
Development of a Human Estrogen Receptor Dimerization Assay for the Estrogenic Endocrine-Disrupting Chemicals Using Bioluminescence Resonance Energy Transfer

 and
and 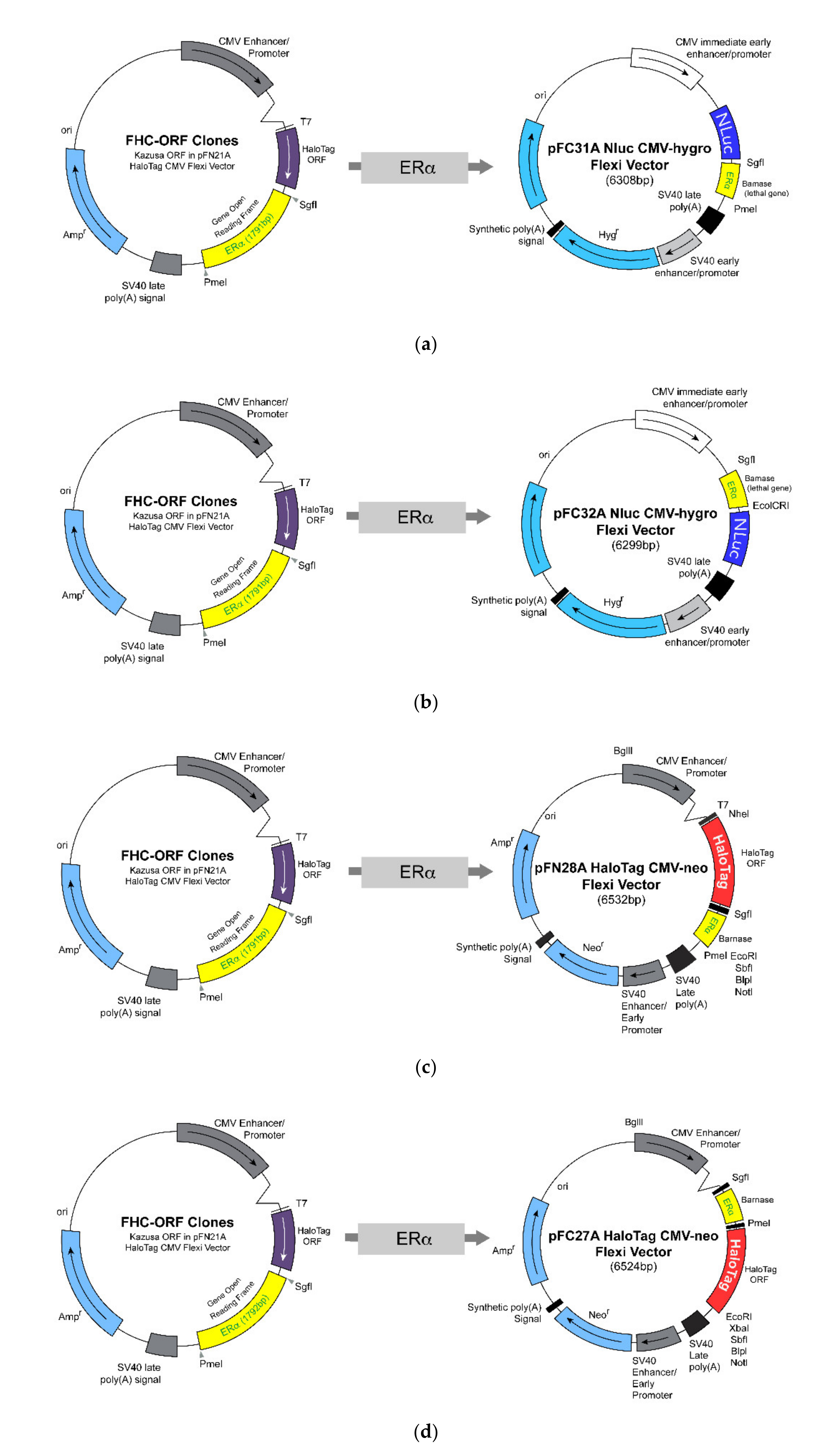{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Construction and Preparation of Vector Containing the Estrogen Receptor-Alpha
2.4. Transient Transfection to HEK293 Cells
2.5. Bioluminescence Resonance Energy Transfer-Based Dimerization Assay
2.6. Data Acquisition and Analyses
3. Results
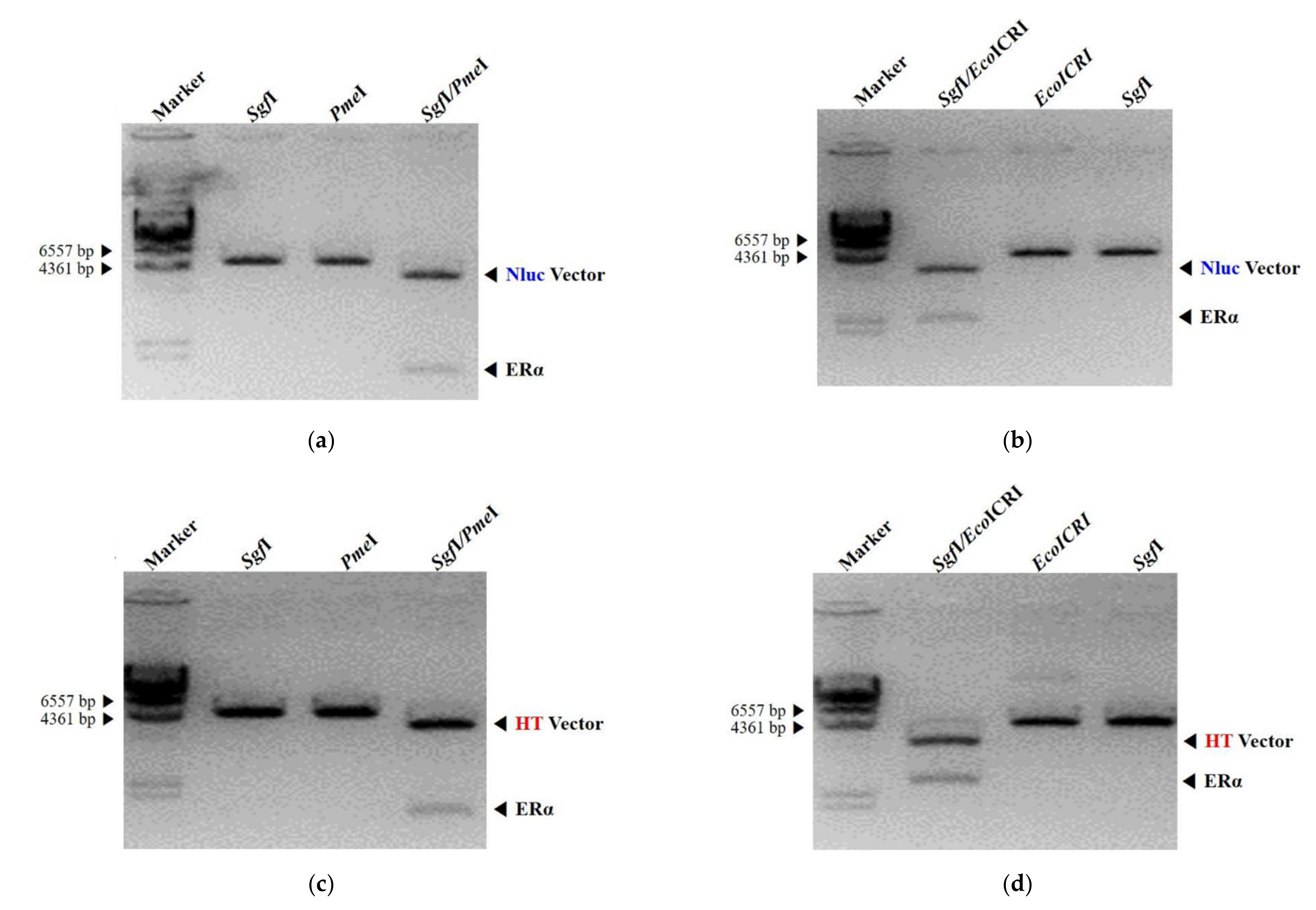
3.1. Design and Construction of NanoLuc Luciferase and HaloTag Fused with Estrogen Receptor-Alpha
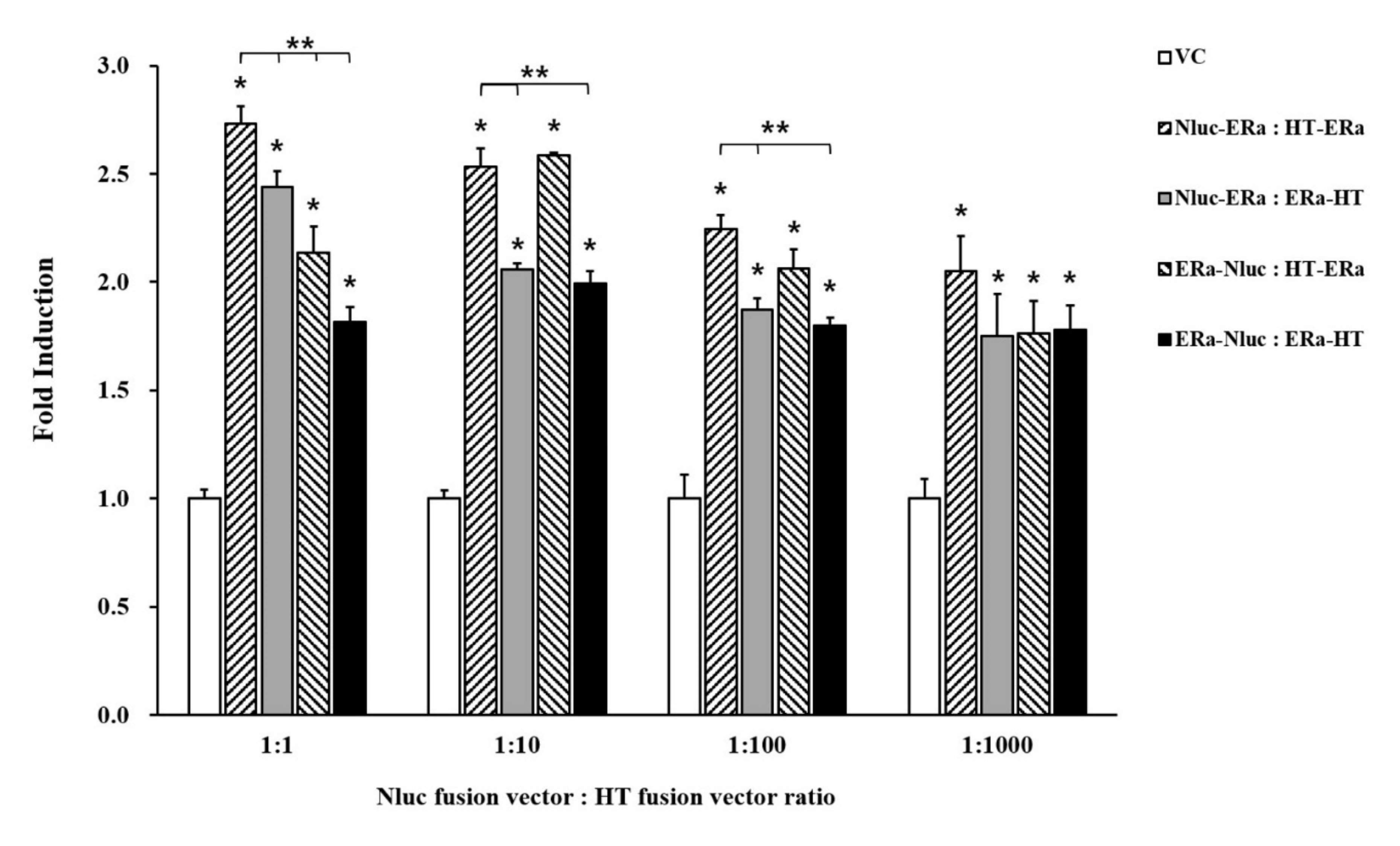
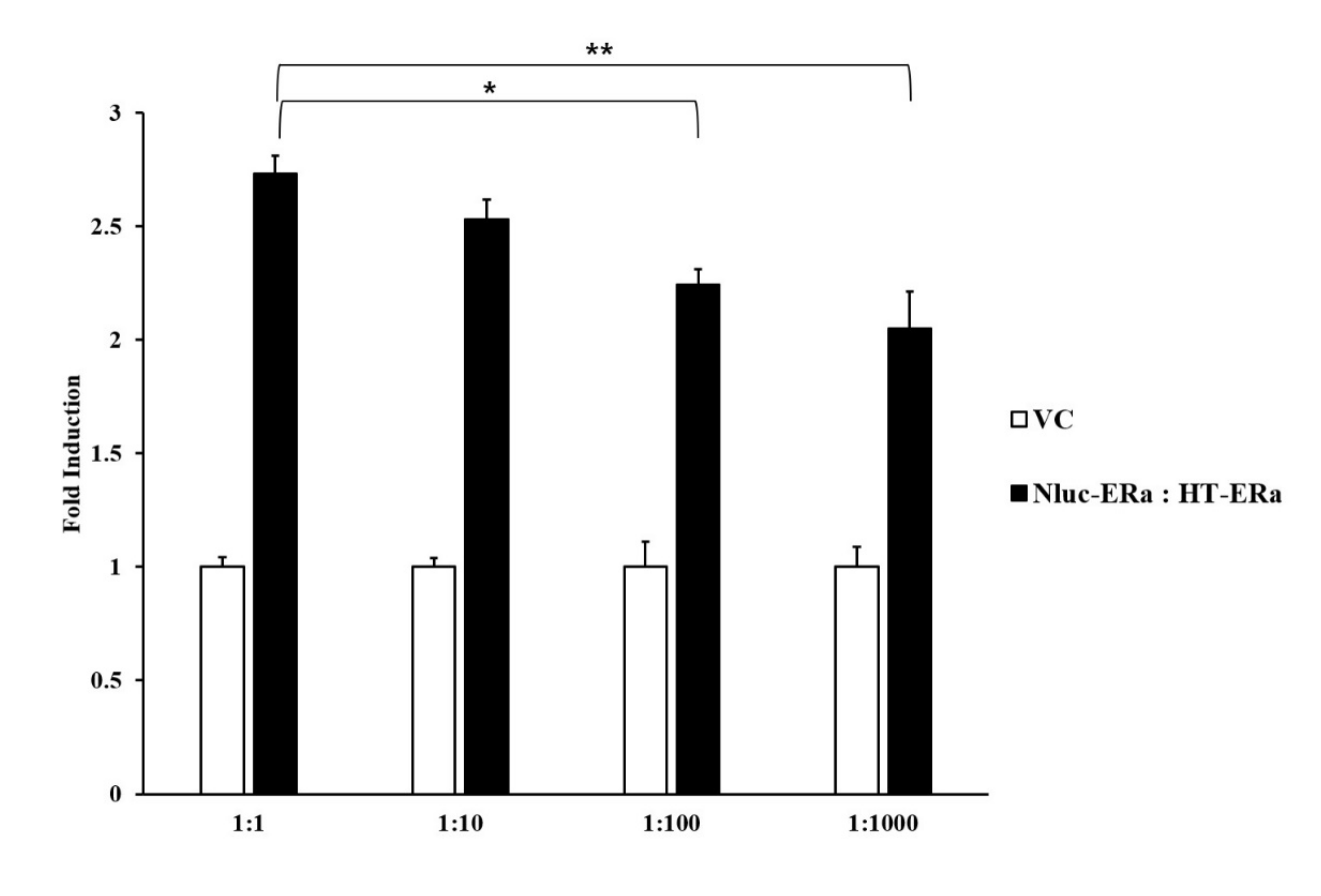
3.2. Bioluminescence Resonance Energy Transfer Signal by Transient Transfection
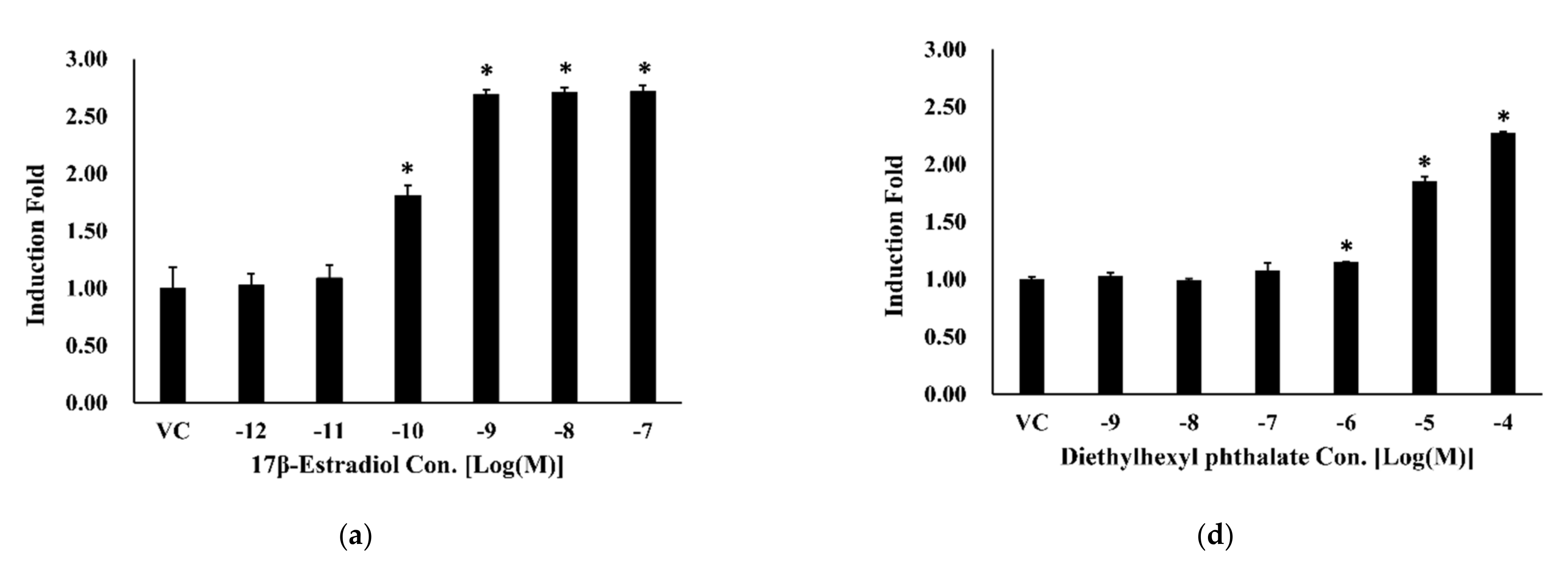
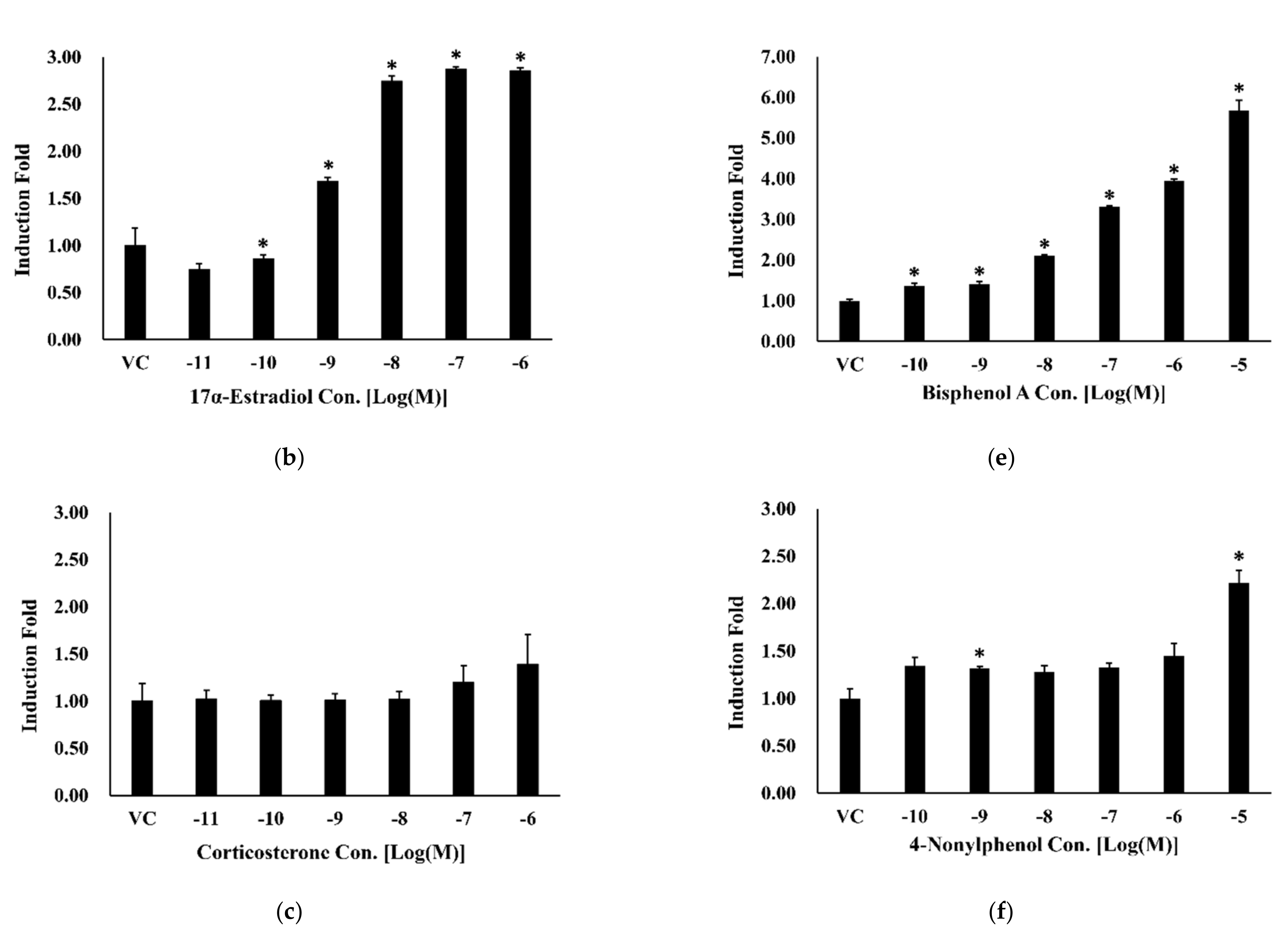
3.3. Bioluminescence Resonance Energy Transfer-Based Estrogen Receptor-Alpha Dimerization Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schug, T.T.; Janesick, A.; Blumberg, B.; Heindel, J.J. Endocrine disrupting chemicals and disease susceptibility. J. Steroid Biochem. Mol. Biol. 2011, 127, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Cotterill, J.V.; Palazzolo, L.; Ridgway, C.; Price, N.; Rorije, E.; Moretto, A.; Peijnenburg, A.; Eberini, I. Predicting estrogen receptor binding of chemicals using a suite of in silico methods—Complementary approaches of (Q)SAR, molecular docking and molecular dynamics. Toxicol. Appl. Pharmacol. 2019, 378, 114630. [Google Scholar] [CrossRef]
- Welshons, W.V.; Thayer, K.A.; Judy, B.M.; Taylor, J.A.; Curran, E.M.; vom Saal, F.S. Large effects from small exposures. I. Mechanisms for endocrine-disrupting chemicals with estrogenic activity. Environ. Health Perspect. 2003, 111, 994–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.H.; Li, N.; Jiang, H.L.; Lin, J.M.; Zhao, R.S. Pretreatment techniques and analytical methods for phenolic endocrine disrupting chemicals in food and environmental samples. TrAC Trends Anal. Chem. 2019, 119, 115592. [Google Scholar] [CrossRef]
- Locatelli, M.; Sciascia, F.; Cifelli, R.; Malatesta, L.; Bruni, P.; Croce, F. Analytical methods for the endocrine disruptor compounds determination in environmental water samples. J. Chromatogr. A 2016, 1434, 1–18. [Google Scholar] [CrossRef]
- Mezcua, M.; Martínez-Uroz, M.A.; Gómez-Ramos, M.M.; Gómez, M.J.; Navas, J.M.; Fernández-Alba, A.R. Analysis of synthetic endocrine-disrupting chemicals in food: A review. Talanta 2012, 100, 90–106. [Google Scholar] [CrossRef]
- Eng, D.S.; Lee, J.M.; Gebremariam, A.; Meeker, J.D.; Peterson, K.; Padmanabhan, V. Bisphenol A and chronic disease risk factors in US children. Pediatrics 2013, 132, e637–e645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, B.S.; Paranjpe, M.; DaFonte, T.; Schaeberle, C.; Soto, A.M.; Obin, M.; Greenberg, A.S. Perinatal BPA exposure alters body weight and composition in a dose specific and sex specific manner: The addition of peripubertal exposure exacerbates adverse effects in female mice. Reprod. Toxicol. 2017, 68, 130–144. [Google Scholar] [CrossRef] [Green Version]
- Colborn, T.; Vom Saal, F.S.; Soto, A.M. Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environ. Health Perspect. 1993, 101, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lam, M.H.W.; Wu, R.S.S.; Jiang, B. Rapid magnetic-mediated solid-phase extraction and pre-concentration of selected endocrine disrupting chemicals in natural waters by poly(divinylbenzene-co-methacrylic acid) coated Fe3O4 core-shell magnetite microspheres for their liquid chromatography-ta. J. Chromatogr. A 2010, 1217, 1219–1226. [Google Scholar] [CrossRef]
- Diel, P.; Smolnikar, K.; Michna, H. In vitro test systems for the evaluation of the estrogenic activity of natural products. Planta Med. 1999, 65, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Kiyama, R.; Wada-Kiyama, Y. Estrogenic endocrine disruptors: Molecular mechanisms of action. Environ. Int. 2015, 83, 11–40. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, H.; Gibson, M.; Li, J. An evaluation on combination effects of phenolic endocrine disruptors by estrogen receptor binding assay. Toxicol. Vitr. 2012, 26, 769–774. [Google Scholar] [CrossRef]
- OECD. OECD Test No. 493: Performance-Based Test Guideline for Human Recombinant Estrogen Receptor (hrER) In vitro Assays to Detect Chemicals with ER Binding Affinity. In OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing: Paris, France, 2015; ISBN 9789264242623. [Google Scholar]
- De Jong, L.A.A.; Uges, D.R.A.; Franke, J.P.; Bischoff, R. Receptor-ligand binding assays: Technologies and applications. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 829, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Han, T.; Deng, A.; Li, L.; Zhu, J.J. Resonance energy transfer in electrochemiluminescent and photoelectrochemical bioanalysis. TrAC Trends Anal. Chem. 2020, 123, 115745. [Google Scholar] [CrossRef]
- Wang, C.; Bian, W.; Xia, C.; Zhang, T.; Guillemot, F.; Jing, N. Visualization of bHLH transcription factor interactions in living mammalian cell nuclei and developing chicken neural tube by FRET. Cell Res. 2006, 16, 585–598. [Google Scholar] [CrossRef]
- Piston, D.W.; Kremers, G.J. Fluorescent protein FRET: The good, the bad and the ugly. Trends Biochem. Sci. 2007, 32, 407–414. [Google Scholar] [CrossRef]
- Michelini, E.; Mirasoli, M.; Karp, M.; Virta, M.; Roda, A. Development of a bioluminescene resonance energy-transfer assay for estrogen-like compound in vivo monitoring. Anal. Chem. 2005, 76, 7069–7076. [Google Scholar] [CrossRef]
- Xu, Y.; Piston, D.W.; Johnson, C.H. A bioluminescence resonance energy transfer (BRET) system: Application to interacting circadian clock proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boute, N.; Jockers, R.; Issad, T. The use of resonance energy transfer in high-throughput screening: BRET versus FRET. Trends Pharmacol. Sci. 2002, 23, 351–354. [Google Scholar] [CrossRef]
- ICCVAM. ICCVAM Evaluation of In Vitro Test Methods for Detecting Potential Endocrine Disruptors: Estrogen Receptor and Androgen Receptor Binding and Transcriptional Activation Assays; No. 03-4503; NIH Publication: Bethesda, MD, USA, 2003.
- Yang, F.W.; Li, Y.X.; Ren, F.Z.; Luo, J.; Pang, G.F. Assessment of the endocrine-disrupting effects of organophosphorus pesticide triazophos and its metabolites on endocrine hormones biosynthesis, transport and receptor binding in silico. Food Chem. Toxicol. 2019, 133, 110759. [Google Scholar] [CrossRef]
- Park, M.-A.; Hwang, K.-A.; Choi, K.-C. Diverse animal models to examine potential role(s) and mechanism of endocrine disrupting chemicals on the tumor progression and prevention: Do they have tumorigenic or anti-tumorigenic property? Lab. Anim. Res. 2011, 27, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akash, M.S.H.; Sabir, S.; Rehman, K. Bisphenol A-induced metabolic disorders: From exposure to mechanism of action. Environ. Toxicol. Pharmacol. 2020, 77, 103373. [Google Scholar] [CrossRef]
- Wang, C.; Yang, L.; Wang, S.; Zhang, Z.; Yu, Y.; Wang, M.; Cromie, M.; Gao, W.; Wang, S.-L. The classic EDCs, phthalate esters and organochlorines, in relation to abnormal sperm quality: A systematic review with meta-analysis. Sci. Rep. 2016, 6, 19982. [Google Scholar] [CrossRef]
- Li, Y.; Luh, C.J.; Burns, K.A.; Arao, Y.; Jiang, Z.; Teng, C.T.; Tice, R.R.; Korach, K.S. Endocrine-disrupting chemicals (EDCs): In vitro mechanism of estrogenic activation and differential effects on ER target genes. Environ. Health Perspect. 2013, 121, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Shanle, E.K.; Xu, W. Endocrine disrupting chemicals targeting estrogen receptor signaling: Identification and mechanisms of action. Chem. Res. Toxicol. 2011, 24, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Zeilinger, M.; Pichler, F.; Nics, L.; Wadsak, W.; Spreitzer, H.; Hacker, M.; Mitterhauser, M. New approaches for the reliable in vitro assessment of binding affinity based on high-resolution real-time data acquisition of radioligand-receptor binding kinetics. EJNMMI Res. 2017, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Parag-Sharma, K.; Amelio, A.L.; Cao, Y. A Bioluminescence Resonance Energy Transfer-Based Approach for Determining Antibody-Receptor Occupancy In vivo. Iscience 2019, 15, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issad, T.; Boute, N.; Pernet, K. A homogenous assay to monitor the activity of the insulin receptor using Bioluminescence Resonance Energy Transfer. Biochem. Pharmacol. 2002, 64, 813–817. [Google Scholar] [CrossRef]
- Hall, J.M.; Couse, J.F.; Korach, K.S. The Multifaceted Mechanisms of Estradiol and Estrogen Receptor Signaling. J. Biol. Chem. 2001, 276, 36869–36872. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; MacHleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef]
- Dale, N.C.; Johnstone, E.K.M.; White, C.W.; Pfleger, K.D.G. NanoBRET: The bright future of proximity-based assays. Front. Bioeng. Biotechnol. 2019, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, L.A.; Kilpatrick, L.E.; Hill, S.J. NanoBRET Approaches to Study Ligand Binding to GPCRs and RTKs. Trends Pharmacol. Sci. 2018, 39, 136–147. [Google Scholar] [CrossRef]
- Machleidt, T.; Woodroofe, C.C.; Schwinn, M.K.; Méndez, J.; Robers, M.B.; Zimmerman, K.; Otto, P.; Daniels, D.L.; Kirkland, T.A.; Wood, K.V. NanoBRET-A Novel BRET Platform for the Analysis of Protein-Protein Interactions. ACS Chem. Biol. 2015, 10, 1797–1804. [Google Scholar] [CrossRef]
- Nath, N.; Flemming, R.; Godat, B.; Urh, M. Development of NanoLuc bridging immunoassay for detection of anti-drug antibodies. J. Immunol. Methods 2017, 450, 17–26. [Google Scholar] [CrossRef]
- Soltysik, K.; Czekaj, P. Membrane estrogen receptors—Is it an alternative way of estrogen action? J. Physiol. Pharmacol. 2013, 64, 129–142. [Google Scholar]
- Hall, J.M.; Mcdonnell, D.P. Coregulators in Nuclear Estrogen Receptor Action. Mol. Interv. 2005, 5, 343–357. [Google Scholar] [CrossRef]
- Powell, E.; Xu, W. Intermolecular interactions identify ligand-selective activity of estrogen receptor α/β dimers. Proc. Natl. Acad. Sci. USA 2008, 105, 19012–19017. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.S.; De Fatima, L.A.; Frank, A.P.; Carneiro, E.M.; Clegg, D.J. The effects of 17 alpha-estradiol to inhibit inflammation in vitro. Biol. Sex. Differ. 2017, 8, 30. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.M.; Seo, H.; Park, Y.; Lee, H.-S.; Lee, S.-H.; Ko, K.S. Development of a Human Estrogen Receptor Dimerization Assay for the Estrogenic Endocrine-Disrupting Chemicals Using Bioluminescence Resonance Energy Transfer. Int. J. Environ. Res. Public Health 2021, 18, 8875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18168875
Kim HM, Seo H, Park Y, Lee H-S, Lee S-H, Ko KS. Development of a Human Estrogen Receptor Dimerization Assay for the Estrogenic Endocrine-Disrupting Chemicals Using Bioluminescence Resonance Energy Transfer. International Journal of Environmental Research and Public Health. 2021; 18(16):8875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18168875
Chicago/Turabian StyleKim, Hye Mi, Hyeyeong Seo, Yooheon Park, Hee-Seok Lee, Seok-Hee Lee, and Kwang Suk Ko. 2021. "Development of a Human Estrogen Receptor Dimerization Assay for the Estrogenic Endocrine-Disrupting Chemicals Using Bioluminescence Resonance Energy Transfer" International Journal of Environmental Research and Public Health 18, no. 16: 8875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18168875