
Association of COMT Polymorphisms with Multiple Physical Activity-Related Injuries among University Students in China

 , , ,
, , ,
Abstract
:1. Introduction
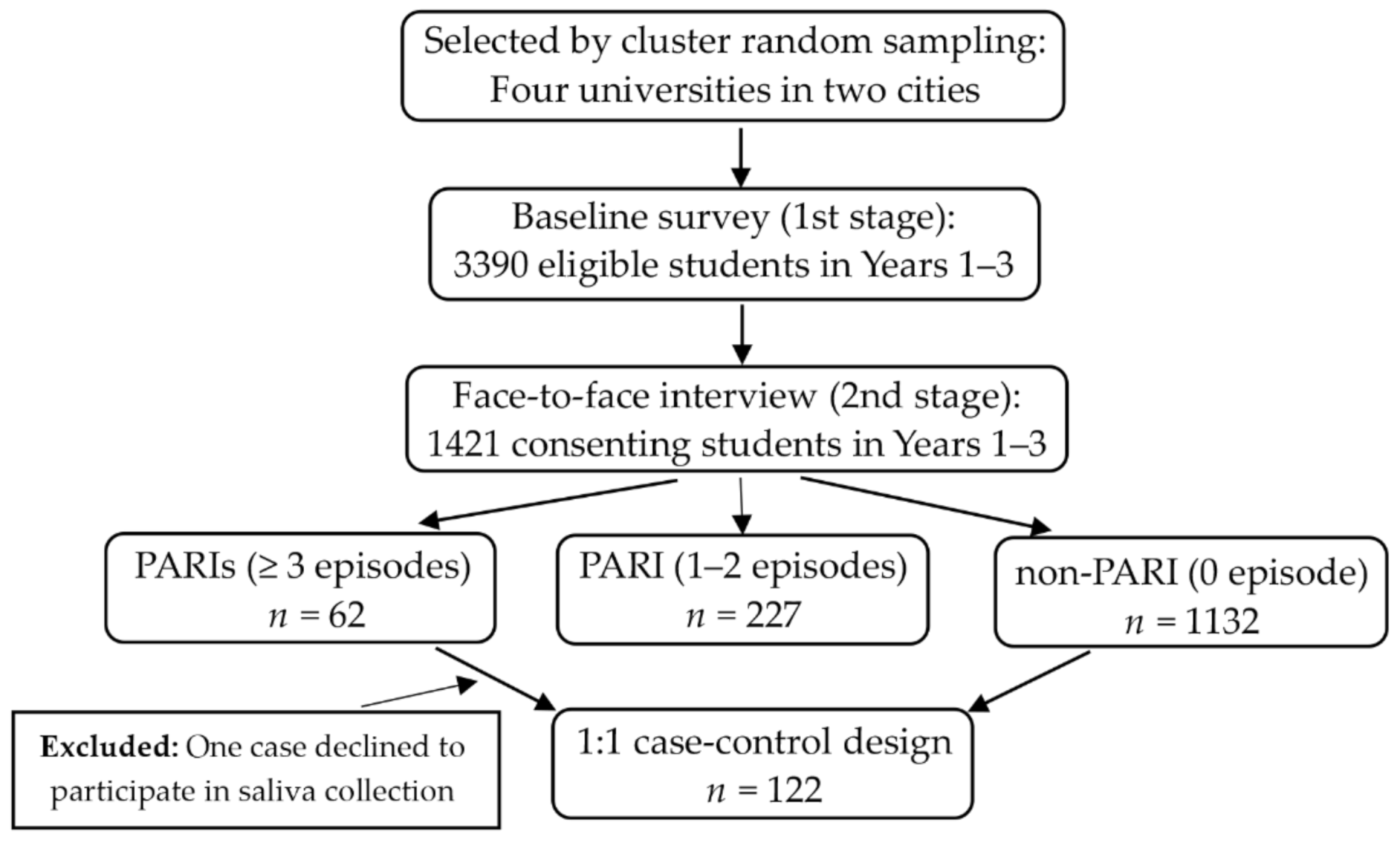
2. Materials and Methods
2.1. Participants
2.2. Data Collection
2.3. SNP Genotyping
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garber, C.E.; Blissmer, B.; Deschenes, M.R.; Franklin, B.A.; Lamonte, M.J.; Lee, I.M.; Nieman, D.C.; Swain, D.P.; The American College of Sports Medicine. American College of Sports Medicine position stand. Quantity and quality of exercise for developing and maintaining cardiorespiratory, musculoskeletal, and neuromotor fitness in apparently healthy adults: Guidance for prescribing exercise. Med. Sci. Sports Exerc. 2011, 43, 1334–1359. [Google Scholar] [CrossRef]
- 2018 Physical Activity Guidelines Advisory Committee. 2018 Physical Activity Guidelines Advisory Committee Scientific Report; U.S. Department of Health and Human Services: Washington, DC, USA, 2018.
- Nielsen, R.O.; Bertelsen, M.L.; Ramskov, D.; Moller, M.; Hulme, A.; Theisen, D.; Finch, C.F.; Fortington, L.V.; Mansournia, M.A.; Parner, E.T. Time-to-event analysis for sports injury research part 2: Time-varying outcomes. Br. J. Sports Med. 2019, 53, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Anson, J.; Waddington, G.; Adams, R.; Liu, Y. The Role of Ankle Proprioception for Balance Control in relation to Sports Performance and Injury. Biomed. Res. Int. 2015, 2015, 842804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logerstedt, D.; Arundale, A.; Lynch, A.; Snyder-Mackler, L. A conceptual framework for a sports knee injury performance profile (SKIPP) and return to activity criteria (RTAC). Braz. J. Phys. Ther. 2015, 19, 340–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weggemans, R.M.; Backx, F.J.G.; Borghouts, L.; Chinapaw, M.; Hopman, M.T.E.; Koster, A.; Kremers, S.; van Loon, L.J.C.; May, A.; Mosterd, A.; et al. The 2017 Dutch Physical Activity Guidelines. Int. J. Behav. Nutr. Phys. Act. 2018, 15, 58. [Google Scholar] [CrossRef]
- Verhagen, E.; Collard, D.; Paw, M.C.; van Mechelen, W. A prospective cohort study on physical activity and sports-related injuries in 10-12-year-old children. Br. J. Sports Med. 2009, 43, 1031–1035. [Google Scholar] [CrossRef] [Green Version]
- Bloemers, F.; Collard, D.; Mai, C.A.P.; Mechelen, W.V.; Twisk, J.; Verhagen, E. Physical inactivity is a risk factor for physical activity-related injuries in children. Br. J. Sports Med. 2012, 46, 669–674. [Google Scholar] [CrossRef]
- Zuckerman, S.L.; Wegner, A.M.; Roos, K.G.; Djoko, A.; Dompier, T.P.; Kerr, Z.Y. Injuries sustained in National Collegiate Athletic Association men’s and women’s basketball, 2009/2010-2014/2015. Br. J. Sports Med. 2018, 52, 261–268. [Google Scholar] [CrossRef]
- Roos, K.G.; Wasserman, E.B.; Dalton, S.L.; Gray, A.; Djoko, A.; Dompier, T.P.; Kerr, Z.Y. Epidemiology of 3825 injuries sustained in six seasons of National Collegiate Athletic Association men’s and women’s soccer (2009/2010–2014/2015). Br. J. Sports Med. 2016, 51, 1029–1034. [Google Scholar] [CrossRef]
- Baugh, C.M.; Weintraub, G.S.; Gregory, A.J.; Djoko, A.; Dompier, T.P.; Kerr, Z.Y. Descriptive Epidemiology of Injuries Sustained in National Collegiate Athletic Association Men’s and Women’s Volleyball, 2013–2014 to 2014–2015. Sports Health 2018, 10, 60–69. [Google Scholar] [CrossRef]
- Gao, Y.; Cai, W.; Gao, L.; Wang, J.; Liang, J.; Kwok, H.; Jia, C.; Li, L. Physical activity-related injuries among university students: A multicentre cross-sectional study in China. BMJ Open 2018, 8, e021845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collard, D.C.; Verhagen, E.A.; van Mechelen, W.; Heymans, M.W.; Chinapaw, M.J. Economic burden of physical activity-related injuries in Dutch children aged 10-12. Br. J. Sports Med. 2011, 45, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Finch, C.; Cassell, E. The public health impact of injury during sport and active recreation. J. Sci. Med. Sport 2006, 9, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Abernethy, L.; Bleakley, C. Strategies to prevent injury in adolescent sport: A systematic review. Br. J. Sports Med. 2007, 41, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Goossens, L.; Verrelst, R.; Cardon, G.; De Clercq, D. Sports injuries in physical education teacher education students. Scand. J. Med. Sci. Sports 2014, 24, 683–691. [Google Scholar] [CrossRef]
- Cai, W.; Gao, Y.; Yang, W.; Cheng, F.; Tang, D.; Li, L. Physical Activity-Related Injury and Its Associated Factors among Middle School Students in Southern China. Int. J. Environ. Res. Public Health 2018, 15, 1244. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Chen, S.; Li, L.; Yue, P.; Yu, X.; Gao, L.; Yang, W.; Jia, C.; Gao, Y. Gender-specific physical activity-related injuries and risk factors among university students in China: A multicentre population-based cross-sectional study. BMJ Open 2020, 10, e040865. [Google Scholar] [CrossRef]
- Pickett, W.; Molcho, M.; Simpson, K.; Janssen, I.; Kuntsche, E.; Mazur, J.; Harel, Y.; Boyce, W.F. Cross national study of injury and social determinants in adolescents. Inj. Prev. 2005, 11, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Wiese-Bjornstal, D.M. Psychology and socioculture affect injury risk, response, and recovery in high-intensity athletes: A consensus statement. Scand. J. Med. Sci. Sports 2010, 20, 103–111. [Google Scholar] [CrossRef]
- Cai, W.; Tang, D.; Gao, Y.; Yang, W.; Chen, S.; Gao, L.; Jia, C.; Li, L. Characteristics and associated factors of physical activity-related injuries among university students in southern China. Sci. Rep. 2020, 10, 4174. [Google Scholar] [CrossRef]
- Perkovic, M.N.; Strac, D.S.; Tudor, L.; Konjevod, M.; Erjavec, G.N.; Pivac, N. Catechol-O-methyltransferase, Cognition and Alzheimer’s Disease. Curr. Alzheimer Res. 2018, 15, 408–419. [Google Scholar] [CrossRef]
- Bastos, P.; Gomes, T.; Ribeiro, L. Catechol-O-Methyltransferase (COMT): An Update on Its Role in Cancer, Neurological and Cardiovascular Diseases. Rev. Physiol. Biochem. Pharmacol. 2017, 173, 1–39. [Google Scholar] [CrossRef]
- Craddock, N.; Owen, M.J.; O’Donovan, M.C. The catechol-O-methyl transferase (COMT) gene as a candidate for psychiatric phenotypes: Evidence and lessons. Mol. Psychiatry 2006, 11, 446–458. [Google Scholar] [CrossRef] [Green Version]
- Crum, A.J.; Akinola, M.; Turnwald, B.P.; Kaptchuk, T.J.; Hall, K.T. Catechol-O-Methyltransferase moderates effect of stress mindset on affect and cognition. PLoS ONE 2018, 13, e0195883. [Google Scholar] [CrossRef] [Green Version]
- Smolka, M.N. Catechol-O-Methyltransferase val158met Genotype Affects Processing of Emotional Stimuli in the Amygdala and Prefrontal Cortex. J. Neurosci. 2005, 25, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Raffaella, C.; Stefano, P.; Ina, G.; Hartmann, A.M.; Hans-Jürgen, M.; Diana, D.R.; Alessandro, S.; Dan, R. Catechol-o-methyltransferase gene modulation on suicidal behavior and personality traits: Review, meta-analysis and association study. J. Psychiatr. Res. 2011, 45, 309–321. [Google Scholar] [CrossRef]
- Harrison, P.J.; Tunbridge, E.M. Catechol-O-Methyltransferase (COMT): A Gene Contributing to Sex Differences in Brain Function, and to Sexual Dimorphism in the Predisposition to Psychiatric Disorders. Neuropsychopharmacology 2008, 33, 3037–3045. [Google Scholar] [CrossRef] [PubMed]
- van Rooij, S.J.; Stevens, J.S.; Ely, T.D.; Fani, N.; Smith, A.K.; Kerley, K.A.; Lori, A.; Ressler, K.J.; Jovanovic, T. Childhood Trauma and COMT Genotype Interact to Increase Hippocampal Activation in Resilient Individuals. Front Psychiatry 2016, 7, 156. [Google Scholar] [CrossRef] [Green Version]
- Haraldsson, H.M.; Ettinger, U.; Magnusdottir, B.B.; Sigmundsson, T.; Sigurdsson, E.; Ingason, A.; Petursson, H. Catechol-O-Methyltransferase Val158Met Polymorphism and Antisaccade Eye Movements in Schizophrenia. Schizophr. Bull. 2010, 36, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Lipska, B.K.; Halim, N.; Ma, Q.D.; Matsumoto, M.; Melhem, S.; Kolachana, B.S.; Hyde, T.M.; Herman, M.M.; Apud, J.; et al. Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): Effects on mRNA, protein, and enzyme activity in postmortem human brain. Am. J. Hum. Genet. 2004, 75, 807–821. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Hinoda, Y.; Okayama, N.; Suehiro, Y.; Kawamoto, K.; Kikuno, N.; Rabban, J.T.; Chen, L.-M.; Dahiya, R. COMT polymorphisms affecting protein expression are risk factors for endometrial cancer. Mol. Carcinog. 2008, 47, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Diatchenko, L.; Slade, G.D.; Nackley, A.G.; Bhalang, K.; Sigurdsson, A.; Belfer, I.; Goldman, D.; Xu, K.; Shabalina, S.A.; Shagin, D.; et al. Genetic basis for individual variations in pain perception and the development of a chronic pain condition. Hum. Mol. Genet. 2005, 14, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nackley, A.G.; Shabalina, S.A.; Tchivileva, I.E.; Satterfield, K.; Korchynskyi, O.; Makarov, S.S.; Maixner, W.; Diatchenko, L. Human catechol-O-methyltransferase haplotypes modulate protein expression by altering mRNA secondary structure. Science 2006, 314, 1930–1933. [Google Scholar] [CrossRef] [Green Version]
- Okochi, T.; Ikeda, M.; Kishi, T.; Kawashima, K.; Kinoshita, Y.; Kitajima, T.; Yamanouchi, Y.; Tomita, M.; Inada, T.; Ozaki, N.; et al. Meta-analysis of association between genetic variants in COMT and schizophrenia: An update. Schizophr. Res. 2009, 110, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lu, R.B.; Yeh, Y.W.; Shih, M.C.; Huang, S.Y. Association study of catechol-O-methyltransferase gene polymorphisms with schizophrenia and psychopathological symptoms in Han Chinese. Genes Brain Behav. 2011, 10, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, D.; Liu, B.; Yao, G.; Fu, Y.; Bi, Z. Catechol-O-methyltransferase (COMT) Val158Met polymorphism and risk of osteoporotic fracture. Mol. Biol. Rep. 2012, 39, 2975–2979. [Google Scholar] [CrossRef]
- Michaelovsky, E.; Gothelf, D.; Korostishevsky, M.; Frisch, A.; Burg, M.; Carmel, M.; Steinberg, T.; Inbar, D.; Apter, A.; Weizman, A. Association between a common haplotype in the COMT gene region and psychiatric disorders in individuals with 22q11.2DS. Int. J. Neuropsychopharmacol. 2008, 11, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Gauderman, W.J. Sample size requirements for matched case-control studies of gene-environment interaction. Stat. Med. 2002, 21, 35–50. [Google Scholar] [CrossRef]
- Yan, L.H.; Chen, P.J.; Zhuang, J. Revision and reliability validity assessment of Children’s Leisure Activities Study Survey. Chin. J. Sch. Health 2011, 32, 268–270. [Google Scholar]
- Taylor, H.L.; Jacobs, D.R., Jr.; Schucker, B.; Knudsen, J.; Leon, A.S.; Debacker, G. A questionnaire for the assessment of leisure time physical activities. J. Chronic Dis. 1978, 31, 741–755. [Google Scholar] [CrossRef]
- Shieh, G. Power and sample size calculations for comparison of two regression lines with heterogeneous variances. PLoS ONE 2018, 13, e0207745. [Google Scholar] [CrossRef]
- Nogueira, N.; Bacelar, M.F.B.; Ferreira, B.P.; Parma, J.O.; Lage, G.M. Association between the catechol-O-methyltransferase (COMT) Val(158)Met polymorphism and motor behavior in healthy adults: A study review. Brain Res. Bull. 2019, 144, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, R.P. Warriors, Worriers, and COVID-19: An Exploratory Study of the Catechol O-Methyltransferase Val158Met Polymorphism Across Populations. Cureus 2020, 12, e10103. [Google Scholar] [CrossRef] [PubMed]
- Roten, L.T.; Fenstad, M.H.; Forsmo, S.; Johnson, M.P.; Moses, E.K.; Austgulen, R.; Skorpen, F. A low COMT activity haplotype is associated with recurrent preeclampsia in a Norwegian population cohort (HUNT2). Mol. Hum. Reprod. 2011, 17, 439–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [Green Version]
- Park, D.J.; Kim, S.H.; Nah, S.S.; Lee, J.H.; Kim, S.K.; Lee, Y.A.; Hong, S.J.; Kim, H.S.; Lee, H.S.; Kim, H.A.; et al. Association between catechol-O-methyl transferase gene polymorphisms and fibromyalgia in a Korean population: A case-control study. Eur. J. Pain 2016, 20, 1131–1139. [Google Scholar] [CrossRef]
- McAllister, T.W. Genetic factors in traumatic brain injury. Handb. Clin. Neurol. 2015, 128, 723–739. [Google Scholar] [CrossRef]
- Hou, T.T.; Lin, F.; Bai, S.; Cleves, M.A.; Xu, H.M.; Lou, X.Y. Generalized multifactor dimensionality reduction approaches to identification of genetic interactions underlying ordinal traits. Genet. Epidemiol. 2019, 43, 24–36. [Google Scholar] [CrossRef] [Green Version]
- Olesen, A.E.; Nielsen, L.M.; Feddersen, S.; Erlenwein, J.; Petzke, F.; Przemeck, M.; Christrup, L.L.; Drewes, A.M. Association Between Genetic Polymorphisms and Pain Sensitivity in Patients with Hip Osteoarthritis. Pain Pract. 2018, 18, 587–596. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Characteristics | Responders (n = 1421) | Non-Responders (n = 1969) | χ2/t | p-Value |
|---|---|---|---|---|
| Gender | 0.938 | 0.333 | ||
| Male | 377 (26.5%) | 552 (28.0) | ||
| Female | 1 044 (73.5%) | 1417 (72.0) | ||
| Grade | 30.89 | 0.001 | ||
| Year 1 | 556 (39.1) | 620 (31.5) | ||
| Year 2 | 447 (31.5) | 605 (30.7) | ||
| Year 3 | 418 (29.4) | 744 (37.8) | ||
| Club member | 3.111 | 0.078 | ||
| Yes | 1065 (74.95) | 1527 (77.56) | ||
| No | 356 (25.05) | 442 (22.44) | ||
| Age (Years) | 20.09 ± 1.15 | 20.04 ± 1.38 | 0.575 | 0.561 |
| PA participation (min/wk) | 691.21 ± 632.64 | 537.90 ± 702.472 | 4.382 | 0.001 |
| PARI | 5.340 | 0.069 | ||
| 0 | 1 132 (79.7) | 1517 (77.1) | ||
| 1–2 | 227 (16.0) | 334 (17.0) | ||
| ≥3 | 62 (4.4) | 118 (5.9) |
| Characteristics | Controls (n = 61) | Cases (n = 61) | χ2/vs | p-Value |
|---|---|---|---|---|
| Gender | 0.000 | 1.000 | ||
| Male | 30 (49.2) | 30 (49.2) | ||
| Female | 31 (50.8) | 31 (50.8) | ||
| Grade | 0.000 | 1.000 | ||
| Year 1 | 28 (45.9) | 28 (45.9) | ||
| Year 2 | 20 (32.8) | 20 (32.8) | ||
| Year 3 | 13 (21.3) | 13 (21.3) | ||
| Club member | 30.500 | 0.001 | ||
| Yes | 10 (16.4) | 40 (65.6) | ||
| No | 51 (83.6) | 21 (34.4) | ||
| Age (Years) | 19.89 ± 1.02 | 20.02 ± 1.23 | −0.641 | 0.523 |
| BMI (kg/m2) | 20.24 ± 2.77 | 21.12 ± 3.30 | −1.593 | 0.114 |
| WHR | 0.78 ± 0.06 | 0.79 ± 0.06 | −0.086 | 0.932 |
| Screen time (hours/day) | 7.16 ± 2.78 | 6.66 ± 2.81 | 0.989 | 0.324 |
| PA participation (min/wk) | 781.38 ± 575.87 | 1070.41 ± 545.52 | −2.846 | 0.005 |
| SNPs | Controls | Cases | χ2, p-Value | OR |
|---|---|---|---|---|
| rs9265 | 0.482, 0.786 | |||
| CC | 17 (28.8) | 16 (27.6) | 1.000 (ref.) | |
| AC | 25 (42.4) | 28 (48.3) | 0.875 (0.327–2.339) | |
| AA | 17 (28.8) | 14 (24.1) | 1.190 (0.499–2.840) | |
| A% | 50.0 | 48.3 | 0.070, 0.792 | 0.933 (0.559–1.558) |
| rs4680 | 3.607, 0.165 | |||
| GG | 32 (56.1) | 23 (39.7) | 1.000 (ref.) | |
| AG | 24 (42.1) | 32 (55.2) | 1.855 (0.874–3.939) | |
| AA | 1 (1.8) | 3 (5.2) | 4.174 (0.408–42.716) | |
| A% | 22.8 | 32.8 | 2.835, 0.092 | 1.649 (0.919–2.959) |
| rs6269 | 6.047, 0.049 | |||
| AA | 20 (33.3) | 30 (50.8) | 1.000 (ref.) | |
| GA | 30 (50.0) | 26 (44.1) | 0.578 (0.267–1.250) | |
| GG | 10 (16.7) | 3 (5.1) | 0.200 (0.049–0.818) | |
| G% | 41.7 | 27.1 | 5.576, 0.018 | 0.521 (0.302–0.898) |
| rs4818 | 5.788, 0.055 | |||
| CC | 20 (32.8) | 30 (51.7) | 1.000 (ref.) | |
| GC | 32 (52.5) | 25 (43.1) | 0.521 (0.241–1.126) | |
| GG | 9 (14.8) | 3 (5.2) | 0.222 (0.054–0.923) | |
| G% | 41.0 | 26.7 | 5.385, 0.020 | 0.525 (0.304–0.908) |
| rs4633 | 1.995, 0.369 | |||
| CC | 39 (63.9) | 33 (54.1) | 1.000 (ref.) | |
| TC | 20 (32.8) | 23 (37.7) | 1.359 (0.637–2.899) | |
| TT | 2 (3.3) | 5 (8.2) | 2.955 (0.538–16.239) | |
| T% | 19.7 | 27.1 | 1.854, 0.173 | 1.514 (0.832–2.756) |
| rs165655 | 0.081, 0.960 | |||
| GG | 17 (27.9) | 18 (30.0) | 1.000 (ref.) | |
| AG | 34 (55.7) | 32 (53.3) | 0.889 (0.291–2.018) | |
| AA | 10 (16.4) | 10 (16.7) | 0.944(0.315–2.834) | |
| A% | 44.3 | 43.3 | 0.021, 0.884 | 0.963 (0.579–1.600) |
| rs165656 | 1.395, 0.498 | |||
| GG | 38 (63.3) | 32 (53.3) | 1.000 (ref.) | |
| CG | 19 (31.7) | 23 (38.3) | 1.438 (0.667–3.099) | |
| CC | 3 (5.0) | 5 (8.3) | 1.979 (0.439–8.929) | |
| C% | 20.8 | 27.5 | 1.455, 0.228 | 1.441 (0.795–2.615) |
| rs165722 | 1.019, 0.601 | |||
| CC | 37 (61.7) | 32 (55.2) | 1.000 (ref.) | |
| TC | 21 (35.0) | 22 (37.9) | 1.211 (0.565–2.957) | |
| TT | 2 (3.3) | 4 (6.9) | 2.312 (0.397–13.469) | |
| T% | 20.8 | 25.9 | 0.835, 0.361 | 1.326 (0.723–2.429) |
| SNPs | OR (95% CI) 1 | OR (95% CI) 2 | OR (95% CI) 3 |
|---|---|---|---|
| rs9265 | |||
| DOM | 1.062 (0.457–2.378) | 0.693 (0.267–1.800) | 0.719 (0.273–1.897) |
| REC | 0.786 (0.345–1.792) | 0.875 (0.332–2.302) | 0.850 (0.297–2.437) |
| rs4680 | |||
| DOM | 1.948 (0.928–4.090) | 1.684 (0.726–3.908) | 1.938 (0.798–4.699) |
| REC | 3.055 (0.308–30.271) | 2.166 (0.168–27.946) | 0.967 (0.043–21.733) |
| rs6269 | |||
| DOM | 0.483 (0.230–1.014) | 0.370 (0.150–0.914) | 0.355 (0.137–0.918) |
| REC | 0.268 (0.070–1.028) | 0.426 (0.096–1.887) | 0.351 (0.075–1.641) |
| rs4818 | |||
| DOM | 0.455 (0.217–0.956) | 0.389 (0.161–0.938) | 0.364 (0.144–0.924) |
| REC | 0.315 (0.081–1.229) | 0.484 (0.107–2.182) | 0.385 (0.082–1.822) |
| rs4633 | |||
| DOM | 1.504 (0.728–3.108) | 1.427 (0.611–3.334) | 1.523 (0.626–3.707) |
| REC | 2.634 (0.491–14.134) | 3.255 (0.522–20.286) | 2.418 (0.324–18.037) |
| rs165655 | |||
| DOM | 0.902 (0.411–1.979) | 0.660 (0.262–1.663) | 0.711 (0.276–1.827) |
| REC | 1.020 (0.391–2.662) | 0.994 (0.314–3.144) | 0.900 (0.262–3.098) |
| rs165656 | |||
| DOM | 1.511 (0.728–3.316) | 1.418 (0.598–3.362) | 1.532 (0.616–3.813) |
| REC | 1.727 (0.394–7.577) | 2.118 (0.411–10.906) | 1.545 (0.237–10.079) |
| rs165722 | |||
| DOM | 1.307 (0.627–2.723) | 1.233 (0.525–2.899) | 1.320 (0.539–3.237) |
| REC | 2.148 (0.378–12.207) | 3.023 (0.465–19.631) | 2.214 (0.291–16.882) |
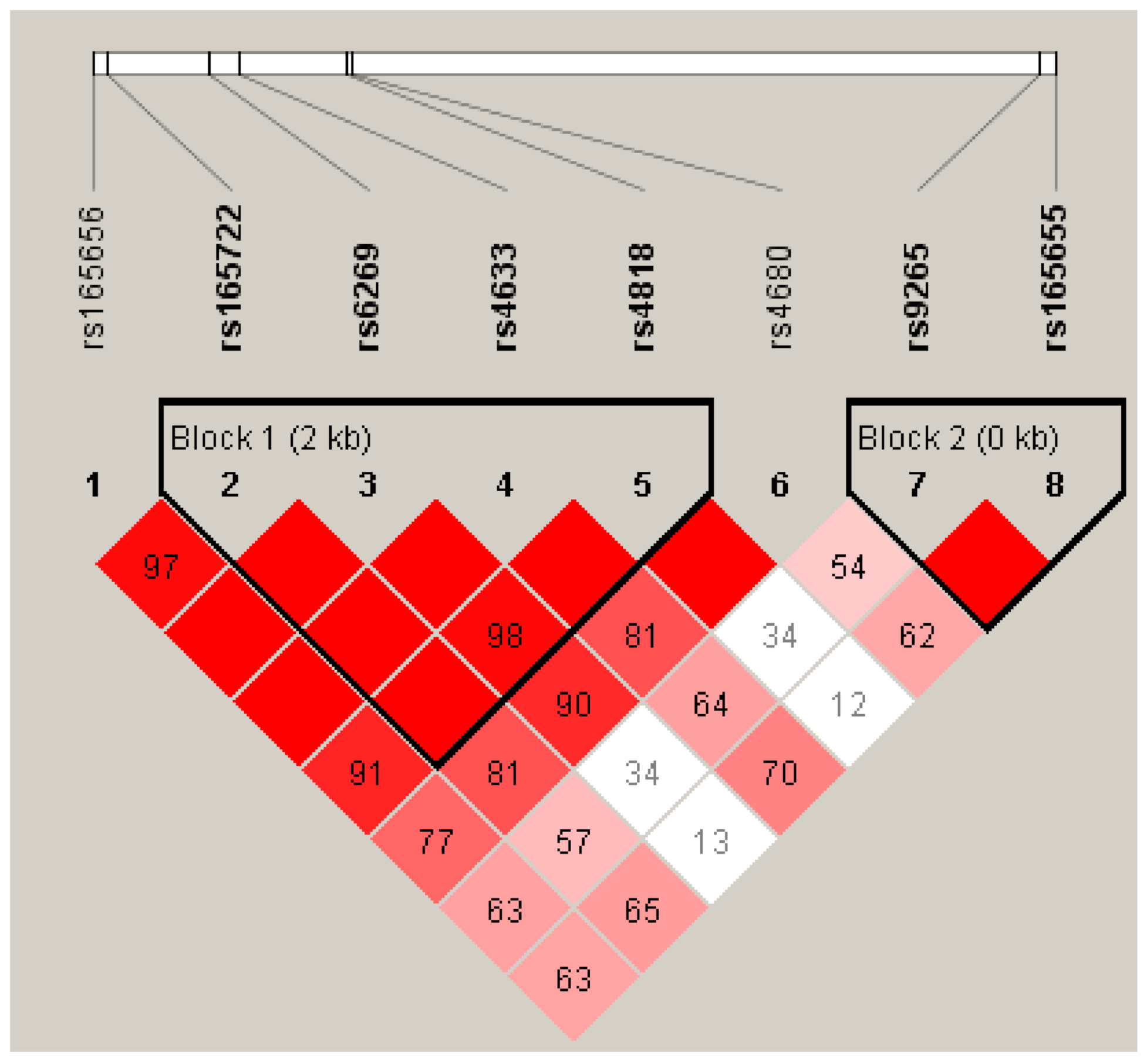
| Block | Haplotype | Controls | Cases | χ2 | p-Value | OR (95% CI) |
|---|---|---|---|---|---|---|
| 1 a | ||||||
| C-A-C-C | 0.385 | 0.452 | 1.137 | 0.286 | 1.490 (0.876–2.533) | |
| C-G-C-G | 0.393 | 0.268 | 4.333 | 0.037 | 0.474 (0.269–0.834) | |
| T-A-T-C | 0.197 | 0.270 | 1.854 | 0.173 | 1.466 (0.786–2.735) | |
| 2 b | ||||||
| C-G | 0.499 | 0.522 | 0.131 | 0.717 | 1.111 (0.664–1.860) | |
| A-A | 0.443 | 0.441 | 0.001 | 0.980 | 0.992 (0.590–1.666) | |
| A-G | 0.059 | 0.037 | 0.621 | 0.431 | 0.577 (0.164–2.026) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Cai, W.; Duan, S.; Gao, L.; Yang, W.; Gao, Y.; Jia, C.; Zhang, H.; Li, L. Association of COMT Polymorphisms with Multiple Physical Activity-Related Injuries among University Students in China. Int. J. Environ. Res. Public Health 2021, 18, 10828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182010828
Chen S, Cai W, Duan S, Gao L, Yang W, Gao Y, Jia C, Zhang H, Li L. Association of COMT Polymorphisms with Multiple Physical Activity-Related Injuries among University Students in China. International Journal of Environmental Research and Public Health. 2021; 18(20):10828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182010828
Chicago/Turabian StyleChen, Shangmin, Weicong Cai, Shiwei Duan, Lijie Gao, Wenda Yang, Yang Gao, Cunxian Jia, Hongjuan Zhang, and Liping Li. 2021. "Association of COMT Polymorphisms with Multiple Physical Activity-Related Injuries among University Students in China" International Journal of Environmental Research and Public Health 18, no. 20: 10828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182010828