
Residual Force Enhancement Is Present in Consecutive Post-Stretch Isometric Contractions of the Hamstrings during a Training Simulation

 , and
, and
Abstract
:1. Introduction
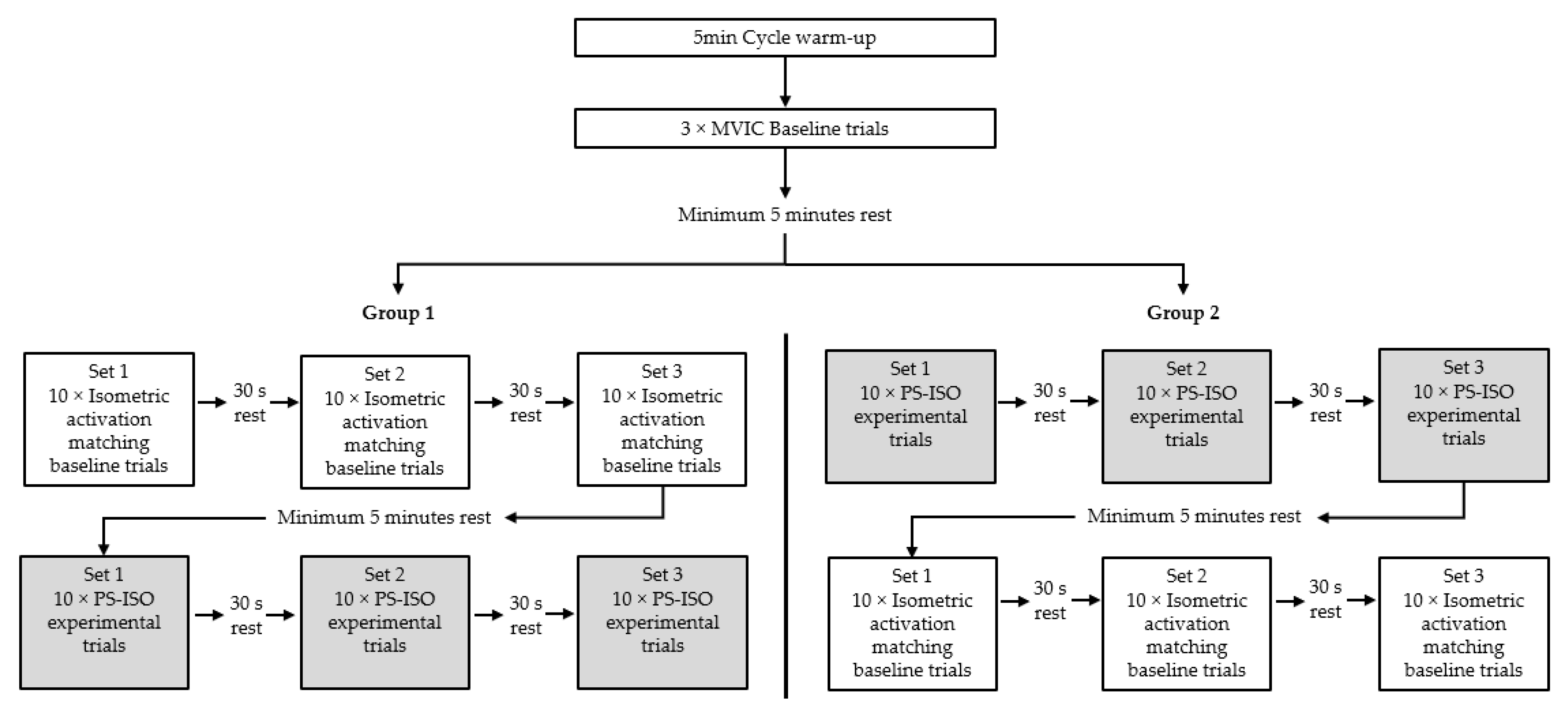
2. Materials and Methods
2.1. Protocols and Measurements
2.2. Data Analysis
2.3. Statistical Analysis
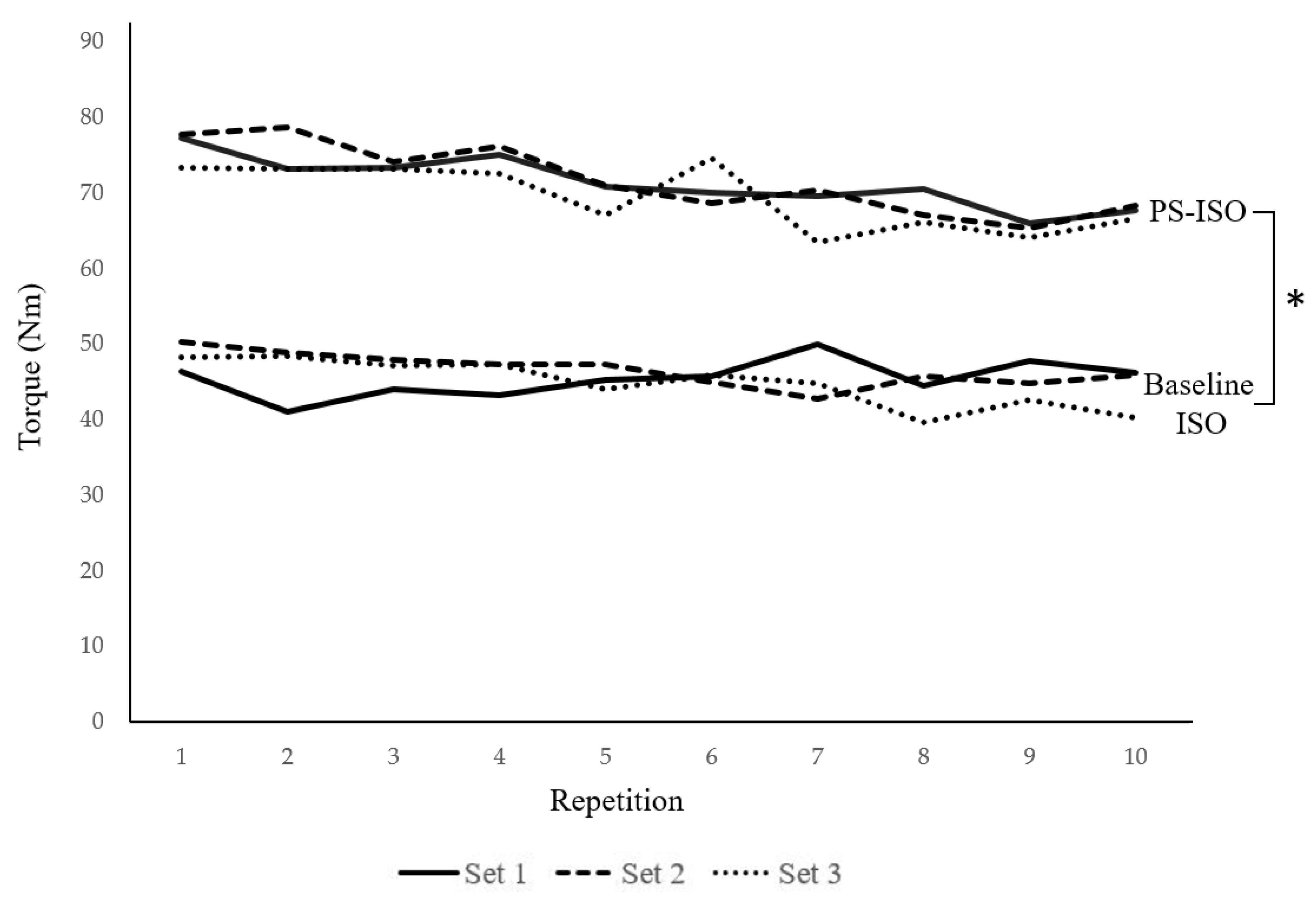
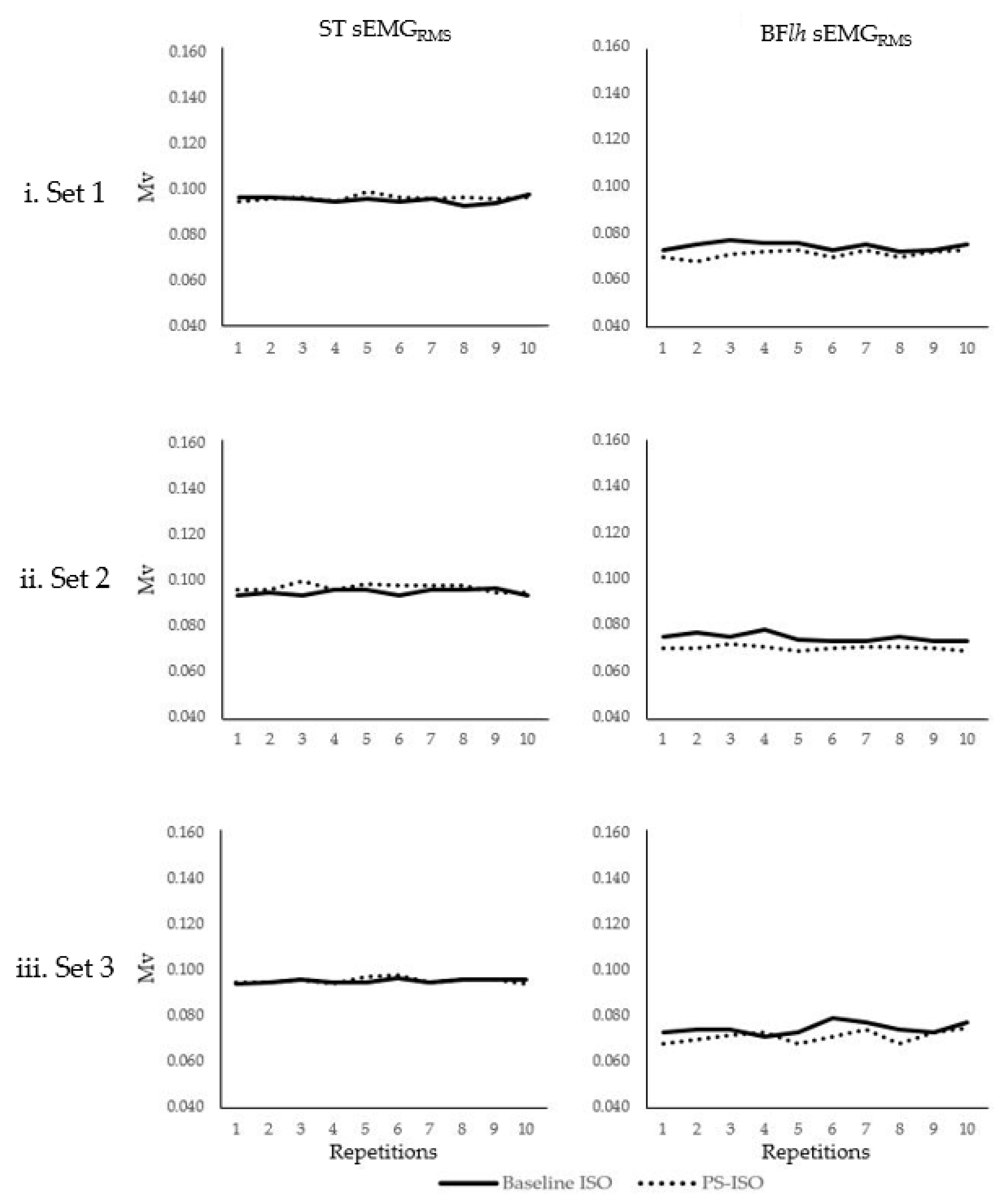
3. Results
3.1. Torque
3.2. sEMGRMS
4. Discussion
- (i)
- (ii)
- The levels of isometric pre-activation in the current study were sufficient to influence muscle lengthening and activation of titin. Previous investigations have suggested that modulation of muscle lengthening is influenced by muscle–tendon interaction [57] and the elimination of muscle slack [58] during the isometric pre-activation phase. The influence of sufficient isometric pre-activation on muscle stretch and magnitude of rFE has been demonstrated in maximal and submaximal PS-ISO contractions [30,50,59].
- (iii)
- A recent investigation of submaximal PS-ISO contractions was undertaken by the current authors, which directly confirmed muscle lengthening of BFlh via ultrasound during PS-ISO contractions [30]. That study used the same body position, joint excursion, angular velocity and submaximal contraction intensity as the current study [30]. We therefore surmise that it is highly likely that muscle lengthening, and therefore engagement of titin, took place in the current study. However, it is acknowledged that other non-contractile elements, such as tendons and aponeuroses, may also have contributed to the enhanced PS-ISO steady-state force.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Green, B.; Bourne, M.N.; Van Dyk, N.; Pizzari, T. Recalibrating the risk of hamstring strain injury (HSI): A 2020 systematic review and meta-analysis of risk factors for index and recurrent hamstring strain injury in sport. Br. J. Sports Med. 2020, 54, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Van Hooren, B.; Frans, B. Is There Really an Eccentric Action of the Hamstrings During the Swing Phase of High-Speed Running? Part I: A Critical Review of the Literature. J. Sports Sci. 2017, 35, 2313–2321. [Google Scholar] [CrossRef] [PubMed]
- Chumanov, E.S.; Heiderscheit, B.C.; Thelen, D.G. Hamstring Musculotendon Dynamics during Stance and Swing Phases of High-Speed Running. Med. Sci. Sports Exerc. 2011, 43, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Higashihara, A.; Nagano, Y.; Ono, T.; Fukubayashi, T. Relationship between the peak time of hamstring stretch and activation during sprinting. Eur. J. Sport Sci. 2016, 16, 36–41. [Google Scholar] [CrossRef]
- Higashihara, A.; Nagano, Y.; Takahashi, K.; Fukubayashi, T. Effects of forward trunk lean on hamstring muscle kinematics during sprinting. J. Sports Sci. 2014, 33, 1366–1375. [Google Scholar] [CrossRef]
- Nagano, Y.; Higashihara, A.; Takahashi, K.; Fukubayashi, T. Mechanics of the muscles crossing the hip joint during sprint running. J. Sports Sci. 2014, 32, 1722–1728. [Google Scholar] [CrossRef]
- Schache, A.G.; Dorn, T.W.; Wrigley, T.V.; Brown, N.A.T.; Pandy, M.G. Stretch and activation of the human biarticular hamstrings across a range of running speeds. Graefe’s Arch. Clin. Exp. Ophthalmol. 2013, 113, 2813–2828. [Google Scholar] [CrossRef]
- Simonsen, E.B.; Thomsen, L.; Klausen, K. Activity of mono- and bi-articular leg muscles during sprint running. Graefe’s Arch. Clin. Exp. Ophthalmol. 1985, 54, 524–532. [Google Scholar]
- Thelen, D.G.; Chumanov, E.S.; Hoerth, D.M.; Best, T.M.; Swanson, S.C.; Li, L.; Young, M.; Heiderscheit, B.C. Hamstring Muscle Kinematics during Treadmill Sprinting. Med. Sci. Sports Exerc. 2005, 37, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Maniar, N.; Schache, A.; Heiderscheit, B.; Opar, D. Hamstrings Biomechanics Related to Running. In Prevention and Rehabilitation of Hamstring Injuries; Springer: Berlin, Germany, 2020; pp. 65–81. [Google Scholar]
- Pizzari, T.; Green, B.; Van Dyk, N. Extrinsic and Intrinsic Risk Factors Associated with Hamstring Injury. In Prevention and Rehabilitation of Hamstring Injuries; Springer: Berlin, Germany, 2020; pp. 83–115. [Google Scholar]
- Askling, C.; Karlsson, J.; Thorstensson, A. Hamstring injury occurrence in elite soccer players after preseason strength training with eccentric overload. Scand. J. Med. Sci. Sports 2003, 13, 244–250. [Google Scholar] [CrossRef]
- Brooks, J.H.M.; Fuller, C.W.; Kemp, S.P.T.; Reddin, D.B. Incidence, Risk, and Prevention of Hamstring Muscle Injuries in Professional Rugby Union. Am. J. Sports Med. 2006, 34, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Askling, C.M.; Tengvar, M.; Tarassova, O.; Thorstensson, A. Acute hamstring injuries in Swedish elite sprinters and jumpers: A prospective randomised controlled clinical trial comparing two rehabilitation protocols. Br. J. Sports Med. 2014, 48, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Askling, C.M.; Tengvar, M.; Thorstensson, A. Acute hamstring injuries in Swedish elite football: A prospective randomised controlled clinical trial comparing two rehabilitation protocols. Br. J. Sports Med. 2013, 47, 953–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrissey, M.C.; Harman, E.A.; Johnson, M.J. Resistance training modes: Specificity and effectiveness. Med. Sci. Sports Exerc. 1995, 27, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Van Hooren, B.; Frans, B. Preventing Hamstring Injuries-Part 2: There Is Possibly an Isometric Action of the Hamstrings in High-Speed Running and It Does Matter. Sport Perf. Sci. Rep. 2018, 1, 1–5. [Google Scholar]
- Van Hooren, B.; Frans, B. Is There Really an Eccentric Action of the Hamstrings During the Swing Phase of High-Speed Running? Part II: Implications for Exercise. J. Sports Sci. 2017, 35, 2322–2333. [Google Scholar] [CrossRef]
- Chapman, N.; Whitting, J.; Broadbent, S.; Crowley-McHattan, Z.; Meir, R.A. Residual Force Enhancement in Humans: A Systematic Review. J. Appl. Biomech. 2018, 34, 240–248. [Google Scholar] [CrossRef]
- Edman, K.A.P.; Elzinga, G.; Noble, M.I. Residual force enhancement after stretch of contracting frog single muscle fibers. J. Gen. Physiol. 1982, 80, 769–784. [Google Scholar] [CrossRef]
- Campbell, S.G.; Campbell, K.S. Mechanisms of residual force enhancement in skeletal muscle: Insights from experiments and mathematical models. Biophys. Rev. 2011, 3, 199–207. [Google Scholar] [CrossRef]
- Herzog, W.; Leonard, T. The history dependence of force production in mammalian skeletal muscle following stretch-shortening and shortening-stretch cycles. J. Biomech. 2000, 33, 531–542. [Google Scholar] [CrossRef]
- Peterson, D.R.; Rassier, D.E.; Herzog, W. Force Enhancement in Single Skeletal Muscle Fibres on the Ascending Limb of the Force-Length Relationship. J. Exp. Biol. 2004, 16, 2787–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rassier, D.E.; Herzog, W.; Wakeling, J.; Syme, D.A. Stretch-Induced, Steady-State Force Enhancement in Single Skeletal Muscle Fibers Exceeds the Isometric Force at Optimum Fiber Length. J. Biomech. 2003, 36, 1309–1316. [Google Scholar] [CrossRef]
- Schachar, R.; Herzog, W.; Leonard, T. The effects of muscle stretching and shortening on isometric forces on the descending limb of the force–length relationship. J. Biomech. 2004, 37, 917–926. [Google Scholar] [CrossRef]
- Cook, C.S.; McDonagh, M.J. Force Responses to Controlled Stretches of Electrically Stimulated Human Muscle-Tendon Complex. Exp. Physiol. 1995, 80, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Herzog, W. Force enhancement following muscle stretch of electrically stimulated and voluntarily activated human adductor pollicis. J. Physiol. 2002, 545, 321–330. [Google Scholar] [CrossRef]
- Pinniger, G.J.; Cresswell, A.G. Residual force enhancement after lengthening is present during submaximal plantar flexion and dorsiflexion actions in humans. J. Appl. Physiol. 2007, 102, 18–25. [Google Scholar] [CrossRef] [Green Version]
- De Ruiter, C.J.; Didden, W.J.M.; Jones, D.A.; De Haan, A. The force-velocity relationship of human adductor pollicis muscle during stretch and the effects of fatigue. J. Physiol. 2000, 526, 671–681. [Google Scholar] [CrossRef]
- Chapman, N.; Whitting, J.; Broadbent, S.; Crowley-McHattan, Z.; Meir, R. Maximal and Submaximal Isometric Torque Is Elevated Following Highly Controlled Active Stretches of the Hamstrings. J. Electromyogr. Kinesiol. 2020, 56, 102500. [Google Scholar] [CrossRef]
- Hahn, D.; Seiberl, W.; Schmidt, S.; Schweizer, K.; Schwirtz, A. Evidence of residual force enhancement for multi-joint leg extension. J. Biomech. 2010, 43, 1503–1508. [Google Scholar] [CrossRef]
- Oskouei, A.E.; Herzog, W. Observations on force enhancement in submaximal voluntary contractions of human adductor pollicis muscle. J. Appl. Physiol. 2005, 98, 2087–2095. [Google Scholar] [CrossRef] [Green Version]
- Tilp, M.; Steib, S.; Herzog, W. Force–time history effects in voluntary contractions of human tibialis anterior. Graefe’s Arch. Clin. Exp. Ophthalmol. 2009, 106, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Julian, F.J.; Morgan, D.L. The effect on tension of non-uniform distribution of length changes applied to frog muscle fibres. J. Physiol. 1979, 293, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.L.; Whitehead, N.P.; Wise, A.K.; Gregory, J.E.; Proske, U. Tension changes in the cat soleus muscle following slow stretch or shortening of the contracting muscle. J. Physiol. 2000, 522, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Bullimore, S.R.; Leonard, T.; Rassier, D.E.; Herzog, W. History-dependence of isometric muscle force: Effect of prior stretch or shortening amplitude. J. Biomech. 2007, 40, 1518–1524. [Google Scholar] [CrossRef] [PubMed]
- Edman, K.A.; Elzinga, G.; Noble, M.I. Enhancement of Mechanical Performance by Stretch during Tetanic Contractions of Vertebrate Skeletal Muscle Fibres. J. Physiol. 1978, 281, 139–155. [Google Scholar] [CrossRef] [Green Version]
- Herzog, W.; Leonard, T. The role of passive structures in force enhancement of skeletal muscles following active stretch. J. Biomech. 2005, 38, 409–415. [Google Scholar] [CrossRef]
- De Vos, R.-J.; Reurink, G.; Van Der Made, A.D.; Kerkhoffs, G.M.; Purdam, C.; Thorborg, K. When Hamstring Injury Rehabilitation Fails. In Prevention and Rehabilitation of Hamstring Injuries; Springer: Berlin, Germany, 2020; pp. 315–347. [Google Scholar]
- Brughelli, M.; Cronin, J. Altering the Length-Tension Relationship with Eccentric Exercise: Implications for Performance and Injury. Sports Med. 2007, 37, 807–826. [Google Scholar] [CrossRef]
- Oranchuk, D.J.; Storey, A.G.; Nelson, A.R.; Cronin, J.B. Isometric training and long-term adaptations: Effects of muscle length, intensity, and intent: A systematic review. Scand. J. Med. Sci. Sports 2018, 29, 484–503. [Google Scholar] [CrossRef]
- Akagi, R.; Hinks, A.; Power, G.A. Differential changes in muscle architecture and neuromuscular fatigability induced by isometric resistance training at short and long muscle-tendon unit lengths. J. Appl. Physiol. 2020, 129, 173–184. [Google Scholar] [CrossRef]
- Rhea, M.R. Determining the Magnitude of Treatment Effects in Strength Training Research Through the Use of the Effect Size. J. Strength Cond. Res. 2004, 18, 918–920. [Google Scholar]
- Hermens, H.J.; Freriks, B.; Merletti, R.; Stegeman, D.; Blok, J.; Rau, G.; Disselhorst-Klug, C.; Hägg, G. European Recommendations for Surface Electromyography. Roessingh Res. Dev. 1999, 8, 13–54. [Google Scholar]
- Bourne, M.N.; Opar, D.A.; Williams, M.D.; Al Najjar, A.; Shield, A.J. Muscle Activation Patterns in the N Ordic Hamstring Exercise: Impact of Prior Strain Injury. Scand. J. Med. Sci. Sports 2016, 26, 666–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudley, G.A.; Tesch, P.A.; Miller, B.J.; Buchanan, P. Importance of eccentric actions in performance adaptations to resistance training. Aviat. Space Environ. Med. 1991, 62, 543–550. [Google Scholar] [PubMed]
- Dalton, B.H.; Contento, V.S.; Power, G.A. Residual force enhancement during submaximal and maximal effort contractions of the plantar flexors across knee angle. J. Biomech. 2018, 78, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Bakeman, R. Recommended effect size statistics for repeated measures designs. Behav. Res. Methods 2005, 37, 379–384. [Google Scholar] [CrossRef]
- Chen, J.; Power, G.A. Modifiability of the history dependence of force through chronic eccentric and concentric biased resistance training. J. Appl. Physiol. 2019, 126, 647–657. [Google Scholar] [CrossRef]
- Fukutani, A.; Herzog, W. Current Understanding of Residual Force Enhancement: Cross-Bridge Component and Non-Cross-Bridge Component. Int. J. Mol. Sci. 2019, 20, 5479. [Google Scholar] [CrossRef] [Green Version]
- Heidlauf, T.; Klotz, T.; Rode, C.; Altan, E.; Bleiler, C.; Siebert, T.; Röhrle, O. A multi-scale continuum model of skeletal muscle mechanics predicting force enhancement based on actin–titin interaction. Biomech. Model. Mechanobiol. 2016, 15, 1423–1437. [Google Scholar] [CrossRef]
- Nishikawa, K.C. Titin: A Tunable Spring in Active Muscle. Physiology 2020, 35, 209–217. [Google Scholar] [CrossRef]
- Joumaa, V.; Herzog, W. Energy cost of force production is reduced after active stretch in skinned muscle fibres. J. Biomech. 2013, 46, 1135–1139. [Google Scholar] [CrossRef]
- Contento, V.S.; Dalton, B.H.; Power, G.A. The Inhibitory Tendon-Evoked Reflex Is Increased in the Torque-Enhanced State Following Active Lengthening Compared to a Purely Isometric Contraction. Brain Sci. 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, D.; Hoffman, B.W.; Carroll, T.J.; Cresswell, A.G. Cortical and Spinal Excitability during and after Lengthening Contractions of the Human Plantar Flexor Muscles Performed with Maximal Voluntary Effort. PLoS ONE 2012, 7, e49907. [Google Scholar] [CrossRef] [PubMed]
- Sypkes, C.T.; Kozlowski, B.J.; Grant, J.; Bent, L.R.; McNeil, C.J.; Power, G.A. The Influence of Residual Force Enhancement on Spinal and Supraspinal Excitability. PeerJ 2018, 6, e5421. [Google Scholar] [CrossRef] [PubMed]
- Farris, D.J.; Lichtwark, G.A.; Brown, N.A.T.; Cresswell, A.G. The Role of Human Ankle Plantar Flexor Muscle—Tendon Interaction and Architecture in Maximal Vertical Jumping Examined in Vivo. J. Exp. Biol. 2016, 219, 528–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbert, R.D.; Heroux, M.E.; Diong, J.; Bilston, L.E.; Gandevia, S.C.; Lichtwark, G.A. Changes in the Length and Three-Dimensional Orientation of Muscle Fascicles and Aponeuroses with Passive Length Changes in Human Gastrocnemius Muscles. J. Physiol. 2015, 593, 441–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukutani, A.; Shimoho, K.; Isaka, T. Isometric preactivation before active lengthening increases residual force enhancement. Scand. J. Med. Sci. Sports 2019, 29, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Ahtiainen, J.P.; Pakarinen, A.; Alén, M.; Kraemer, W.; Häkkinen, K. Short vs. Long Rest Period Between the Sets in Hypertrophic Resistance Training: Influence on Muscle Strength, Size, and Hormonal Adaptations in Trained Men. J. Strength Cond. Res. 2005, 19, 572–582. [Google Scholar] [CrossRef]
- Timmins, R.G.; Bourne, M.N.; Shield, A.J.; Williams, M.D.; Lorenzen, C.; Opar, D.A. Short biceps femoris fascicles and eccentric knee flexor weakness increase the risk of hamstring injury in elite football (soccer): A prospective cohort study. Br. J. Sports Med. 2015, 50, 1524–1535. [Google Scholar] [CrossRef]
- Van der Horst, N.; Thorborg, K.; Opar, D. Hamstring Injury Prevention and Implementation. In Prevention and Rehabilitation of Hamstring Injuries; Springer: Berlin, Germany, 2020; pp. 145–163. [Google Scholar]
- Macdonald, B.; O’Neill, J.; Pollock, N.; Van Hooren, B. Single-Leg Roman Chair Hold Is More Effective Than the Nordic Hamstring Curl in Improving Hamstring Strength-Endurance in Gaelic Footballers with Previous Hamstring Injury. J. Strength Cond. Res. 2019, 33, 3302–3308. [Google Scholar] [CrossRef]
- Seiberl, W.; Hahn, D.; Herzog, W.; Schwirtz, A. Feedback controlled force enhancement and activation reduction of voluntarily activated quadriceps femoris during sub-maximal muscle action. J. Electromyogr. Kinesiol. 2012, 22, 117–123. [Google Scholar] [CrossRef]
- Brockett, C.L.; Morgan, D.L.; Proske, U. Human hamstring muscles adapt to eccentric exercise by changing optimum length. Med. Sci. Sports Exerc. 2001, 33, 783–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmins, R.G.; Ruddy, J.D.; Presland, J.; Maniar, N.; Williams, M. Architectural Changes of the Biceps Femoris Long Head after Concentric or Eccentric Training. Med. Sci. Sports Exerc. 2016, 48, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinks, A.; Davidson, B.; Akagi, R.; Power, G.A. Influence of isometric training at short and long muscle-tendon unit lengths on the history dependence of force. Scand. J. Med. Sci. Sports 2021, 31, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Siebert, T.; Kurch, D.; Blickhan, R.; Stutzig, N. Does Weightlifting Increase Residual Force Enhancement? J. Biomech. 2016, 49, 2047–2052. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Mashouri, P.; Fontyn, S.; Valvano, M.; Elliott-Mohamed, S.; Noonan, A.M.; Brown, S.H.M.; Power, G.A. The influence of training-induced sarcomerogenesis on the history dependence of force. J. Exp. Biol. 2020, 223, 218776. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Set 1 | Set 2 | Set 3 | |||||
|---|---|---|---|---|---|---|---|
| Rep | BL | PS-ISO | BL | PS-ISO | BL | PS-ISO | |
| 1 | 46.43 (27.00) | 77.29 (21.97) * | 50.29 (24.57) | 77.86 (24.03) * | 48.34 (20.11) | 73.40 (19.08) * | |
| 2 | 41.18 (24.47) | 73.20 (21.01) * | 48.92 (22.19) | 78.81 (18.50) * | 48.42 (17.76) | 73.29 (17.78) * | |
| 3 | 44.09 (27.79) | 73.35 (26.20) * | 48.07 (26.55) | 74.22 (20.11) * | 47.15 (17.77) | 73.27 (19.16) * | |
| 4 | 43.29 (30.09) | 75.12 (23.98) * | 47.43 (23.02) | 76.22 (22.65) * | 47.43 (20.40) | 72.59 (17.60) * | |
| 5 | 45.28 (24.11) | 70.84 (27.35) * | 47.45 (23.19) | 71.01 (20.04) * | 44.06 (18.74) | 67.19 (19.34) * | |
| 6 | 45.81 (28.76) | 70.14 (24.25) * | 45.02 (21.67) | 68.74 (20.98) * | 46.01 (17.07) | 74.62 (15.12) * | |
| 7 | 50.11 (30.22) | 69.63 (23.06) * | 42.83 (23.85) | 70.43 (19.59) * | 44.85 (18.98) | 63.48 (17.55) * | |
| 8 | 44.63 (25.91) | 70.53 (21.78) * | 45.84 (21.14) | 67.13 (18.81) * | 39.74 (22.10) | 66.23 (15.38) * | |
| 9 | 47.90 (30.78) | 66.01 (20.04) * | 44.91 (17.86) | 65.38 (20.58) * | 42.63 (18.06) | 64.23 (16.61) * | |
| 10 | 46.28 (27.02) | 67.72 (20.76) * | 45.94 (20.90) | 68.34 (21.43) * | 40.25 (18.69) | 66.66 (15.98) * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chapman, N.D.; Whitting, J.W.; Broadbent, S.; Crowley-McHattan, Z.J.; Meir, R. Residual Force Enhancement Is Present in Consecutive Post-Stretch Isometric Contractions of the Hamstrings during a Training Simulation. Int. J. Environ. Res. Public Health 2021, 18, 1154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18031154
Chapman ND, Whitting JW, Broadbent S, Crowley-McHattan ZJ, Meir R. Residual Force Enhancement Is Present in Consecutive Post-Stretch Isometric Contractions of the Hamstrings during a Training Simulation. International Journal of Environmental Research and Public Health. 2021; 18(3):1154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18031154
Chicago/Turabian StyleChapman, Neil D., John W. Whitting, Suzanne Broadbent, Zachary J. Crowley-McHattan, and Rudi Meir. 2021. "Residual Force Enhancement Is Present in Consecutive Post-Stretch Isometric Contractions of the Hamstrings during a Training Simulation" International Journal of Environmental Research and Public Health 18, no. 3: 1154. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18031154