Ultraviolet (UV) and Hydrogen Peroxide Activate Ceramide-ER Stress-AMPK Signaling Axis to Promote Retinal Pigment Epithelium (RPE) Cell Apoptosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
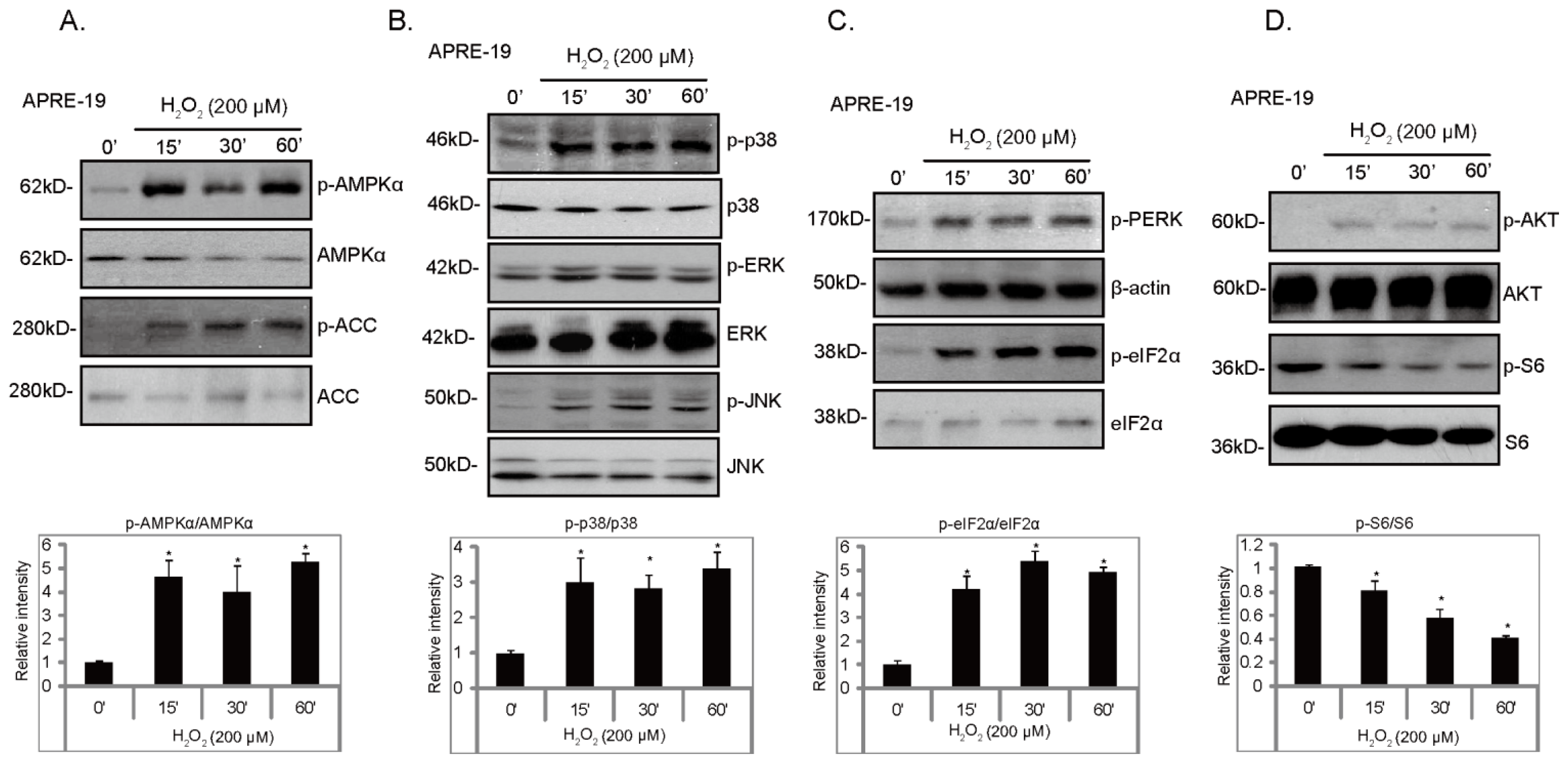
2.1. H2O2 Activates ER Stress, AMPK and MAPK Signal Pathways in Cultured RPE Cells
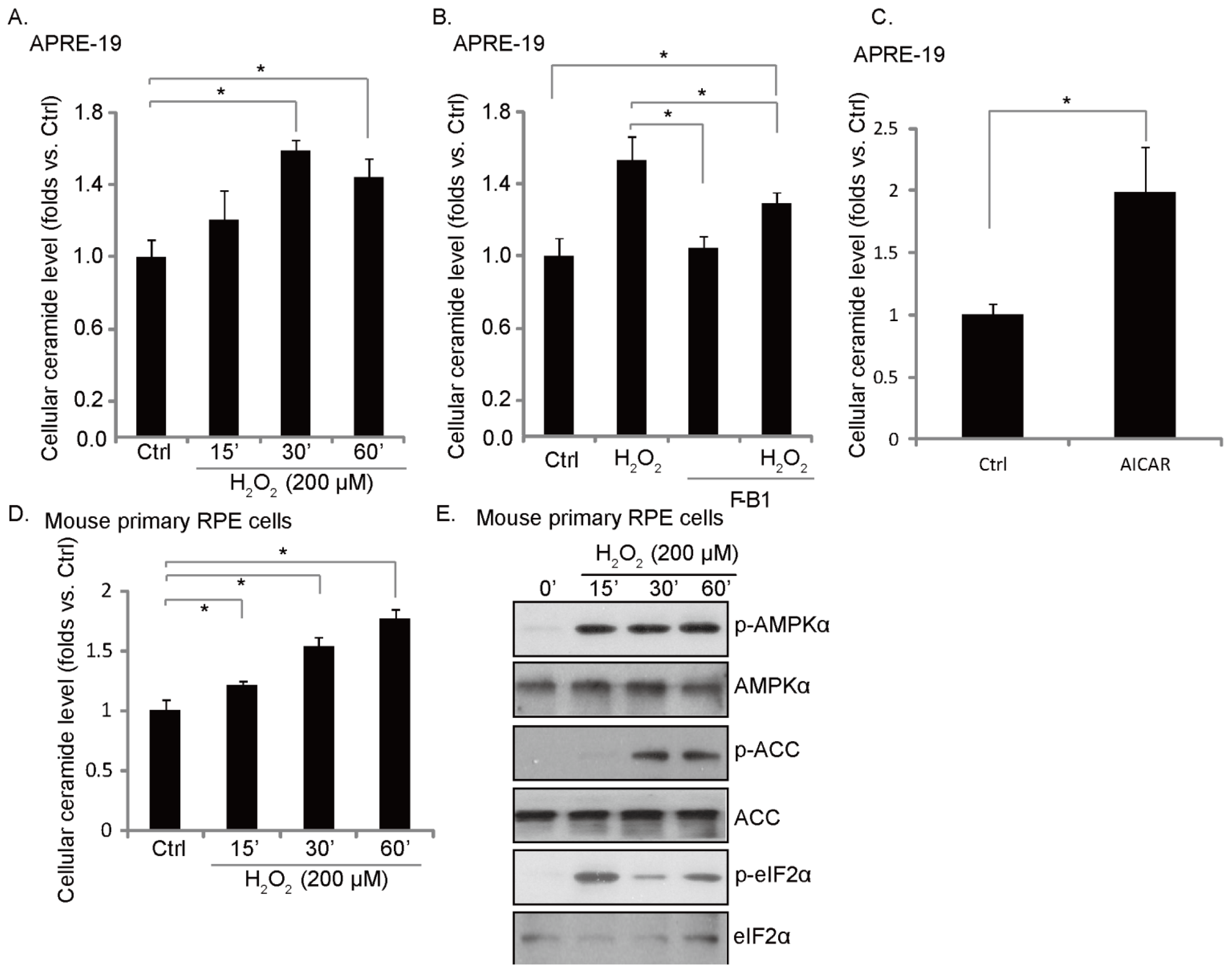
2.2. H2O2 Induces an Early Ceramide Production, Inhibited by Fumonisin B1
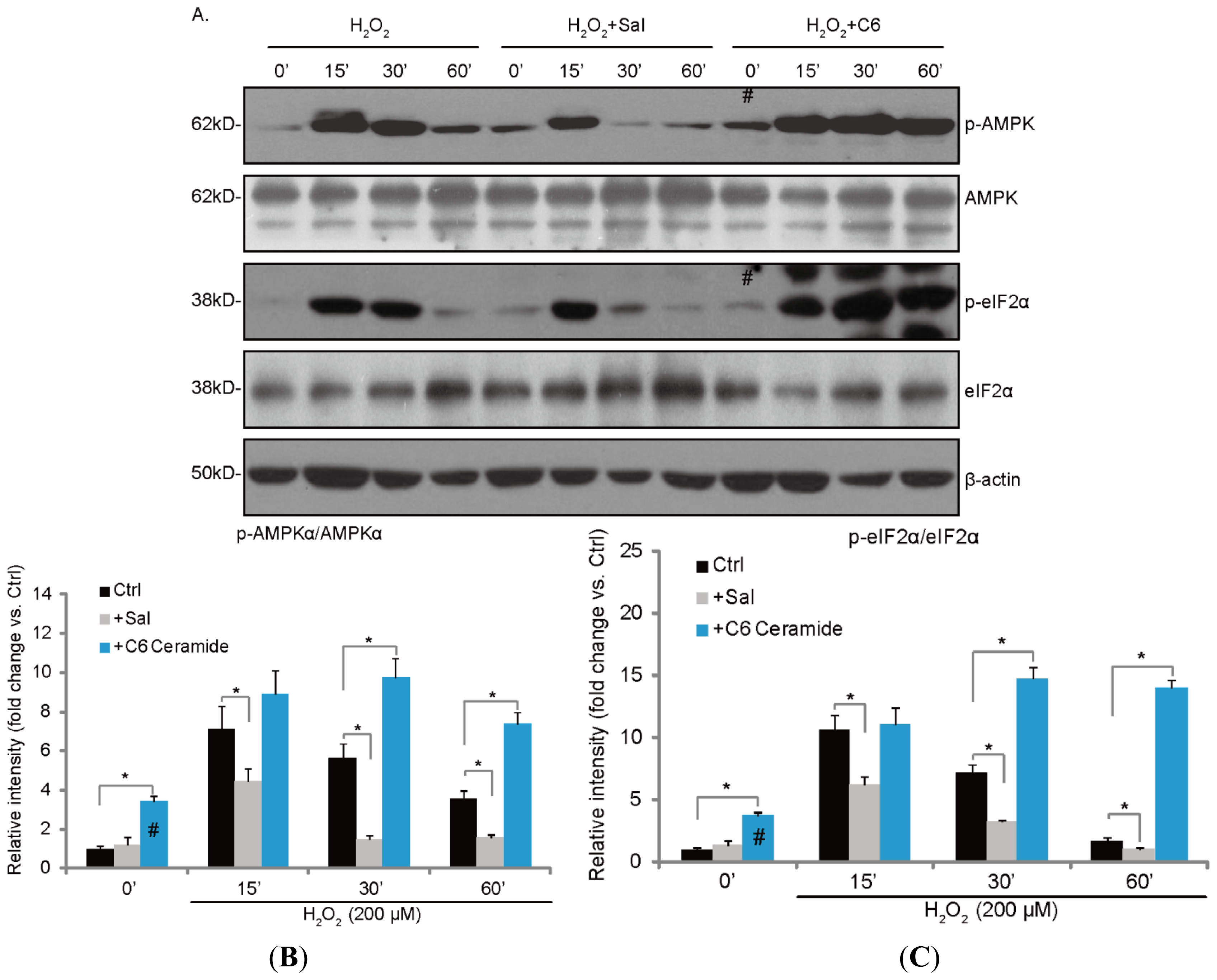
2.3. H2O2-Induced ER Stress and AMPK Activation Is Inhibited by Salubrinal (Sal), but Enhanced by C6 Ceramide
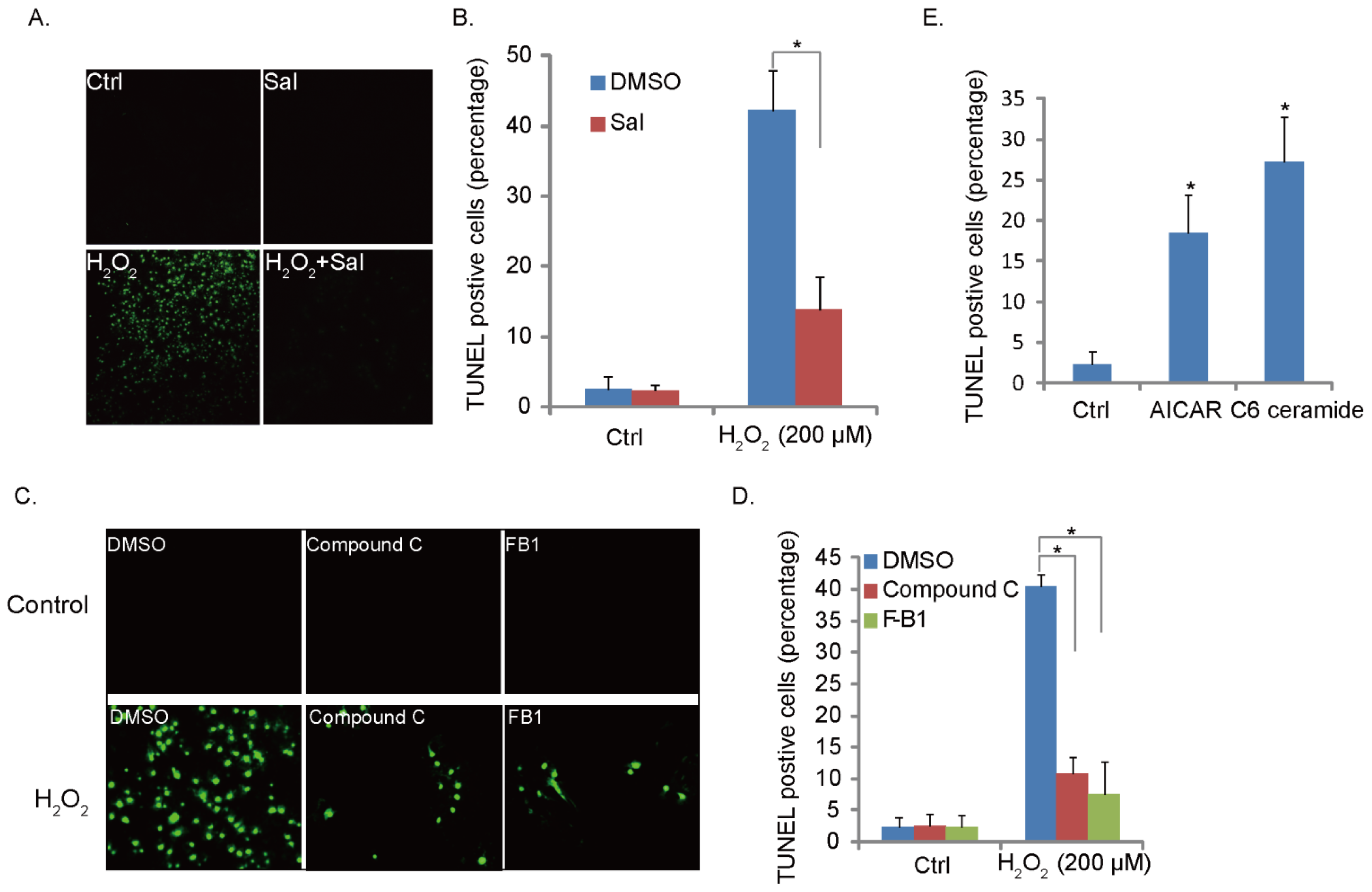
2.4. H2O2-Induced RPE Cell Apoptosis Is Suppressed by Ceramide-ER Stress-AMPK Inhibitors
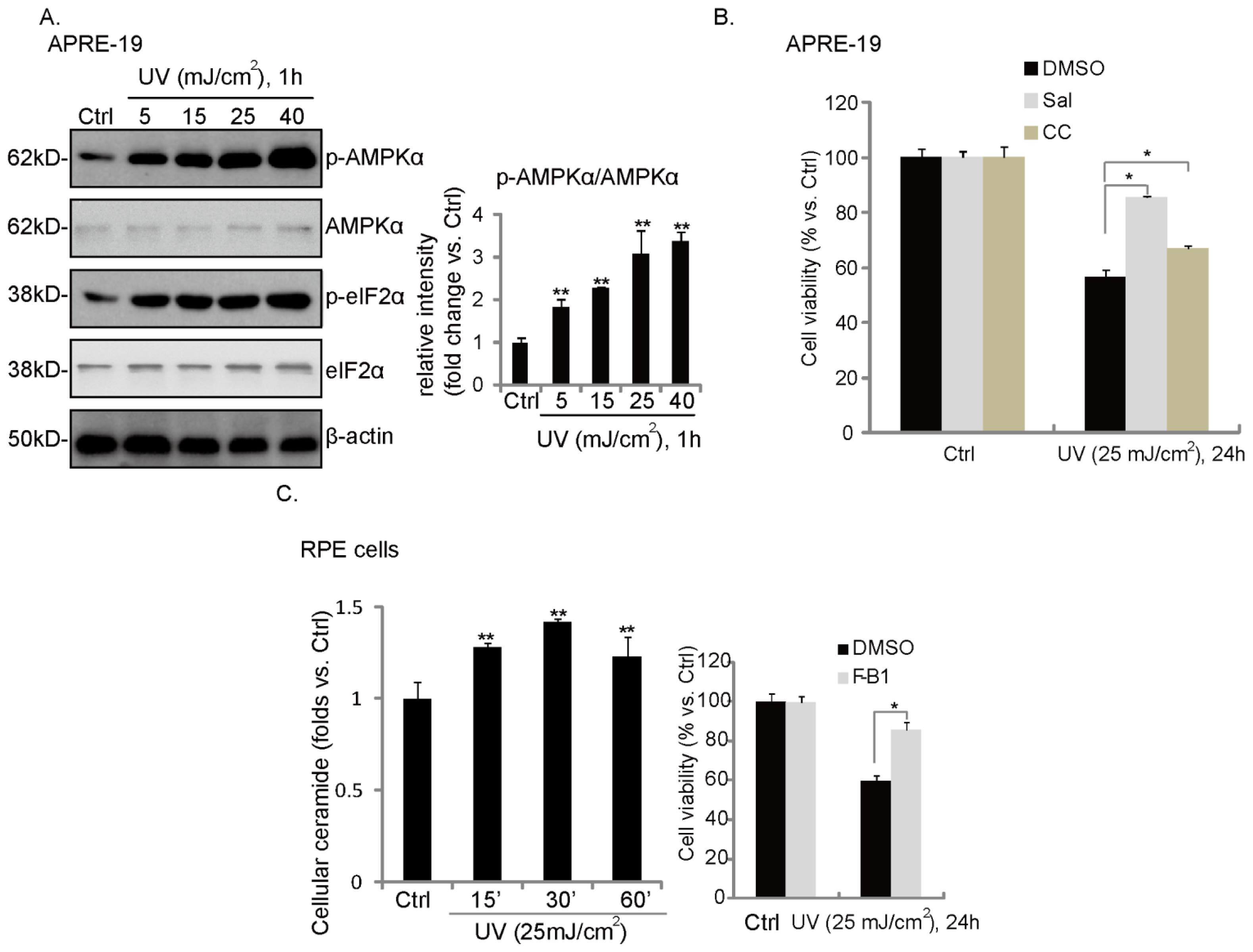
2.5. UV Induces Ceramide Production, ER Stress/AMPK Activation and RPE Cell Death
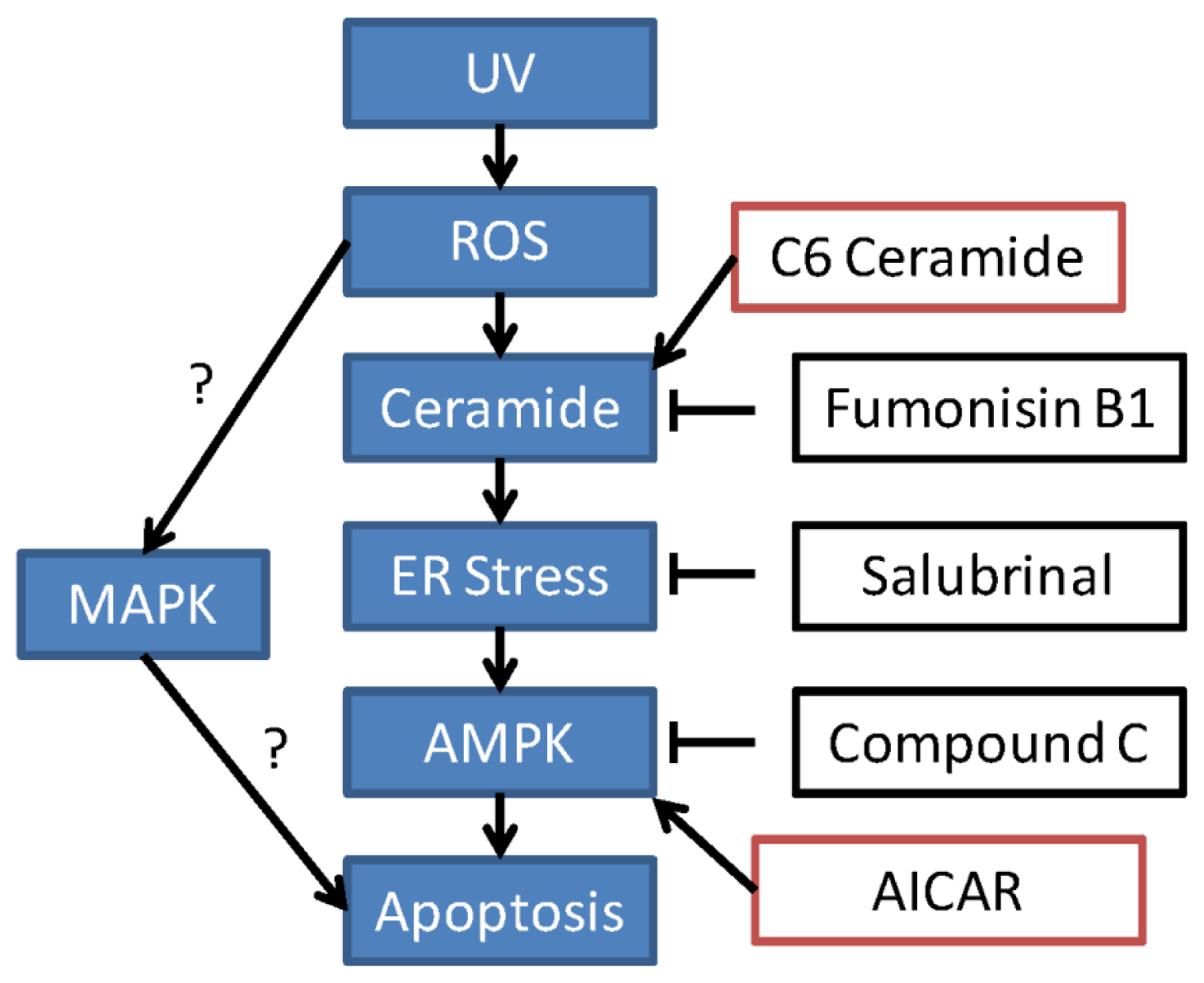
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reagents and Chemicals
4.3. TUNEL Staining and Counting
4.4. Cell Viability Assay
4.5. Measurement of Cellular Ceramide Levels
4.6. Western Blots Analysis
4.7. Statistical Analysis
5. Conclusion
Acknowledgements
Conflict of Interest
Abbreviations
| AMD | age-related macular degeneration |
| Sal | salubrinal |
| H2O2 | hydrogen peroxide |
| AICAR | 5-amino-1-β-D-ribofuranosyl-imidazole-4-carboxamide |
| UV | ultraviolet |
| MTT | 3-[4,5-dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide |
| NGF | nerve growth factor |
| RPE | retinal pigment epithelium |
| ROS | reactive oxygen species |
| mTOR | mammalian target of rapamycin |
| AMPK | AMP activated protein kinase |
| MAPK | mitogen-activated protein kinase |
| ER stress | endoplasmic reticulum stress |
| eIF2α | α-subunit of eukaryotic translation initiation factor 2 |
| TUNEL | terminal deoxynucleotidyl transferase dUTP nick-end labeling. |
References
- Friedman, D.S.; O’Colmain, B.J.; Munoz, B.; Tomany, S.C.; McCarty, C.; de Jong, P.T.; Nemesure, B.; Mitchell, P.; Kempen, J. Prevalence of age-related macular degeneration in the United States. Arch. Ophthalmol 2004, 122, 564–572. [Google Scholar]
- Roduit, R.; Schorderet, D.F. MAP kinase pathways in UV-induced apoptosis of retinal pigment epithelium ARPE19 cells. Apoptosis 2008, 13, 343–353. [Google Scholar]
- Liang, Y.G.; Jorgensen, A.G.; Kaestel, C.G.; Wiencke, A.K.; Lui, G.M.; la Cour, M.H.; Ropke, C.H.; Nissen, M.H. Bcl-2, Bax, and c-Fos expression correlates to RPE cell apoptosis induced by UV-light and daunorubicin. Curr. Eye Res 2000, 20, 25–34. [Google Scholar]
- Nilsson, S.E.; Sundelin, S.P.; Wihlmark, U.; Brunk, U.T. Aging of cultured retinal pigment epithelial cells: Oxidative reactions, lipofuscin formation and blue light damage. Doc. Ophthalmol 2003, 106, 13–16. [Google Scholar]
- Young, R.W. Solar radiation and age-related macular degeneration. Surv. Ophthalmol 1988, 32, 252–269. [Google Scholar]
- Chen, Y.; Gibson, S.B. Is mitochondrial generation of reactive oxygen species a trigger for autophagy? Autophagy 2008, 4, 246–248. [Google Scholar]
- Pacifici, R.E.; Davies, K.J. Protein, lipid and DNA repair systems in oxidative stress: The free-radical theory of aging revisited. Gerontology 1991, 37, 166–180. [Google Scholar]
- Bartlett, H.; Eperjesi, F. A randomised controlled trial investigating the effect of nutritional supplementation on visual function in normal, and age-related macular disease affected eyes: Design and methodology [ISRCTN78467674]. Nutr. J. 2003, 2. [Google Scholar] [CrossRef] [Green Version]
- Richer, S.; Stiles, W.; Statkute, L.; Pulido, J.; Frankowski, J.; Rudy, D.; Pei, K.; Tsipursky, M.; Nyland, J. Double-masked, placebo-controlled, randomized trial of lutein and antioxidant supplementation in the intervention of atrophic age-related macular degeneration: The Veterans LAST study (Lutein Antioxidant Supplementation Trial). Optometry 2004, 75, 216–230. [Google Scholar]
- Cao, G.F.; Liu, Y.; Yang, W.; Wan, J.; Yao, J.; Wan, Y.; Jiang, Q. Rapamycin sensitive mTOR activation mediates nerve growth factor (NGF) induced cell migration and pro-survival effects against hydrogen peroxide in retinal pigment epithelial cells. Biochem. Biophys. Res. Commun 2011, 414, 499–505. [Google Scholar]
- Hannun, Y.A.; Obeid, L.M. Ceramide: An intracellular signal for apoptosis. Trends Biochem. Sci 1995, 20, 73–77. [Google Scholar]
- Kurinna, S.M.; Tsao, C.C.; Nica, A.F.; Jiffar, T.; Ruvolo, P.P. Ceramide promotes apoptosis in lung cancer-derived A549 cells by a mechanism involving c-Jun NH2-terminal kinase. Cancer Res 2004, 64, 7852–7856. [Google Scholar]
- Zheng, Q.Y.; Yao, C.; Jin, F.; Zhang, Y.; Zhang, G.H. Ursolic acid-induced AMP-activated protein kinase (AMPK) activation contributes to growth inhibition and apoptosis in human bladder cancer T24 cells. Biochem. Biophys Res. Commun 2012, 419, 741–747. [Google Scholar]
- Ji, C.; Yang, B.; Yang, Y.L.; He, S.H.; Miao, D.S.; He, L.; Bi, Z.G. Exogenous cell-permeable C6 ceramide sensitizes multiple cancer cell lines to Doxorubicin-induced apoptosis by promoting AMPK activation and mTORC1 inhibition. Oncogene 2010, 29, 6557–6568. [Google Scholar]
- Ji, C.; Yang, Y.L.; Yang, Z.; Tu, Y.; Cheng, L.; Chen, B.; Xia, J.P.; Sun, W.L.; Su, Z.L.; He, L.; et al. Perifosine sensitizes UVB-induced apoptosis in skin cells: New implication of skin cancer prevention? Cell. Signal 2012, 24, 1781–1789. [Google Scholar]
- Cao, C.; Lu, S.; Kivlin, R.; Wallin, B.; Card, E.; Bagdasarian, A.; Tamakloe, T.; Chu, W.M.; Guan, K.L.; Wan, Y. AMP-activated protein kinase contributes to UV- and H2O2-induced apoptosis in human skin keratinocytes. J. Biol. Chem 2008, 283, 28897–28908. [Google Scholar]
- Goldkorn, T.; Balaban, N.; Shannon, M.; Chea, V.; Matsukuma, K.; Gilchrist, D.; Wang, H.; Chan, C. H2O2 acts on cellular membranes to generate ceramide signaling and initiate apoptosis in tracheobronchial epithelial cells. J. Cell. Sci 1998, 111, 3209–3220. [Google Scholar]
- Wang, E.; Norred, W.P.; Bacon, C.W.; Riley, R.T.; Merrill, A.H., Jr. Inhibition of sphingolipid biosynthesis by fumonisins. Implications for diseases associated with Fusarium moniliforme. J. Biol. Chem 1991, 266, 14486–14490. [Google Scholar]
- Merrill, A.H., Jr; van Echten, G.; Wang, E.; Sandhoff, K. Fumonisin B1 inhibits sphingosine (sphinganine) N-acyltransferase and de novo sphingolipid biosynthesis in cultured neurons in situ. J. Biol. Chem 1993, 268, 27299–27306. [Google Scholar]
- Healy, S.J.; Gorman, A.M.; Mousavi-Shafaei, P.; Gupta, S.; Samali, A. Targeting the endoplasmic reticulum-stress response as an anticancer strategy. Eur. J. Pharmacol 2009, 625, 234–246. [Google Scholar]
- Wlodkowic, D.; Skommer, J.; McGuinness, D.; Hillier, C.; Darzynkiewicz, Z. ER-Golgi network—A future target for anti-cancer therapy. Leuk Res 2009, 33, 1440–1447. [Google Scholar]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell. Biol 2012, 13, 89–102. [Google Scholar]
- Urano, F.; Wang, X.; Bertolotti, A.; Zhang, Y.; Chung, P.; Harding, H.P.; Ron, D. Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science 2000, 287, 664–666. [Google Scholar]
- Wu, S.; Hu, Y.; Wang, J.L.; Chatterjee, M.; Shi, Y.; Kaufman, R.J. Ultraviolet light inhibits translation through activation of the unfolded protein response kinase PERK in the lumen of the endoplasmic reticulum. J. Biol. Chem 2002, 277, 18077–18083. [Google Scholar]
- Jiang, H.Y.; Wek, R.C. GCN2 phosphorylation of eIF2alpha activates NF-kappaB in response to UV irradiation. Biochem. J 2005, 385, 371–380. [Google Scholar]
- Wang, S.; Song, P.; Zou, M.H. AMP-activated protein kinase, stress responses and cardiovascular diseases. Clin. Sci. (Lond.) 2012, 122, 555–573. [Google Scholar]
- Vakana, E.; Altman, J.K.; Platanias, L.C. Targeting AMPK in the treatment of malignancies. J. Cell. Biochem 2012, 113, 404–409. [Google Scholar]
- Meisse, D.; van de Casteele, M.; Beauloye, C.; Hainault, I.; Kefas, B.A.; Rider, M.H.; Foufelle, F.; Hue, L. Sustained activation of AMP-activated protein kinase induces c-Jun N-terminal kinase activation and apoptosis in liver cells. FEBS Lett 2002, 526, 38–42. [Google Scholar]
- Zhang, W.B.; Wang, Z.; Shu, F.; Jin, Y.H.; Liu, H.Y.; Wang, Q.J.; Yang, Y. Activation of AMP-activated protein kinase by temozolomide contributes to apoptosis in glioblastoma cells via p53 activation and mTORC1 inhibition. J. Biol. Chem 2010, 285, 40461–40471. [Google Scholar]
- Chen, L.; Xu, B.; Liu, L.; Luo, Y.; Yin, J.; Zhou, H.; Chen, W.; Shen, T.; Han, X.; Huang, S. Hydrogen peroxide inhibits mTOR signaling by activation of AMPKalpha leading to apoptosis of neuronal cells. Lab Invest 2010, 90, 762–773. [Google Scholar]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell. Biol 2011, 13, 1016–1023. [Google Scholar]
- Inoki, K.; Ouyang, H.; Zhu, T.; Lindvall, C.; Wang, Y.; Zhang, X.; Yang, Q.; Bennett, C.; Harada, Y.; Stankunas, K.; et al. TSC2 integrates Wnt and energy signals via a coordinated phosphorylation by AMPK and GSK3 to regulate cell growth. Cell 2006, 126, 955–968. [Google Scholar]
- Hardie, D.G. AMPK and Raptor: Matching cell growth to energy supply. Mol. Cell 2008, 30, 263–265. [Google Scholar]
- Zhu, Q.Y.; Wang, Z.; Ji, C.; Cheng, L.; Yang, Y.L.; Ren, J.; Jin, Y.H.; Wang, Q.J.; Gu, X.J.; Bi, Z.G.; et al. C6-ceramide synergistically potentiates the anti-tumor effects of histone deacetylase inhibitors via AKT dephosphorylation and alpha-tubulin hyperacetylation both in vitro and in vivo. Cell Death Dis 2011, 2, e117. [Google Scholar]
- Van Lummel, M.; van Blitterswijk, W.J.; Vink, S.R.; Veldman, R.J.; van der Valk, M.A.; Schipper, D.; Dicheva, B.M.; Eggermont, A.M.; ten Hagen, T.L.; Verheij, M.; et al. Enriching lipid nanovesicles with short-chain glucosylceramide improves doxorubicin delivery and efficacy in solid tumors. FASEB J 2011, 25, 280–289. [Google Scholar]






© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yao, J.; Bi, H.-E.; Sheng, Y.; Cheng, L.-B.; Wendu, R.-L.; Wang, C.-H.; Cao, G.-F.; Jiang, Q. Ultraviolet (UV) and Hydrogen Peroxide Activate Ceramide-ER Stress-AMPK Signaling Axis to Promote Retinal Pigment Epithelium (RPE) Cell Apoptosis. Int. J. Mol. Sci. 2013, 14, 10355-10368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms140510355
Yao J, Bi H-E, Sheng Y, Cheng L-B, Wendu R-L, Wang C-H, Cao G-F, Jiang Q. Ultraviolet (UV) and Hydrogen Peroxide Activate Ceramide-ER Stress-AMPK Signaling Axis to Promote Retinal Pigment Epithelium (RPE) Cell Apoptosis. International Journal of Molecular Sciences. 2013; 14(5):10355-10368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms140510355
Chicago/Turabian StyleYao, Jin, Hui-E Bi, Yi Sheng, Li-Bo Cheng, Ri-Le Wendu, Cheng-Hu Wang, Guo-Fan Cao, and Qin Jiang. 2013. "Ultraviolet (UV) and Hydrogen Peroxide Activate Ceramide-ER Stress-AMPK Signaling Axis to Promote Retinal Pigment Epithelium (RPE) Cell Apoptosis" International Journal of Molecular Sciences 14, no. 5: 10355-10368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms140510355