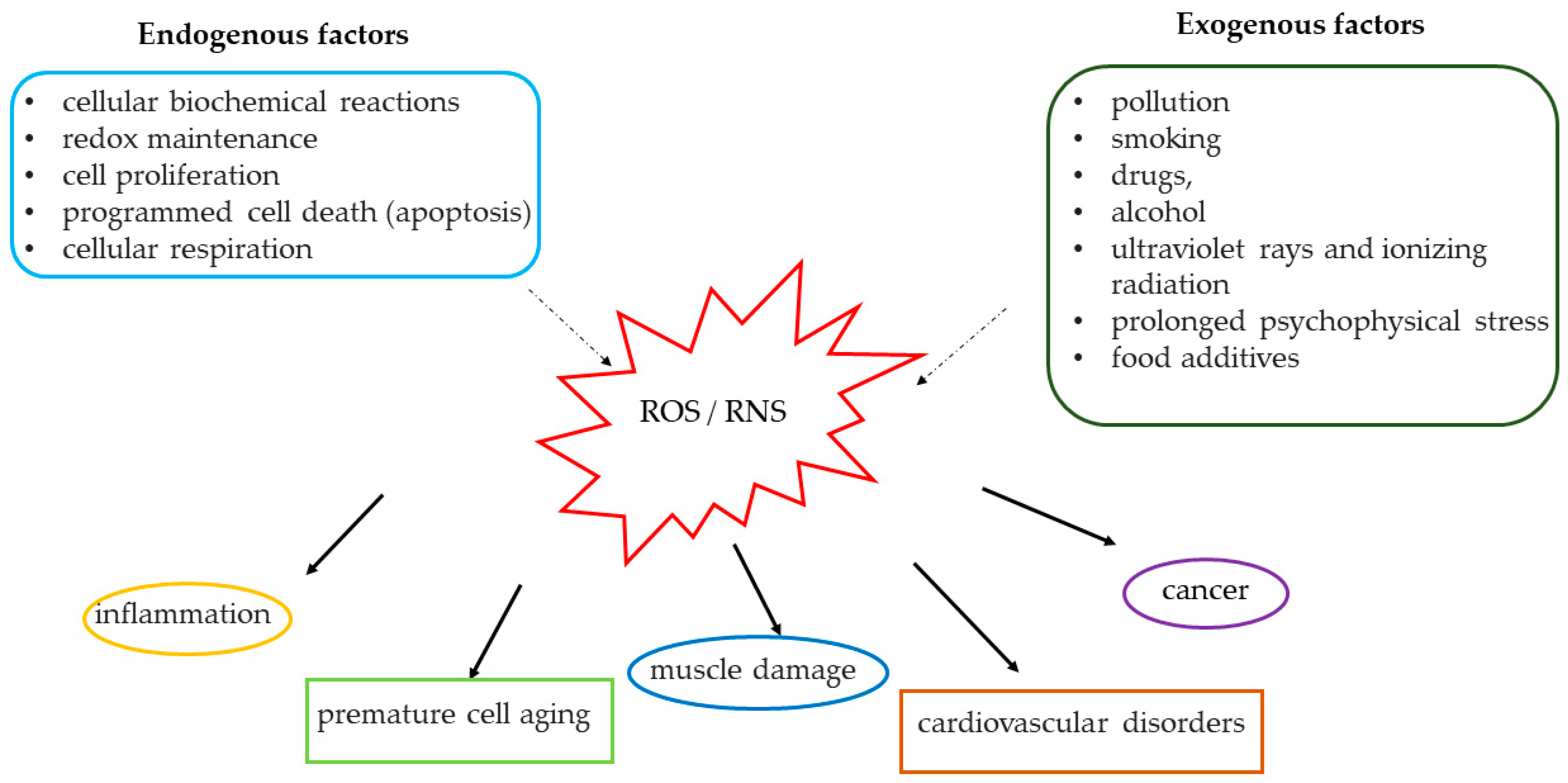
2. Mechanisms of Oxidation
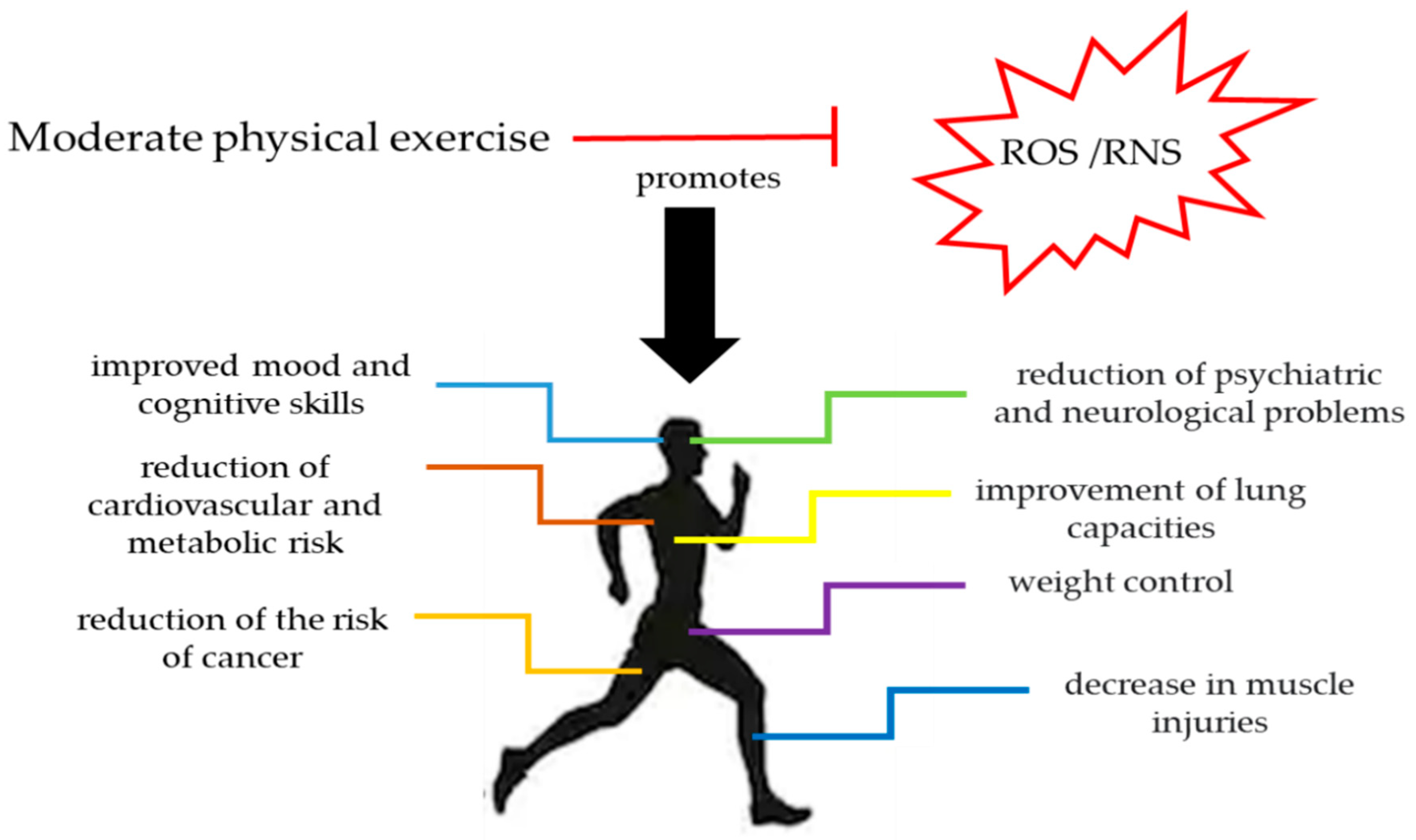
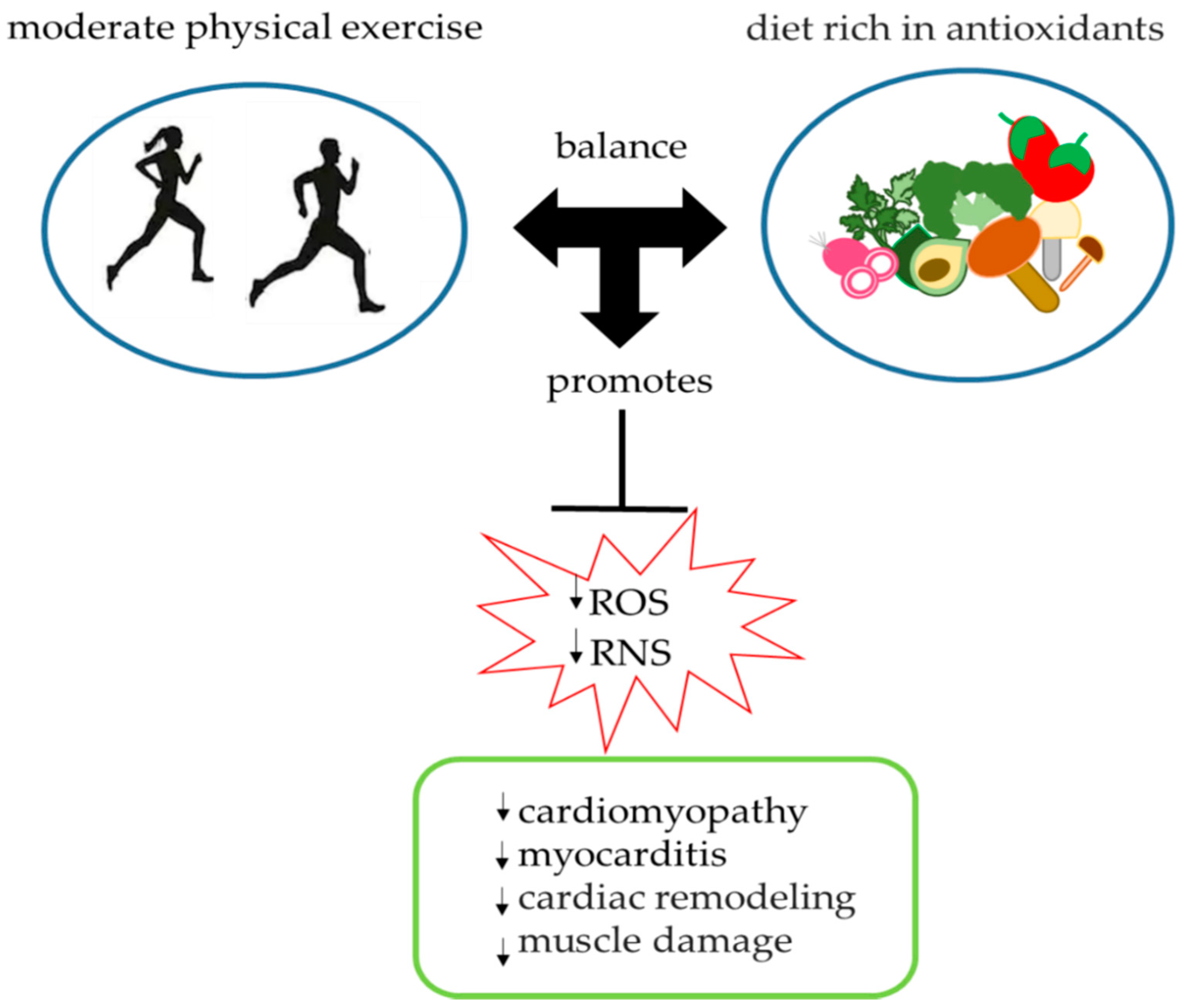
3. Antioxidant Role of Physical Exercise
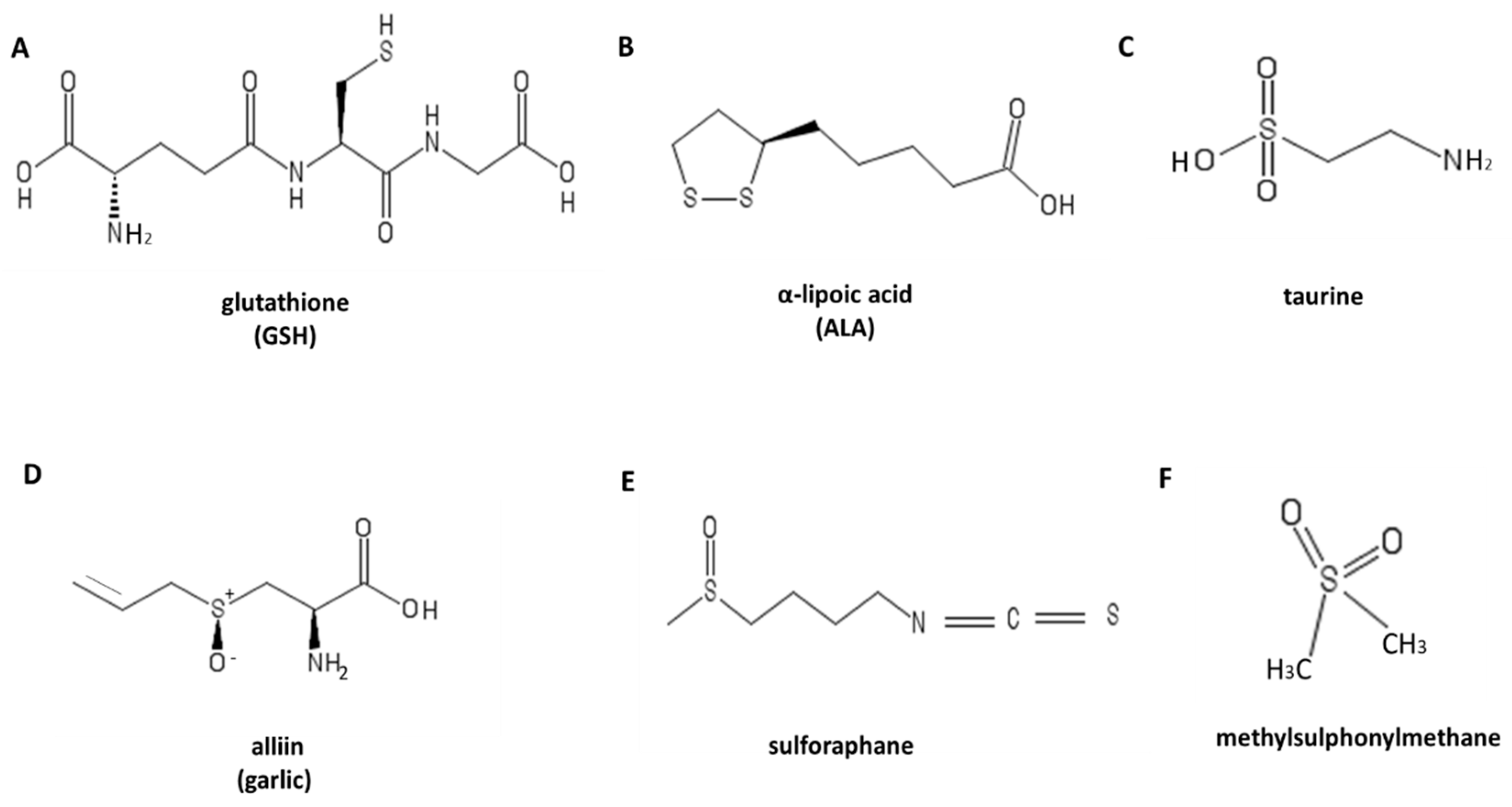
4. Antioxidant: Sulfur-Containing Compounds
4.1. Glutathione
4.2. Lipoic Acid
4.3. Taurine
4.4. Garlic
4.5. Sulforaphane
4.6. Methylsulfonylmethane
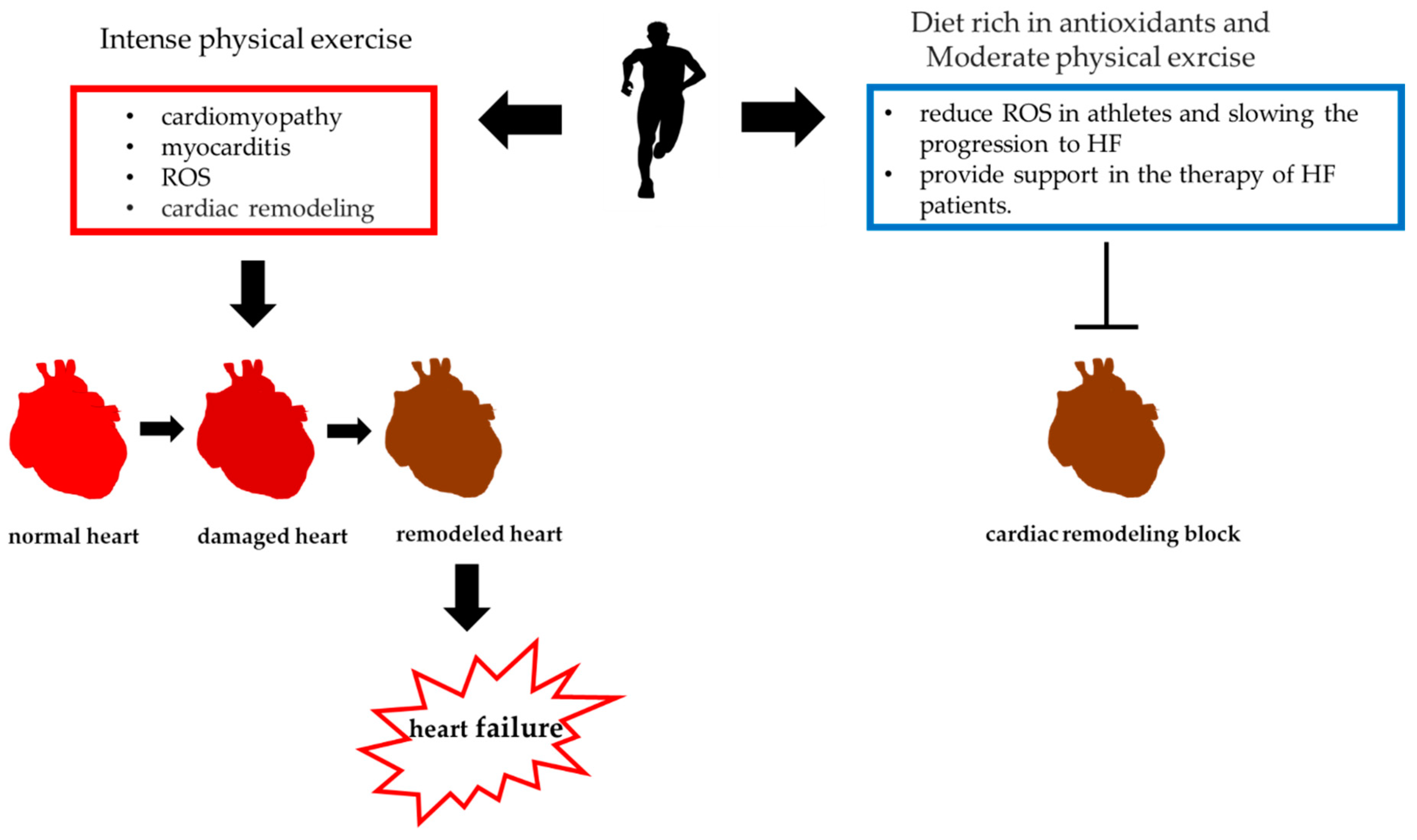
5. Use of Sulfur-Containing Compounds in the Treatment of Heart Failure in Professional Athletes
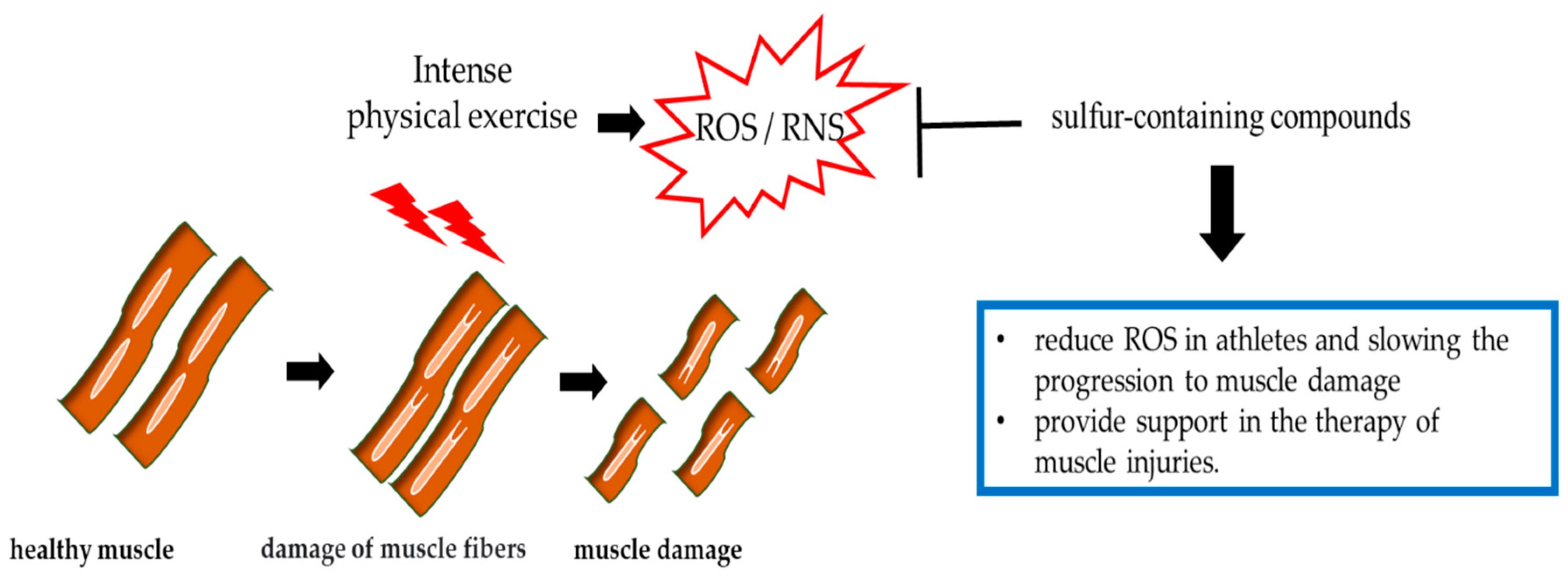
6. Prevention of Muscle Damage by Sulfur-Containing Compounds
7. Conclusions and Future Prospects
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ROS | reactive oxygen species |
| RNS | reactive nitrogen species |
| HF | heart failure |
| MMP | matrix metalloproteinases |
| NADH | nitrate reductase |
| H2O2 | hydrogen peroxide |
| Cys-Sh | Cysteine |
| Cys-SOH | cysteine-sulphenic acid |
| Cys-S-S-Cys | cysteine disulfide |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NOXs | oxidases |
| PLA2 | phospholipase A2 |
| XO | xanthine oxidase |
| TNF-α, | tumor necrosis factor α, |
| IL-1β | interleukin-1β, |
| IL-1ra | interleukin 1 receptor antagonist |
| TNF-R | tumor necrosis factor receptors |
| IL-8 | interleukin-8 |
| MIP-1 | macrophage inflammatory proteins |
| IL-6 | interleukin-6 |
| NF–κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| MDA | malondialdehyde |
| HNE | hydroxynonenal |
| IL-10 | interleukin-10 |
| CoQ10 | coenzyme Q10 |
| SH | sulfhydryl group |
| NO | nitrogen oxide |
| GSH | glutathione |
| GSSG | oxidized glutathione |
| NACN | acetyl cysteine |
| ALA | lipoic acid |
| EPO | erythropoietin |
| 8-iso | 8-isoprostane |
| LPO | lipid peroxides |
| PC | carbonyl proteins |
| NOS | nitric oxide synthase |
| VO2max | maximum oxygen consumption |
| SFN | sulforaphane |
| LDH | lactate dehydrogenase |
| CPK | creatine phosphokinase |
| NQO1 | quinone oxidoreductase 1 |
| NYHA | New York Heart Association |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| MSM | methylsulfonylmethan |
References
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Jiang, Z.; Chen, X.; Liu, M.; Li, J.; Liu, N. The role of autophagy in pro-inflammatory responses of microglia activation via mitochondrial reactive oxygen species in vitro. J. Neurochem. 2017, 142, 215–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Y.; Wang, Y.; Guo, S.; Wang, G. Glutathione peroxidases as oncotargets. Oncotarget 2017, 8, 80093–80102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aseervatham, G.S.B.; Sivasudha, T.; Jeyadevi, R.; Ananth, D.A. Environmental factors and unhealthy lifestyle influence oxidative stress in humans—An overview. Environ. Sci. Pollut. Res. 2013, 20, 4356–4369. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free Radicals, Antioxidants in Disease and Health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef]
- Caggiano, R.; Cattaneo, F.; Moltedo, O.; Esposito, G.; Perrino, C.; Trimarco, B.; Ammendola, R.; Faraonio, R. miR-128 is Implicated in Stress Responses by Targeting MAFG in Skeletal Muscle Cells. Oxidative Med. Cell. Longev. 2017, 2017, 9308310-13. [Google Scholar] [CrossRef] [Green Version]
- Vanzo, A.; Bolner, A.; Nordera, G.; Bosello, O. Obesità, microbiota e stress ossidativo. Biochim. Clin. 2017, 41, 199–207. [Google Scholar]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Polito, R.; Schettino, P.; Grandone, A.; Perrone, L.; Miraglia Del Giudice, E.; Daniele, A. Adiponectin profile and Irisin expression in Italian obese children: Asociation with insulin-resistance. Cytokine 2017, 94, 8–13. [Google Scholar] [CrossRef]
- Giampietro, R.; Spinelli, F.; Contino, M.; Colabufo, N.A. The Pivotal Role of Copper in Neurodegeneration: A New Strategy for the Therapy of Neurodegenerative Disorders. Mol. Pharm. 2018, 15, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhao, W.; Yu, J.; Li, S.; Lin, L.; Chen, X. Induction of ferroptosis and mitochondrial dysfunction by oxidative stress in PC12 cells. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phull, A.-R.; Nasir, B.; ul Haq, I.; Kim, S.J. Oxidative stress, consequences and ROS mediated cellular signaling in rheumatoid arthritis. Chem. Interact. 2018, 281, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Kaffe, E.T.; Rigopoulou, E.I.; Koukoulis, G.K.; Dalekos, G.; Moulas, A.N. Oxidative stress and antioxidant status in patients with autoimmune liver diseases. Redox Rep. 2014, 20, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Carru, C.; Da Boit, M.; Paliogiannis, P.; Zinellu, A.; Sotgia, S.; Sibson, R.; Meakin, J.R.; Aspden, R.M.; Mangoni, A.A.; Gray, S.R. Associations between markers of oxidative stress, skeletal muscle mass and function and to the influence of resistance exercise training, in older adults. Exp. Gerontol. 2018, 103, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, J.; Huang, Z.; Han, J.; Shao, J.; Huang, C. Redox regulation of microRNAs in cancer. Cancer Lett. 2018, 418, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.N.; Sonani, R.R.; Jakharia, K.; Bhastana, B.; Patel, H.M.; Chaubey, M.G.; Singh, N.K.; Madamwar, D. Antioxidant activity and associated structural attributes of Halomicronema phycoerythrin. Int. J. Biol. Macromol. 2018, 111, 359–369. [Google Scholar] [CrossRef]
- Sznarkowska, A.; Kostecka, A.; Meller, K.; Bielawski, K.P. Inhibition of cancer antioxidant defense by natural compounds. Oncotarget 2016, 8, 15996–16016. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Collins, L.B.; Chen, T.-H.; Herr, N.; Takeda, S.; Sun, W.; Swenberg, J.A.; Nakamura, J. Oxidative stress at low levels can induce clustered DNA lesions leading to NHEJ mediated mutations. Oncotarget 2016, 7, 25377–25390. [Google Scholar] [CrossRef]
- Piras, S.; Furfaro, A.L.; Caggiano, R.; Brondolo, L.; Garibaldi, S.; Ivaldo, C.; Marinari, U.M.; Pronzato, M.A.; Faraonio, R.; Nitti, M. MicroRNA-494 Favors HO-1 Expression in Neuroblastoma Cells Exposed to Oxidative Stress in a Bach1-Independent Way. Front. Oncol. 2018, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Verbon, E.H.; Post, J.A.; Boonstra, J. The influence of reactive oxygen species on cell cycle progression in mammalian cells. Gene 2012, 511, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Faraonio, R.; Vergara, P.; Di Marzo, D.; Napolitano, M.; Russo, T.; Cimino, F. Transcription Regulation in NIH3T3 Cell Clones Resistant to Diethylmaleate-Induced Oxidative Stress and Apoptosis. Antioxid. Redox Signal. 2006, 8, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Adraskela, K.; Veisaki, E.; Koutsilieris, M.; Philippou, A. Physical Exercise Positively Influences Breast Cancer Evolution. Clin. Breast Cancer 2017, 17, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, G.M.; Roman, M.G.; Flores, L.C.; Hubbard, G.B.; Salmon, A.B.; Zhang, Y.; Gelfond, J.; Ikeno, Y. The paradoxicalrole of thioredoxin on oxidative stress and aging. Arch. Biochem. Biophys. 2015, 576, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Tenkorang, M.A.; Snyder, B.; Cunningham, R.L. Sex-related differences in oxidative stress and neurodegeneration. Steroids 2018, 133, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Trabucco, S.E.; Zhang, H. Oxidative Stress, Mitochondrial Dysfunction and the Mitochondria Theory of Aging. Cancer Aging 2014, 39, 86–107. [Google Scholar] [CrossRef]
- Ragozzino, E.; Brancaccio, M.; Di Costanzo, A.; Scalabrì, F.; Andolfi, G.; Wanderlingh, L.G.; Patriarca, E.J.; Minchiotti, G.; Altamura, S.; Varrone, F.; et al. 6-Bromoindirubin-3’-oxime intercepts GSK3 signaling to promote and enhance skeletal muscle differentiation affecting miR-206 expression in mice. Sci. Rep. 2019, 9, 18091. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, H.; Kinugawa, S.; Matsushima, S. Oxidative stress and heart failure. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2181–H2190. [Google Scholar] [CrossRef] [Green Version]
- Link, M.S.; Estes, N.A. Sudden cardiac death in the athlete: Bridging the gaps between evidence, policy, and practice. Circulation 2012, 125, 2511–2516. [Google Scholar] [CrossRef]
- Harmon, K.G.; Asif, I.M.; Maleszewski, J.J.; Owens, D.S.; Prutkin, J.M.; Salerno, J.C.; Zigman, M.L.; Ellenbogen, R.; Rao, A.L.; Ackerman, M.J.; et al. Incidence, Cause, and Comparative Frequency of Sudden Cardiac Death in National Collegiate Athletic Association Athletes: A Decade in Review. Circulation 2015, 132, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljungqvist, A.; Jenoure, P.J.; Engebretsen, L.; Alonso, J.M.; Bahr, R.; Clough, A.F.; De Bondt, G.; Dvorak, J.; Maloley, R.; Matheson, G.; et al. The International Olympic Committee (IOC) Consensus Statement on Periodic Health Evaluation of Elite Athletes, March 2009. Clin. J. Sport Med. 2009, 19, 347–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hainline, B.; Drezner, J.A.; Baggish, A.; Harmon, K.G.; Emery, M.S.; Myerburg, R.J.; Sanchez, E.; Molossi, S.; Parsons, J.T.; Thompson, P.D. Interassociation Consensus Statement on Cardiovascular Care of College Student-Athletes. J. Am. Coll. Cardiol. 2016, 51, 344–357. [Google Scholar]
- Delos, D.; Maak, T.G.; Rodeo, S.A. Muscle Injuries in Athletes. Sports Health A Multidiscip. Approach 2013, 5, 346–352. [Google Scholar] [CrossRef]
- Tscholl, P.; Meynard, T.; Le Thanh, N.; Neroladaki, A. Diagnostics and classification of muscle injuries in sport. Swiss Sports Exerc. Med. 2018, 67, 8–15. [Google Scholar]
- Höhn, A.; König, J.; Jung, T. Metabolic Syndrome, Redox State, and the Proteasomal System. Antioxid. Redox Signal. 2016, 25, 902–917. [Google Scholar] [CrossRef]
- Peternelj, T.T.; Coombes, J.S. Antioxidant supplementation during exercise training: Beneficial or detrimental? Sports Med. 2011, 41, 1043–1069. [Google Scholar] [CrossRef]
- Srinivas, U.S.; Tan, B.W.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Evans, M.D.; Cooke, M.S. Oxidative Damage to DNA in Non-Malignant Disease: Biomarker or Biohazard? Genome Dyn. 2006, 1, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Cecarini, V.; Gee, J.; Fioretti, E.; Amici, M.; Angeletti, M.; Eleuteri, A.M.; Keller, J.N. Protein oxidation and cellular homeostasis: Emphasis on metabolism. Biochim. Biophys. Acta (BBA) Bioenerg. 2007, 1773, 93–104. [Google Scholar] [CrossRef]
- Castell, L.M.; Poortmans, J.R.; Leclercq, R.; Brasseur, M.; Duchateau, J.; Newsholme, E.A. Some aspects of the acute phase response after a marathon race, and the effects of glutamine supplementation. Graefe’s Arch. Clin. Exp. Ophthalmol. 1996, 75, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Hellsten, Y.; Frandsen, U.; Orthenblad, N.; Sjødin, B.; Richter, E.A. Xanthine oxidase in human skeletal muscle following eccentric exercise: A role in inflammation. J. Physiol. 1997, 498, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Steensberg, A.; Fischer, C.; Keller, C.; Ostrowski, K.; Schjerling, P. Exercise and cytokines with particular focus on muscle-derived IL-6. Exerc. Immunol. Rev. 2001, 7, 18. [Google Scholar] [PubMed]
- Steinbacher, P.; Eckl, P. Impact of Oxidative Stress on Exercising Skeletal Muscle. Biomolecules 2015, 5, 356–377. [Google Scholar] [CrossRef] [PubMed]
- Mahli, A.; Thasler, W.E.; Patsenker, E.; Müller, S.; Stickel, F.; Müller, M.; Seitz, H.K.; Cederbaum, A.I.; Hellerbrand, C. Identification of cytochrome CYP2E1 as critical mediator of synergistic effects of alcohol and cellular lipid accumulation in hepatocytes in vitro. Oncotarget 2015, 6, 41464–41478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, G.; Bruno, R.; Taranto, A.; Martino, G. Are Polyunsaturated Fatty Acid Metabolites, the Protective Effect of 4-hydroxytyrosol on Human Red Blood Cell Membranes and Oxidative Damage (4-hydroxyalkenals) Compatible in Hypertriglyceridemic Patients? Pharmacogn. Mag. 2017, 13, S561–S566. [Google Scholar] [CrossRef]
- Anthonymuthu, T.-S.; Kim-Campbell, N.; Bayir, H. Oxidative lipidomics: Applications in critical care. Curr. Opin. Crit. Care 2017, 23, 251–256. [Google Scholar] [CrossRef]
- Kuburovic, V.; Vekic, J.; Zeljkovic, A.; Carrié, A.; Kotur-Stevuljevic, J.; Bojanin, D.; Kosutic, J.; Spasojevic-Kalimanovska, V.; Miljkovic, M.; Kuburovic, N.; et al. The usefulness of advanced lipid and oxidative stress testing for diagnosis and management of low HDL-cholesterol phenotype: A case report. Clin. Biochem. 2017, 50, 1323–1325. [Google Scholar] [CrossRef]
- Kim, D.H.; Kwack, S.J.; Yoon, K.S.; Choi, J.S.; Lee, B.M. 4-Hydroxynonenal: A Superior Oxidative Biomarker Compared to Malondialdehyde and Carbonyl Content Induced by Carbon Tetrachloride in Rats. J. Toxicol. Environ. Health Part A 2015, 78, 1051–1062. [Google Scholar] [CrossRef]
- Mao, L.; Zuo, M.-L.; Hu, G.-H.; Duan, X.-M.; Yang, Z.-B. Mir-193 targets ALDH2 and contributes to toxic aldehyde accumulation and tyrosine hydroxylase dysfunction in cerebral ischemia/reperfusion injury. Oncotarget 2017, 8, 99681–99692. [Google Scholar] [CrossRef] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Ruchi, T.; Chakraborty, S.; Dhama, K. Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. BioMed Res. Int. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scudiero, O.; Brancaccio, M.; Mennitti, C.; Laneri, S.; Lombardo, B.; De Biasi, M.G.; De Gregorio, E.; Pagliuca, C.; Colicchio, R.; Salvatore, P.; et al. Human Defensins: A Novel Approach in the Fight against Skin Colonizing Staphylococcus Aureus. Antibiotics 2020, 9, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pero, R.; Brancaccio, M.; Mennitti, C.; Gentile, L.; Franco, A.; Laneri, S.; De Biasi, M.G.; Pagliuca, C.; Colicchio, R.; Salvatore, P.; et al. HNP-1 and HBD-1 as Biomarkers for the Immune Systems of Elite Basketball Athletes. Antibiotics 2020, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, M.; Mennitti, C.; Laneri, S.; Franco, A.; De Biasi, M.G.; Cesaro, A.; Fimiani, F.; Moscarella, E.; Gragnano, F.; Mazzaccara, C.; et al. Methicillin-Resistant Staphylococcus Aureus: Risk for General Infection and Endocarditis among Athletes. Antibiotics 2020, 9, 332. [Google Scholar] [CrossRef] [PubMed]
- Emmendoerffer, A.; Hecht, M.; Boeker, T.; Mueller, M.; Heinrich, U. Role of inflammation in chemical-induced lung cancer. Toxicol. Lett. 2000, 112, 185–191. [Google Scholar] [CrossRef]
- Pero, R.; Brancaccio, M.; Laneri, S.; De Biasi, M.G.; Lombardo, B.; Scudiero, O. A Novel View of Human Helicobacter Pylori Infections: Interplay between Microbiota and Beta-Defensins. Biomolecules 2019, 9, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAnulty, S.R.; McAnulty, L.S.; Morrow, J.D.; Nieman, D.C.; Owens, J.T.; Carper, C.M. Influence of carbohydrate, intense exercise, and rest intervals on hormonal and oxidative changes. Int. J. Sport Nutr. Exerc. Metab. 2007, 17, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-Induced Oxidative Stress: Cellular Mechanisms and Impact on Muscle Force Production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [Green Version]
- Bouzid, M.A.; Filaire, E.; Matran, R.; Robin, S.; Fabre, C. Lifelong Voluntary Exercise Modulates Age-Related Changes in Oxidative Stress. Int. J. Sports Med. 2018, 39, 21–28. [Google Scholar] [CrossRef]
- Baltaci, S.B.; Mogulkoc, R.; Baltaci, A.K. Resveratrol and exercise. Biomed. Rep. 2016, 5, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.K. Muscles and their myokines. J. Exp. Biol. 2011, 214, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golbidi, S.; Badran, M.; Laher, I. Antioxidant and Anti-Inflammatory Effects of Exercise in Diabetic Patients. Exp. Diabetes Res. 2011, 2012, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentinger, M.; Brismar, K.; Dallner, G. The antioxidant role of coenzyme Q. Mitochondrion 2007, 7, S41–S50. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Nieman, D.C.; Pedersen, B.K. Exercise, nutrition and immune function. J. Sports Sci. 2004, 22, 115–125. [Google Scholar] [CrossRef]
- Pingitore, A.; Lima, G.P.P.; Mastorci, F.; Quinones, A.; Iervasi, G.; Vassalle, C. Exercise and oxidative stress: Potential effects of antioxidant dietary strategies in sports. Nutrition 2015, 31, 916–922. [Google Scholar] [CrossRef]
- Kawamura, T.; Muraoka, I.; Kawamura, T.; Muraoka, I. Exercise-Induced Oxidative Stress and the Effects of Antioxidant Intake from a Physiological Viewpoint. Antioxidants 2018, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- McLeay, Y.; Stannard, S.R.; Houltham, S.; Starck, C. Dietary thiols in exercise: Oxidative stress defence, exercise performance, and adaptation. J. Int. Soc. Sports Nutr. 2017, 14, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jacob, C. A scent of therapy: Pharmacological implications of natural products containing redox-active sulphur atoms. Nat. Prod. Rep. 2006, 23, 851–863. [Google Scholar] [CrossRef]
- Dunbar, K.L.; Scharf, D.H.; Litomska, A.; Hertweck, C. Enzymatic Carbon–Sulfur Bond Formation in Natural Product Biosynthesis. Chem. Rev. 2017, 117, 5521–5577. [Google Scholar] [CrossRef]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free. Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Seifirad, S.; Ghaffari, A.; Amoli, M.M. The antioxidants dilemma: Are they potentially immunosuppressants and carcinogens? Front. Physiol. 2014, 5, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.W.; Ryu, J.H.; Kim, S.G. The Essential Role of Phosphatidylinositol 3-Kinase and of p38 Mitogen-Activated Protein Kinase Activation in the Antioxidant Response Element-Mediated rGSTA2 Induction by Decreased Glutathione in H4IIE Hepatoma Cells. Mol. Pharmacol. 2000, 58, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- You, B.R.; Kim, S.H.; Park, W.H. Reactive oxygen species, glutathione, and thioredoxin influence suberoyl bishydroxamic acid-induced apoptosis in A549 lung cancer cells. Tumor Biol. 2014, 36, 3429–3439. [Google Scholar] [CrossRef] [PubMed]
- Herzenberg, L.A.; De Rosa, S.C.; Dubs, J.G.; Roederer, M.; Anderson, M.T.; Ela, S.W.; Deresinski, S.C. Glutathione deficiency is associated with impaired survival in HIV disease. Proc. Natl. Acad. Sci. USA 1997, 94, 1967–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmay, M.J.; Terpstra, M.; Coles, L.D.; Mishra, U.; Ahlskog, M.; Öz, G.; Cloyd, J.C.; Tuite, P.J. N-acetylcysteine Boosts Brain and Blood Glutathione in Gaucher and Parkinson Diseases. Clin. Neuropharmacol. 2013, 36, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Bridgeman, M.M.; Marsden, M.; MacNee, W.; Flenley, D.C.; Ryle, A.P. Cysteine and glutathione concentrations in plasma and bronchoalveolar lavage fluid after treatment with N-acetylcysteine. Thorax 1991, 46, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowska, D.; Grucza, K.; Chołbiński, P.; Szutowski, M. Effects of Supplementation with Glutathione and its Precursors on Athlete Performance. Biomed. J. Sci. Tech. Res. 2019, 12, 001–008. [Google Scholar] [CrossRef]
- Medved, I.; Brown, M.J.; Bjorksten, A.R.; Murphy, K.T.; Petersen, A.C.; Sostaric, S.; Gong, X.; McKenna, M.J. N-acetylcysteine enhances muscle cysteine and glutathione availability and attenuates fatigue during prolonged exercise in endurance-trained individuals. J. Appl. Physiol. 2004, 97, 1477–1485. [Google Scholar] [CrossRef]
- Michailidis, Y.; Karagounis, L.G.; Terzis, G.; Jamurtas, A.Z.; Spengos, K.; Tsoukas, D.; Chatzinikolaou, A.; Mandalidis, D.; Stefanetti, R.J.; Papassotiriou, I.; et al. Thiol-based antioxidant supplementation alters human skeletal muscle signaling and attenuates its inflammatory response and recovery after intense eccentric exercise. Am. J. Clin. Nutr. 2013, 98, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Terrill, J.R.; Pinniger, G.J.; Graves, J.A.; Grounds, M.D.; Arthur, P.G. Increasing taurine intake and taurine synthesis improves skeletal muscle function in the mdx mouse model for Duchenne muscular dystrophy. J. Physiol. 2016, 594, 3095–3110. [Google Scholar] [CrossRef] [Green Version]
- Durrani, A.I.; Schwartz, H.; Nagl, M.; Sontag, G. Determination of free α-lipoic acid in foodstuffs by HPLC coupled with CEAD and ESI-MS. Food Chem. 2010, 120, 1143–1148. [Google Scholar] [CrossRef]
- Zembron-Lacny, A.; Morawin, B.; Gajewski, M.; Naczk, M.; Siatkowski, I.; Turowski, D. The Combination of α-lipoic acid intake with eccentric exercise modulates erythropoietin release. Biol. Sport 2014, 31, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffer, S.; Kim, H.W. Effects and Mechanisms of Taurine as a Therapeutic Agent. Biomol. Ther. 2018, 26, 225–241. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, F.G.; Galan, B.S.M.; Santos, P.C.; Pritchett, K.; Pfrimer, K.; Ferriolli, E.; Papoti, M.; Marchini, J.S.; De Freitas, E.C. Taurine: A Potential Ergogenic Aid for Preventing Muscle Damage and Protein Catabolism and Decreasing Oxidative Stress Produced by Endurance Exercise. Front. Physiol. 2017, 8, 710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charron, C.S.; Dawson, H.D.; Novotny, J.A. Garlic Influences Gene Expression in Vivo and in Vitro. J. Nutr. 2016, 146, 444S–449S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ince, I.D.; Sonmez, G.T.; Ince, M.M. Effects of Garlic on Aerobic Performance. Turk. J. Med. Sci. 2000, 3, 557–561. [Google Scholar]
- Malaguti, M.; Angeloni, C.; Garatachea, N.; Baldini, M.; Leoncini, E.; Collado, P.S.; Teti, G.; Falconi, M.; González-Gallego, J.; Hrelia, S.; et al. Sulforaphane treatment protects skeletal muscle against damage induced by exhaustive exercise in rats. J. Appl. Physiol. 2009, 107, 1028–1036. [Google Scholar] [CrossRef] [Green Version]
- Pearson, T.W.; Dawson, H.J.; Lackey, H.B. Naturally occurring levels of dimethyl sulfoxide in selected fruits, vegetables, grains, and beverages. J. Agric. Food Chem. 1981, 29, 1089–1091. [Google Scholar] [CrossRef]
- Clark, T.; Murray, J.S.; Lane, P.; Politzer, P. Why are dimethyl sulfoxide and dimethyl sulfone such good solvents? J. Mol. Model. 2008, 14, 689–697. [Google Scholar] [CrossRef]
- Ahn, H.; Kim, J.; Lee, M.-J.; Kim, Y.J.; Cho, Y.-W.; Lee, G.-S. Methylsulfonylmethane inhibits NLRP3 inflammasome activation. Cytokine 2015, 71, 223–231. [Google Scholar] [CrossRef]
- Coleman, J.W. Nitric oxide: A regulator of mast cell activation and mast cell-mediated inflammation. Clin. Exp. Immunol. 2002, 129, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Butawan, M.; Benjamin, R.L.; Bloomer, R.J. Methylsulfonylmethane: Applications and Safety of a Novel Dietary Supplement. Nutrition 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8, S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalman, D.; Feldman, S.; Samson, A.; Krieger, D. A Randomized Double Blind Placebo Controlled Evaluation of MSM for Exercise Induced Discomfort/Pain. J. Int. Soc. Sports Nutr. 2017, 14, 24. [Google Scholar]
- Maron, B.J.; Pelliccia, A. The Heart of Trained Athletes: Cardiac Remodeling and the Risks of Sports, including Sudden Death. Circulation 2006, 114, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Abernethy, W.B.; Choo, J.K.; Hutter, A.M. Echocardiographic characteristics of professional football players. J. Am. Coll. Cardiol. 2003, 41, 280–284. [Google Scholar] [CrossRef]
- Pelliccia, A.; Maron, B.J.; De Luca, R.; Di Paolo, F.M.; Spataro, A.; Culasso, F. Remodeling of Left Ventricular Hypertrophy in Elite Athletes after Long-Term Deconditioning. Circulation 2002, 105, 944–949. [Google Scholar] [CrossRef] [Green Version]
- Carbone, A.; D’Andrea, A.; Riegler, L.; Scarafile, R.; Pezzullo, E.; Martone, F.; America, R.; Liccardo, B.; Gaderisi, M.; Bossone, E.; et al. Cardiac damage in athlete’s heart: When the “supernormal” heart fails! World J. Cardiol. 2017, 9, 470. [Google Scholar] [CrossRef]
- Chen, C.; Sung, K.-T.; Shih, S.-C.; Liu, C.-C.; Kuo, J.-Y.; Hou, C.J.-Y.; Hung, C.-L.; Yeh, H.-I. Age, Gender and Load-Related Influences on Left Ventricular Geometric Remodeling, Systolic Mid-Wall Function, and NT-ProBNP in Asymptomatic Asian Population. PLoS ONE 2016, 11, e0156467. [Google Scholar] [CrossRef]
- Deligiannis, A.; Björnstad, H.; Carre, F.; Heidbüchel, H.; Kouidi, E.; Panhuyzen-Goedkoop, N.M.; Pigozzi, F.; Schänzer, W.; Vanhees, L.; ESC Study Group of Sports Cardiology. ESC Study Group of Sports Cardiology Position Paper on adverse cardiovascular effects of doping in athletes. Eur. J. Cardiovasc. Prev. Rehabil. 2006, 13, 687–694. [Google Scholar] [CrossRef]
- Corrado, D.; Pelliccia, A.; Bjørnstad, H.H.; Vanhees, L.; Biffi, A.; Borjesson, M.; Panhuyzen-Goedkoop, N.; Deligiannis, A.; Solberg, E.; Dugmore, D.; et al. Cardiovascular pre-participation screening of young competitive athletes for prevention of sudden death: Proposal for a common European protocol. Eur. Heart J. 2005, 26, 516–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girolami, F.; Frisso, G.; Benelli, M.; Crotti, L.; Iascone, M.; Mango, R.; Mazzaccara, C.; Pilichou, K.; Arbustini, E.; Tomberli, B.; et al. Contemporary genetic testing in inherited cardiac disease: Tools, ethical issues, and clinical applications. J. Cardiovasc. Med. (Hagerstown). 2018, 19, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detta, N.; Frisso, G.; Limongelli, G.; Marzullo, M.; Calabrò, R.; Salvatore, F. Genetic analysis in a family affected by sick sinus syndrome may reduce the sudden death risk in a young aspiring competitive athlete. Int. J. Cardiol. 2014, 170, e63–e65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadžović-Džuvo, A.; Valjevac, A.; Lepara, O.; Pjanić, S.; Hadžimuratović, A.; Mekić, A. Oxidative stress status in elite athletes engaged in different sport disciplines. Bosn. J. Basic. Med. Sci. 2014, 14, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Dhalla, A.K.; Seneviratne, C.; Singal, P.K. Oxidative stress and heart failure. Mol. Cell. Biochem. 1995, 147, 77–81. [Google Scholar] [CrossRef]
- Keith, M.; Geranmayegan, A.; Sole, M.J.; Kurian, R.; Robinson, A.; Omran, A.S.; Jeejeebhoy, K.N. Increased Oxidative Stress in Patients with Congestive Heart Failure. J. Am. Coll. Cardiol. 1998, 31, 1352–1356. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Prieto, M.A.; Miatello, R.M. Organosulfur compounds and cardiovascular disease. Mol. Asp. Med. 2010, 31, 540–545. [Google Scholar] [CrossRef]
- Gu, X.; Zhu, Y.Z. Therapeutic applications of organosulfur compounds as novel hydrogen sulfide donors and/or mediators. Expert Rev. Clin. Pharmacol. 2011, 4, 123–133. [Google Scholar] [CrossRef]
- Bajic, V.P.; Van Neste, C.; Obradovic, M.; Zafirovic, S.; Radak, D.; Bajic, V.B.; Essack, M.; Isenovic, E.R. Glutathione “Redox Homeostasis” and its Relation to Cardiovascular Disease. Oxid. Med. Cell Longev. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Paschalis, V.; Theodorou, A.A.; Margaritelis, N.V.; Kyparos, A.; Nikolaidis, M.G. N-acetylcysteine supplementation increases exercise performance and reduces oxidative stress only in individuals with low levels of glutathione. Free. Radic. Biol. Med. 2018, 115, 288–297. [Google Scholar] [CrossRef]
- Damy, T.; Kirsch, M.; Khouzami, L.; Caramelle, P.; Le Corvoisier, P.; Roudot-Thoraval, F.; Dubois-Randé, J.-L.; Hittinger, L.; Pavoine, C.; Pecker, F. Glutathione Deficiency in Cardiac Patients is Related to the Functional Status and Structural Cardiac Abnormalities. PLoS ONE 2009, 4, e4871. [Google Scholar] [CrossRef] [PubMed]
- Adamy, C.; Mulder, P.; Khouzami, L.; Andrieu-Abadie, N.; Defer, N.; Candiani, G.; Pavoine, C.; Caramelle, P.; Souktani, R.; Le Corvoisier, P.; et al. Neutral sphingomyelinase inhibition participates to the benefits of N-acetylcysteine treatment in post-myocardial infarction failing heart rats. J. Mol. Cell. Cardiol. 2007, 43, 344–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giam, B.; Chu, P.; Kuruppu, S.; Smith, A.I.; Horlock, D.; Kiriazis, H.; Du, X.; Kaye, D.M.; Rajapakse, N.W. N-acetylcysteine attenuates the development of cardiac fibrosis and remodeling in a mouse model of heart failure. Physiol. Rep. 2016, 4, e12757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, A.J.; Berg, H.M. Effect of taurine on sarcoplasmic reticulum function and force in skinned fast-twitch skeletal muscle fibres of the rat. J. Physiol. 2002, 538, 185–194. [Google Scholar] [CrossRef]
- Liu, Z.; Qi, B.; Zhang, M.; Izumi, I.; Kagamimori, S.; Sokejima, S.; Yamagami, T. Role of taurine supplementation to prevent exercise-induced oxidative stress in healthy young men. Amino Acids 2004, 26, 203–207. [Google Scholar] [CrossRef]
- Ahmadian, M.; Roshan, V.D.; Ashourpore, E. Taurine Supplementation Improves Functional Capacity, Myocardial Oxygen Consumption, and Electrical Activity in Heart Failure. J. Diet. Suppl. 2017, 14, 422–432. [Google Scholar] [CrossRef]
- Beyranvand, M.R.; Khalafi, M.K.; Roshan, V.D.; Choobineh, S.; Parsa, S.A.; Piranfar, M.A. Effect of taurine supplementation on exercise capacity of patients with heart failure. J. Cardiol. 2011, 57, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Schaffer, S.; Azuma, J. The effect of taurine on chronic heart failure: Actions of taurine against catecholamine and angiotensin II. Amino Acids 2014, 46, 111–119. [Google Scholar] [CrossRef]
- Dhalla, N.S.; Adameová, A.; Kaur, M. Role of catecholamine oxidation in sudden cardiac death. Fundam. Clin. Pharmacol. 2010, 24, 539–546. [Google Scholar] [CrossRef]
- Hanna, J.; Chahine, R.; Aftimos, G.; Nader, M.; Mounayar, A.; Esseily, F.; Chamat, S. Protective effect of taurine against free radicals damage in the rat myocardium. Exp. Toxicol. Pathol. 2004, 56, 189–194. [Google Scholar] [CrossRef]
- Querques, F.; Cantilena, B.; Cozzolino, C.; Esposito, M.T.; Passaro, F.; Parisi, S.; Lombardo, B.; Russo, T.; Pastore, L. Angiotensin receptor I stimulates osteoprogenitor proliferation through TGFβ-mediated signaling. J. Cell. Physiol. 2015, 230, 1466–1474. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Azuma, M.; Taira, K.; Baba, A.; Yamamoto, I.; Schaffer, S.W.; Azuma, J. Effect of Taurine on Angiotensin II-Induced Hypertrophy of Neonatal Rat Cardiac Cells. J. Cardiovasc. Pharmacol. 1997, 30, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Dudek, M.; Bilska-Wilkosz, A.; Knutelska, J.; Mogilski, S.; Bednarski, M.; Zygmunt, M.; Iciek, M.; Sapa, J.; Bugajski, D.; Filipek, B.; et al. Are anti-inflammatory properties of lipoic acid associated with the formation of hydrogen sulfide? Pharmacol. Rep. 2013, 65, 1018–1024. [Google Scholar] [CrossRef]
- Skibska, B.; Goraca, A. The Protective Effect of Lipoic Acid on Selected Cardiovascular Diseases Caused by Age-Related Oxidative Stress. Oxidative Med. Cell. Longev. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, A.M.; Davison, G.W.; Murphy, M.H.; Nadeem, N.; Trinick, T.; Duly, E.; Novials, A.; McEneny, J. Effect of α-lipoic acid and exercise training on cardiovascular disease risk in obesity with impaired glucose tolerance. Lipids Health Dis. 2011, 10, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packer, L.; Cadenas, E. Lipoic acid: Energy metabolism and redox regulation of transcription and cell signaling. J. Clin. Biochem. Nutr. 2010, 48, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Wollin, S.D.; Jones, P.J.H. α-Lipoic Acid and Cardiovascular Disease. J. Nutr. 2003, 133, 3327–3330. [Google Scholar] [CrossRef]
- Guerrero-Beltrán, C.E.; Calderón-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective effect of sulforaphane against oxidative stress: Recent advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef]
- Angeloni, C.; Leoncini, E.; Malaguti, M.; Angelini, S.; Hrelia, P.; Hrelia, S. Modulation of Phase II Enzymes by Sulforaphane: Implications for its Cardioprotective Potential. J. Agric. Food Chem. 2009, 57, 5615–5622. [Google Scholar] [CrossRef]
- Ma, T.; Zhu, D.; Chen, D.; Zhang, Q.; Dong, H.; Wu, W.; Lu, H.; Wu, G. Sulforaphane, a Natural Isothiocyanate Compound, Improves Cardiac Function and Remodeling by Inhibiting Oxidative Stress and Inflammation in a Rabbit Model of Chronic Heart Failure. Med. Sci. Monit. 2018, 24, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Wang, X.; Zhao, S.; Ma, C.; Cui, J.; Zheng, Y. Sulforaphane Protects against Cardiovascular Disease via Nrf2 Activation. Oxidative Med. Cell. Longev. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, Y.; Bai, Y.; Jiang, X.; Zhou, S.; Wang, Y.; Wintergerst, K.A.; Cui, T.; Ji, H.; Tan, Y.; Cai, L. Sulforaphane prevents angiotensin II-induced cardiomyopathy by activation of Nrf2 via stimulating the Akt/GSK-3ß/Fyn pathway. Redox Biol. 2018, 15, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.; Budoff, M.J. Garlic and Heart Disease. J. Nutr. 2016, 146, 416S–421S. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, G.; Cong, X.; Wen, C. Black Garlic Improves Heart Function in Patients with Coronary Heart Disease by Improving Circulating Antioxidant Levels. Front. Physiol. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.; Ottosson, F.; Hellstrand, S.; Ericson, U.; Orho-Melander, M.; Fernandez, C.; Melander, O. Ergothioneine is associated with reduced mortality and decreased risk of cardiovascular disease. Heart 2019, 106, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Cheah, I.K.; Tang, R.M.Y. Ergothioneine—A diet-derived antioxidant with therapeutic potential. FEBS Lett. 2018, 592, 3357–3366. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.Y.; Li, H.; Lim, H.J.; Lee, H.J.; Jeon, R.; Ryu, J.-H. Anti-Inflammatory Activity of Sulfur-Containing Compounds from Garlic. J. Med. Food 2012, 15, 992–999. [Google Scholar] [CrossRef] [Green Version]
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.G.F.; Coats, A.J.S.; Falk, V.; Gonzalez-Juanatey, J.R.; Harjola, V.P.; Jankowska, E.A.; et al. ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J. 2016, 37, 2129–2200. [Google Scholar] [CrossRef]
- Nakhostin-Roohi, B.; Niknam, Z.; Vaezi, N.; Mohammadi, S.; Bohlooli, S. Effect of Single Dose Administration of Methylsulfonylmethane on Oxidative Stress following Acute Exhaustive Exercise. Iran. J. Pharm. Res. 2013, 12, 845–853. [Google Scholar]
- Reid, M.B.; Stokić, D.S.; Koch, S.M.; Khawli, F.A.; Leis, A.A. N-acetylcysteine inhibits muscle fatigue in humans. J. Clin. Investig. 1994, 94, 2468–2474. [Google Scholar] [CrossRef]
- Di Meo, S.; Napolitano, G.; Venditti, P. Mediators of Physical Activity Protection against ROS-Linked Skeletal Muscle Damage. Int. J. Mol. Sci. 2019, 20, 3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maughan, R.J.; Donnelly, A.E.; Gleeson, M.; Whiting, P.H.; Walker, K.A.; Clough, P.J. Delayed-onset muscle damage and lipid peroxidation in man after a downhill run. Muscle Nerve 1989, 12, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Reznick, A.Z.; Witt, E.; Matsumoto, M.; Packer, L. Vitamin E inhibits protein oxidation in skeletal muscle of resting and exercised rats. Biochem. Biophys. Res. Commun. 1992, 189, 801–806. [Google Scholar] [CrossRef]
- Fielding, R.A.; Manfredi, T.J.; Ding, W.; Fatarone, M.; Evans, M.J.; Cannon, J.G. Acute response to exercise III. Neutrophil and IL-1ß accumulation in skeletal muscle. Am. J. Physiol. 1993, 265, R166–R172. [Google Scholar] [PubMed]
- Morillas-Ruiz, J.; Zafrilla, P.; Almar, M.; Cuevas, M.J.; Lopez, F.J.; Abellan, P.; Villegas, J.A.; Gonza lez-Gallego, J. The effects of an antioxidant-supplemented beverage on exercise-induced oxidative stress: Results from a placebo-controlled double-blind study in cyclists. Eur. J. Appl. Physiol. 2005, 95, 543–549. [Google Scholar] [CrossRef]
- Lombardo, B.; Izzo, V.; Terracciano, D.; Ranieri, A.; Mazzaccara, C.; Fimiani, F.; Cesaro, A.; Gentile, L.; Leggiero, E.; Pero, R.; et al. Laboratory medicine: Health evaluation in elite athletes. Clin. Chem. Lab. Med. 2019, 57, 1450–1473. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.-G.; Ahn, B.-W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]
- Moini, H.; Packer, L.; Saris, N.E.L. Antioxidant and prooxidant of α-lipoic acid and dihydrolipoic acid. Toxicol. Appl. Pharmacol. 2002, 182, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Zembron-Lacny, A. Effects of Sulphur-Containing Compounds on Plasma Redox Status in Muscle-Damaging Exercise. Chin. J. Physiol. 2009, 52, 289–294. [Google Scholar] [CrossRef]
- Çakatay, U. Pro-oxidant actions of α-lipoic acid and dihydrolipoic acid. Med. Hypotheses 2006, 66, 110–117. [Google Scholar] [CrossRef]
- Childs, A.; Jacobs, C.; Kaminski, T.; Halliwell, B.; Leeuwenburgh, C. Supplementation with vitamin C and N-acetyl-cysteine increases oxidative stress in humans after an acute muscle injury induced by eccentric exercise. Free. Radic. Biol. Med. 2001, 31, 745–753. [Google Scholar] [CrossRef]
- Kim, L.S.; Axelrod, L.; Howard, P.; Buratovich, N.; Waters, R. Efficacy of methylsulfonylmethane (MSM) in osteoarthritis pain of the knee: A pilot clinical trial. Osteoarthr. Cartil. 2006, 14, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnuson, B.A.; Appleton, J.; Ames, G.B. Pharmacokinetics and Distribution of [35S]Methylsulfonylmethane following Oral Administration to Rats. J. Agric. Food Chem. 2007, 55, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Barrager, E.; Veltmann, J.R.; Schauss, A.G., Jr. A multicentered, open-label trial on the safety and efficacy of methylsulfonylmethane in the treatment of seasonal allergic rhinitis. J. Altern. Complement. Med. 2002, 8, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, J.I.; Siegel, B.V. Effects of Oral Dimethyl Sulfoxide and Dimethyl Sulfone on Murine Autoimmune Lymphoproliferative Disease. Exp. Biol. Med. 1986, 183, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Nakhostin-Roohi, B.; Barmaki, S.; Khoshkhahesh, F.; Bohlooli, S. Effect of chronic supplementation with methylsulfonylmethane on oxidative stress following acute exercise in untrained healthy men. J. Pharm. Pharmacol. 2011, 63, 1290–1294. [Google Scholar] [CrossRef] [Green Version]
- Amirshahrokhi, K.; Bohlooli, S.; Chinifroush, M. The effect of methylsulfonylmethane on the experimental colitis in the rat. Toxicol. Appl. Pharmacol. 2011, 253, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Dini, I.; Laneri, S. Nutricosmetics: A brief overview. Phytother. Res. 2019, 33, 3054–3063. [Google Scholar] [CrossRef]
- Brancaccio, M.; D’Argenio, G.; Lembo, V.; Palumbo, A.; Castellano, I. Antifibrotic Effect of Marine Ovothiol in an in Vivo Model of Liver Fibrosis. Oxidative Med. Cell. Longev. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Brancaccio, M.; Russo, M.; Masullo, M.; Palumbo, A.; Russo, G.L.; Castellano, I. Sulfur-containing histidine compounds inhibit γ-glutamyl transpeptidase activity in human cancer cells. J. Biol. Chem. 2019, 294, 14603–14614. [Google Scholar] [CrossRef]
- D’Argenio, G.; Amoruso, D.C.; Mazzone, G.; Vitaglione, P.; Romano, A.; Ribecco, M.T.; D’Armiento, M.R.; Mezza, E.; Morisco, F.; Fogliano, V.; et al. Garlic extract prevents CCl4-induced liver fibrosis in rats: The role of tissue transglutaminase. Dig. Liver Dis. 2010, 42, 571–577. [Google Scholar] [CrossRef] [PubMed]
- D’Argenio, G.; Mazzone, G.; Ribecco, M.T.; Lembo, V.; Vitaglione, P.; Guarino, M.; Morisco, F.; Napolitano, M.; Fogliano, V.; Caporaso, N. Garlic extract attenuating rat liver fibrosis by inhibiting TGF-β1. Clin. Nutr. 2013, 32, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Milito, A.; Brancaccio, M.; D’Argenio, G.; Castellano, I. Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis. Cells 2019, 8, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milito, A.; Brancaccio, M.; Lisurek, M.; Masullo, M.; Palumbo, A.; Castellano, I. Probing the Interactions of Sulfur-Containing Histidine Compounds with Human Gamma-Glutamyl Transpeptidase. Mar. Drugs 2019, 17, 650. [Google Scholar] [CrossRef] [Green Version]
- Sen, C.K.; Packer, L. Thiol homeostasis and supplements in physical exercise. Am. J. Clin. Nutr. 2000, 72, 653S–669S. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Supplement | Method of Administration | Type of Athletes | Beneficial Action | References |
|---|---|---|---|---|
| GSH | 2 week oral GSH supplementation (1 g/day) | cyclists | reduction in muscle fatigue | [72] |
| ALA | 1200 mg daily for 10 days before exercise | males subjected to physical exertion caused by intense running | reduces RNS/ROS formation, improves the release of EPO and reduces muscle damage | [76] |
| Taurina | 3 g/day of taurine for 8 weeks | male triathletes | reduces RNS/ROS formation | [80] |
| Garlic | 900-mg dose of dried garlic single administration | college endurance athletes | improving the performance of the athletes | [82] |
| SFN | 25 mg/kg body wt ip | group of Male Wistar rats | preventing muscle damage | [83] |
| MSM | 3.0 mg/day for 14 days | males subjected to physical exertion | reduction in muscle fatigue | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brancaccio, M.; Mennitti, C.; Cesaro, A.; Fimiani, F.; Moscarella, E.; Caiazza, M.; Gragnano, F.; Ranieri, A.; D’Alicandro, G.; Tinto, N.; et al. Dietary Thiols: A Potential Supporting Strategy against Oxidative Stress in Heart Failure and Muscular Damage during Sports Activity. Int. J. Environ. Res. Public Health 2020, 17, 9424. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17249424
Brancaccio M, Mennitti C, Cesaro A, Fimiani F, Moscarella E, Caiazza M, Gragnano F, Ranieri A, D’Alicandro G, Tinto N, et al. Dietary Thiols: A Potential Supporting Strategy against Oxidative Stress in Heart Failure and Muscular Damage during Sports Activity. International Journal of Environmental Research and Public Health. 2020; 17(24):9424. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17249424
Chicago/Turabian StyleBrancaccio, Mariarita, Cristina Mennitti, Arturo Cesaro, Fabio Fimiani, Elisabetta Moscarella, Martina Caiazza, Felice Gragnano, Annaluisa Ranieri, Giovanni D’Alicandro, Nadia Tinto, and et al. 2020. "Dietary Thiols: A Potential Supporting Strategy against Oxidative Stress in Heart Failure and Muscular Damage during Sports Activity" International Journal of Environmental Research and Public Health 17, no. 24: 9424. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17249424