Improved Nutritive Quality and Salt Resistance in Transgenic Maize by Simultaneously Overexpression of a Natural Lysine-Rich Protein Gene, SBgLR, and an ERF Transcription Factor Gene, TSRF1

Abstract
:1. Introduction
2. Results and Discussion
2.1. TSRF1, but Not SBgLR Protein Has Homologs in Maize
2.2. Generation of Transgenic Maize Inbred Lines
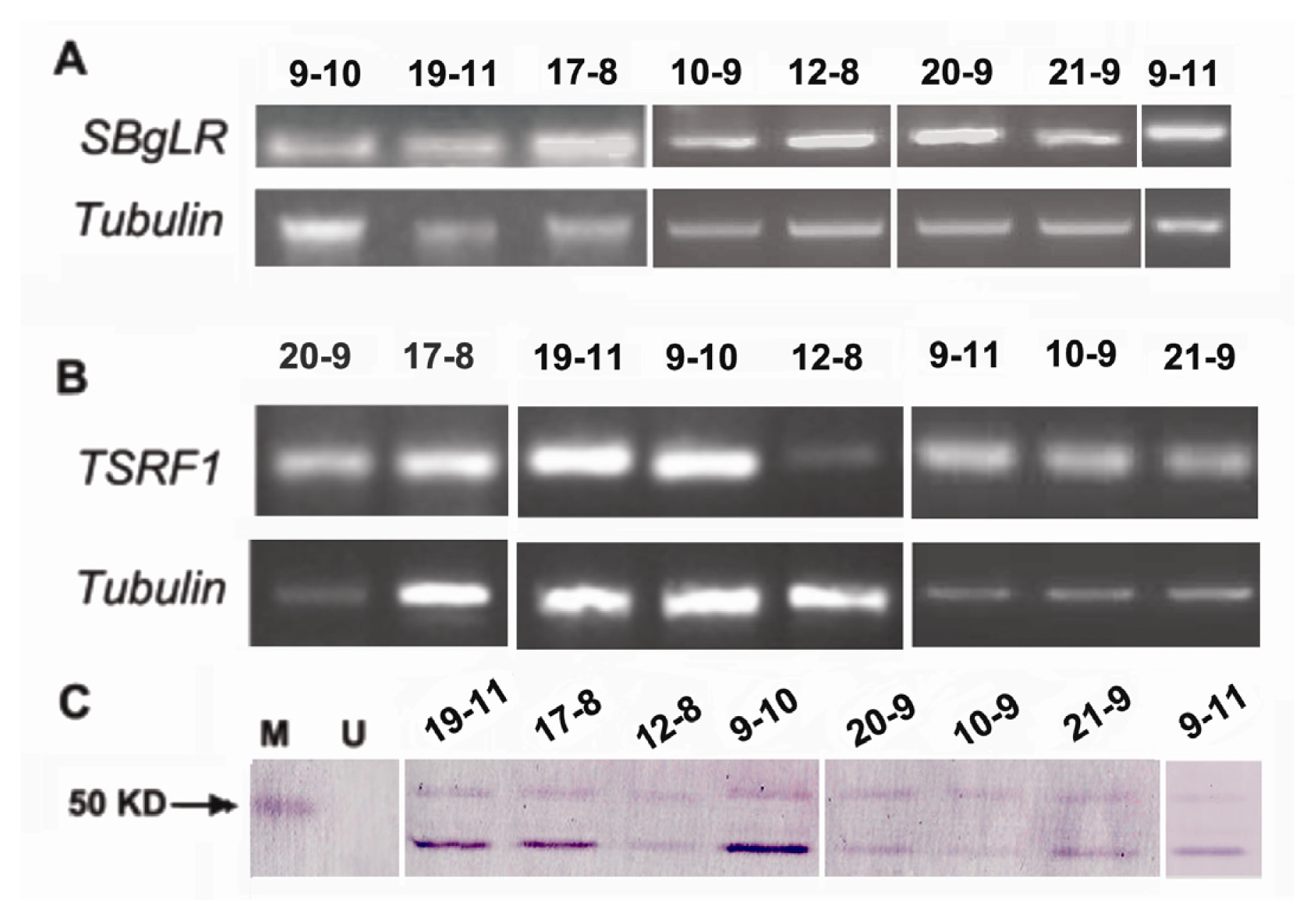
2.3. Overexpression of SBgLR and TSRF1 in Transgenic Maize
2.4. Marker-Free Transgenic Lines Obtained by Segregation in T2 Generation
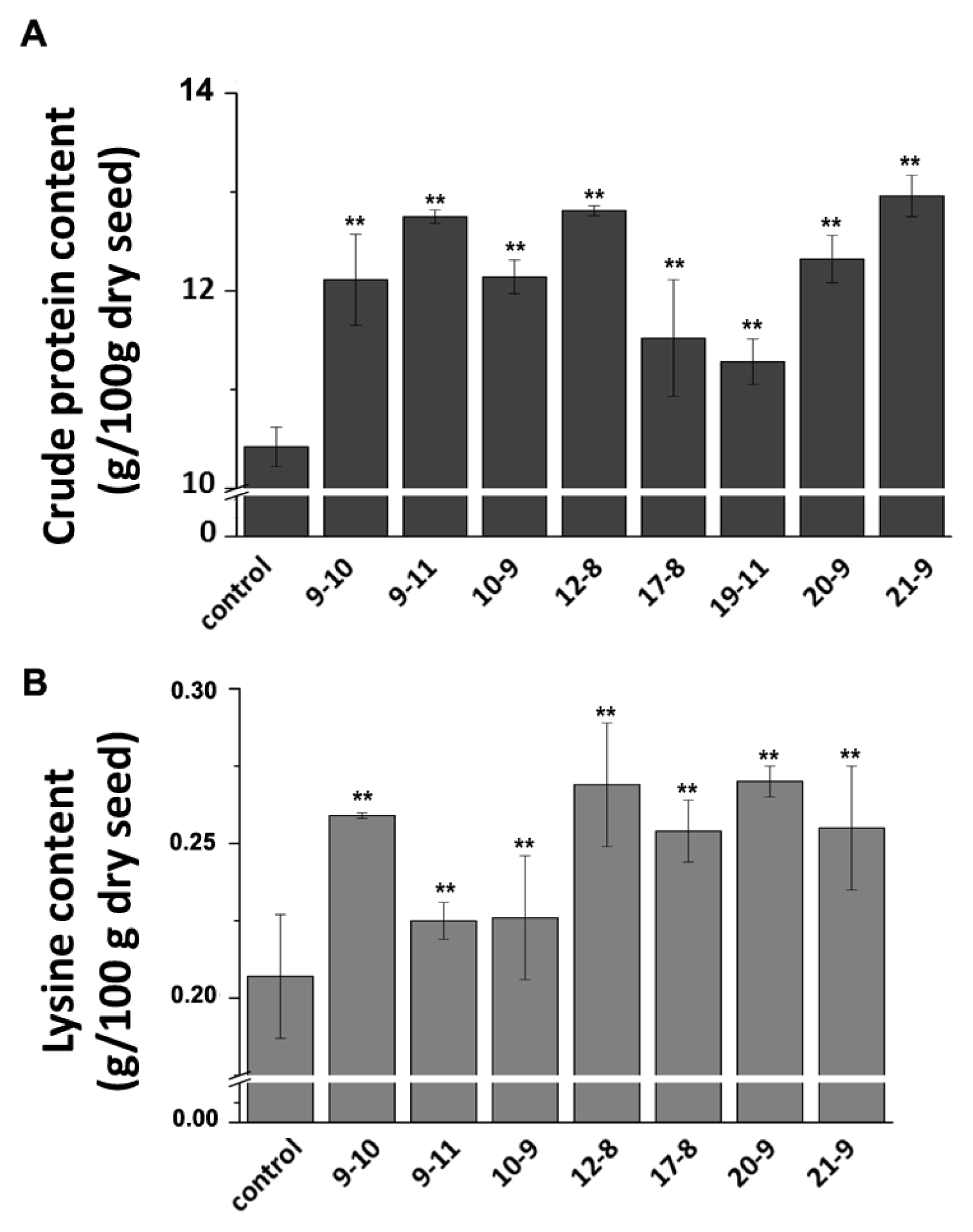
2.5. SBgLR Enhanced Crude Protein and Lysine Contents in Transgenic Maize
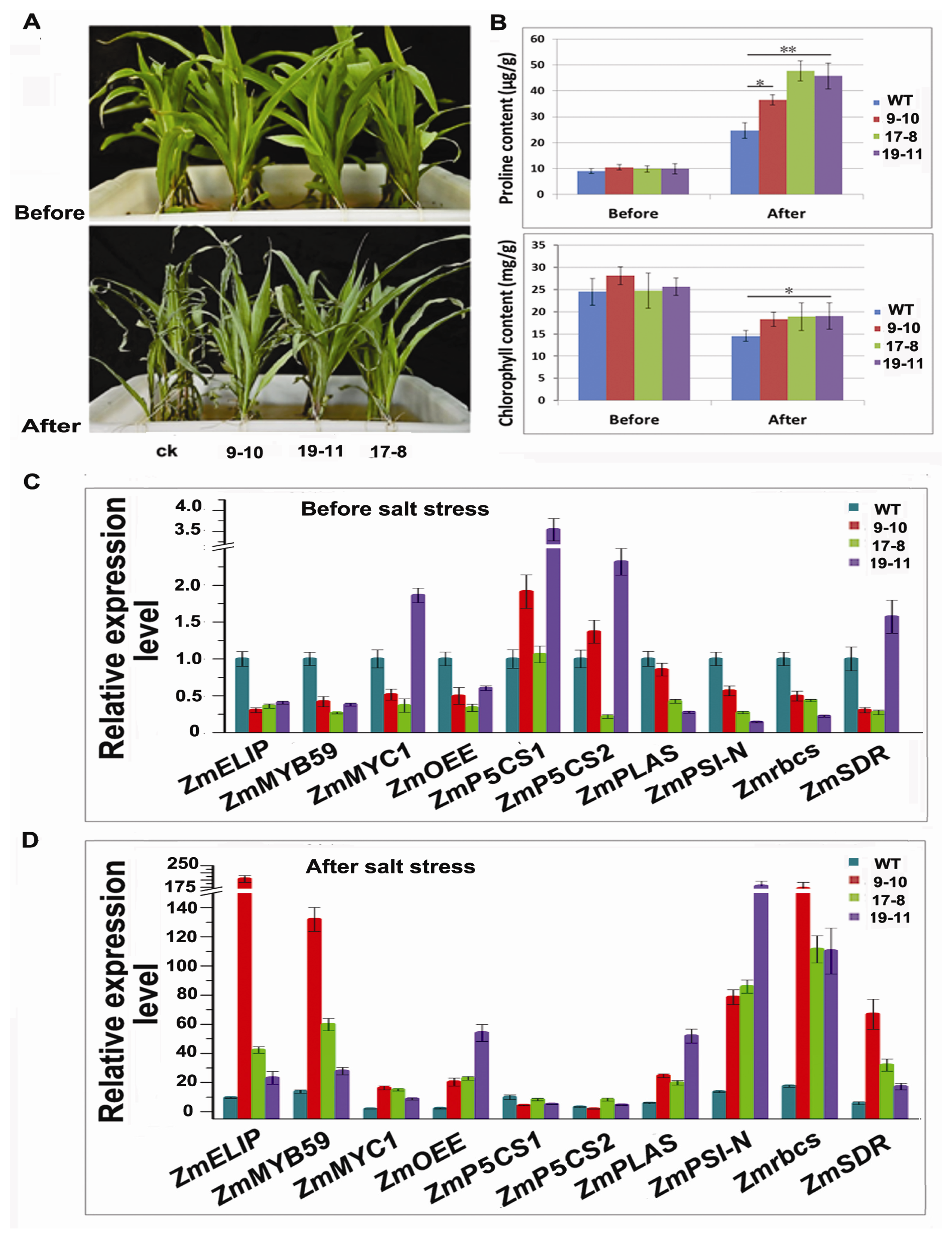
2.6. TSRF1 Increased Salt Tolerance in Transgenic Maize
3. Experimental Section
3.1. Plasmid Construction
3.2. Plant Transformation and Detection
3.3. Salt Stress Treatment
3.4. Detection of Free Proline and Chlorophyll Content
3.5. Gene Expression Analysis by Quantitative Real-Time PCR
3.6. Crude Protein and Lysine Content Analysis
4. Conclusions
Supplemental Information
ijms-14-09459-s001.pdfAcknowledgments
Conflict of Interest
References
- Osborne, T.B.; Mendel, L.B. Nutritional properties of proteins of the maize kernel. J. Biol. Chem 1914, 18, 1–16. [Google Scholar]
- Mertz, E.T.; Bates, L.S.; Nelson, O.E. A mutant gene that changes the protein composition and increases the lysine content of maize endosperm. Science 1964, 145, 279–280. [Google Scholar]
- Vasal, S.K.; Villegas, E.; Bjarnason, M.; Gelaw, B.; Goertz, P. Genetic Modifiers and Breeding Strategies in Developing Hard Endosperm opaque-2 Materials. In Improvement of Quality Traits of Maize for Grain and Silage Use, 2nd ed; Pollmer, W.G., Phipps, R.H., Eds.; Martinus Nijhoff: London, UK, 1980; pp. 37–73. [Google Scholar]
- Segal, G.; Song, R.; Messing, J. A new opaque variant of maize by a single dominant RNA interference-inducing transgene. Genetics 2003, 165, 387–397. [Google Scholar]
- Huang, S.; Adams, W.R.; Zhou, Q.; Malloy, K.P.; Voyles, D.A.; Anthony, J.; Kriz, A.L.; Luethy, M.H. Improving nutritional quality of maize proteins by expressing sense and antisense zein genes. J. Agric. Food Chem 2004, 52, 1958–1964. [Google Scholar]
- Huang, S.; Frizzi, A.; Florida, C.A.; Kruger, D.E.; Luethy, M.H. High lysine and high tryptophan transgenic maize resulting from the reduction of both 19- and 22-kD-zeins. Plant Mol. Biol 2006, 61, 525–535. [Google Scholar]
- Huang, S.; Kruger, D.E.; Frizzi, A.; D’Ordine, R.L.; Florida, C.A.; Adams, W.R.; Brown, W.E.; Luethy, M.H. High-lysine corn produced by the combination of enhanced lysine biosynthesis and reduced zein accumulation. Plant Biotechnol. J 2005, 3, 555–569. [Google Scholar]
- Houmard, N.M.; Mainville, J.L.; Bonin, C.P.; Huang, S.; Luethy, M.H.; Malvar, T.M. High lysine corn generated by endosperm specific suppression of lysine catabolism using RNAi. Plant Biotechnol. J 2007, 5, 605–614. [Google Scholar]
- Lucas, D.M.; Glenn, K.C.; Bu, J.Y. Petition for determination of nonregulated status for lysine maize LY038. Available online: http://www.aphis.usda.gov/brs/aphisdocs/0422901p.pdf (accessed on 13 August 2004).
- Yang, S.H.; Moran, D.L.; Jia, H.W.; Bicar, E.H.; Lee, M.; Scott, M.P. Expression of a synthetic porcine-lactalbumin gene in the kernels of transgenic maize. Transgenic Res 2002, 11, 11–20. [Google Scholar]
- Bicar, E.H.; Woodman-Clikeman, W.; Sangtong, V.; Peterson, J.M.; Yang, S.S.; Lee, M.; Scott, M.P. Transgenic maize endosperm containing a milk protein has improved amino acid balance. Transgenic Res 2008, 17, 59–71. [Google Scholar]
- Liu, J.Q.; Seul, U.; Thompson, R. Cloning and characterization of pollen-specific cDNA encoding a glutamic acid-rich protein (GARP) from potato Solanum berthautii. Plant Mol. Biol 1997, 33, 291–300. [Google Scholar]
- Yu, J.; Peng, P.; Zhang, X.; Zhao, Q.; Zhu, D.; Sun, X.; Liu, J.; Ao, G. Seed-specific expression of a lysine rich protein sb401 gene significantly increases both lysine and total protein content in maize seeds. Mol. Breed 2004, 14, 1–7. [Google Scholar]
- Lang, Z.; Zhao, Q.; Yu, J.; Zhu, D.; Ao, G. Cloning of potato SBgLR gene and its intron splicing in transgenic maize. Plant Sci 2004, 166, 1227–1233. [Google Scholar]
- Xiong, L.; Schumaker, K.S.; Zhu, J. Cell signaling during cold, drought and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar]
- Xiong, L.; Zhu, J.K. Regulation of abscisic acid biosynthesis. Plant Physiol 2003, 133, 29–36. [Google Scholar]
- Guo, H.; Ecker, J.R. The ethylene signaling pathway: New insights. Curr. Opin. Plant Biol 2004, 7, 40–49. [Google Scholar]
- Stockinger, E.J.; Gilmour, A.J.; Thomashow, M.F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 1997, 94, 1035–1040. [Google Scholar]
- Chakravarthy, S.; Tuori, R.P.; D’Ascenzo, M.D.; Fobert, P.R.; Despres, C.; Martin, G.B. The tomato transcription factor Pti4 regulates defense-related gene expression via GCC box and non-GCC box cis elements. Plant Cell 2003, 15, 3033–3050. [Google Scholar]
- Guo, Z.J.; Chen, X.J.; Wu, X.L.; Ling, J.Q.; Xu, P. Overexpression of the AP2/EREBP transcription factor OPBP1 enhances disease resistance and salt tolerance in tobacco. Plant Mol. Biol 2004, 55, 607–618. [Google Scholar]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol 2006, 57, 781–803. [Google Scholar]
- Mantiri, F.R.; Kurdyukov, S.; Lohar, D.P.; Sharopova, N.; Saeed, N.A.; Wang, X.D.; Vandenbosch, K.A.; Rose, R.J. The transcription factor MtSERF1 of the ERF subfamily identiWed by transcriptional prowling is required for somatic embryogenesis induced by auxin plus cytokinin in Medicago truncatula. Plant Physiol 2008, 146, 1622–1636. [Google Scholar]
- Ohme-Takagi, M.; Shinshi, H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell 1995, 7, 173–182. [Google Scholar]
- Fujimoto, S.Y.; Ohta, M.; Usui, A.; Shinshi, H.; Ohme-Takagi, M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 2000, 12, 393–404. [Google Scholar]
- Gu, Y.Q.; Wildermuth, M.C.; Chakravarthy, S.; Loh, Y.T.; Yang, C.; He, X.; Han, Y.; Martin, G.B. Tomato transcription factors pti4, pti5, and pti6 activate defense responses when expressed in Arabidopsis. Plant Cell 2002, 14, 817–831. [Google Scholar]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar]
- Hong, J.P.; Kim, W.T. Isolation and functional characterization of the Ca-DREBLP1 gene encoding a dehydration-responsive element binding-factor-like protein 1 in hot pepper (Capsicum annuum L. cv. Pukang). Planta 2005, 220, 875–888. [Google Scholar]
- Liu, N.; Zhong, N.Q.; Wang, G.L.; Li, L.J.; Liu, X.L.; He, Y.K.; Xia, G.X. Cloning and functional characterization of PpDBF1 gene encoding a DRE-binding transcription factor from Physcomitrella patens. Planta 2007, 226, 827–838. [Google Scholar]
- Zhang, H.; Zhang, D.; Chen, J.; Yang, Y.; Huang, Z.; Huang, D.; Wang, X.; Huang, R. Tomato stress-responsive factor TSRF1 interacts with ethylene responsive element GCC box and regulates pathogen resistance to Ralstonia solanacearum. Plant Mol. Biol 2004, 55, 825–834. [Google Scholar]
- Zhang, H.; Li, W.; Chen, J.; Yang, Y.; Zhang, Z.; Wang, X.C.; Huang, R. Transcriptional activator TSRF1 reversely regulates pathogen resistance and osmotic stress tolerance in tobacco. Plant Mol. Biol 2007, 63, 63–71. [Google Scholar]
- Zhou, J.; Zhang, H.; Yang, Y.; Zhang, Z.; Zhang, H.; Hu, X.; Chen, J.; Wang, X.C.; Huang, R. Abscisic acid regulates TSRF1-mediated resistance to Ralstonia solanacearum by modifying the expression of GCC box-containing genes in tobacco. J. Exp. Bot 2008, 59, 645–652. [Google Scholar]
- Quan, R.; Hu, S.; Zhang, Z.; Zhang, H.; Zhang, Z.; Huang, R. Overexpression of an ERF transcription factor TSRF1 improves rice drought tolerance. Plant Biotech. J 2010, 8, 476–488. [Google Scholar]
- Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K. Transgenic approaches for abiotic stress tolerance in plants: Retrospect and prospects. Plant Cell Rep 2008, 27, 411–424. [Google Scholar]
- James, C. Global Status of Commercialized Biotech/GM Crops: 2010. ISAAA Brief No. 42; ISAAA: Ithaca, NY, USA, 2010. [Google Scholar]
- Tuteja, N.; Verma, S.; Sahoo, R.K.; Raveendar, S.; Reddy, B.L. Recent advances in development of marker-free transgenic plants: Regulation and biosafety concern. J. Biosci 2012, 37, 167–197. [Google Scholar]
- Holger, P. Removing selectable marker genes: Taking shortcut. Trends Plant Sci 2000, 7, 273–274. [Google Scholar]
- De Neve, M.; de Buck, S.; Jacobs, A.; van Montagu, M.; Depicker, A. T-DNA integration patterns in co-transformed plant cells suggest that T-DNA repeats originate from co-integration of separate T-DNAs. Plant J 1997, 11, 15–29. [Google Scholar]
- Sripriya, R.; Raghupathy, V.; Veluthambi, K. Generation of selectable marker-free sheath blight resistant transgenic rice plants by efficient co-transformation of a cointegrate vector T-DNA and a binary vector T-DNA in one Agrobacterium tumefaciens strain. Plant Cell Rep 2008, 27, 1635–1644. [Google Scholar]
- Miller, M.; Tagliani, L.; Wang, N.; Berka, B.; Bidney, D. High efficiency transgene segregation in co-transformed maize plants using an Agrobacterium tumefaciens 2 T-DNA binary system. Transgenic Res 2002, 11, 381–396. [Google Scholar]
- Shiva, P.N.; Bhojaraja, R.; Shivbachan, S.K.; Hari, P.G.G.; Nagraj, T.K.; Prasad, V.; Srikanth, B.V.; Jayaprakash, T.L.; Dasgupta, S.; Spencer, T.M.; et al. Marker-free transgenic corn plant production through co-bombardment. Plant Cell Rep 2009, 28, 1655–1668. [Google Scholar]
- Kumar, S.; Arul, L.; Talwar, D. Generation of marker-free Bt transgenic indica rice and evaluation of its yellow stem borer resistance. J. Appl. Genet 2010, 51, 243–257. [Google Scholar]
- Qi, Y.; Ye, S.; Lu, Y.; Jin, Q.; Zhang, X. Development of marker-free transgenic Cry1Ab rice with Lepidopteran pest resistance by Agrobacterium mixture-mediated co-transformation. Rice Sci 2009, 16, 181–186. [Google Scholar]
- Ramana, R.M.V.; Parameswari, C.; Sripriya, R.; Veluthambi, K. Transgene stacking and marker elimination in transgenic rice by sequential Agrobacterium-mediated co-transformation with the same selectable marker gene. Plant Cell Rep 2011, 30, 1241–1252. [Google Scholar]
- Wang, D.; Zhao, Q.; Zhu, D.; Ao, G.; Yu, J. Particle-bombardment-mediated co-transformation of maize with a lysine rich protein gene (sb401) from potato. Euphytica 2006, 150, 75–85. [Google Scholar]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 20, 1112–1115. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol 2006, 140, 411–432. [Google Scholar]
- Clustal, W. Available online: http://www.ebi.ac.uk/Tools/clustalw2/index.html (accessed on 10 September 2007).
- Song, R.; Segal, G.; Messing, J. Expression of the sorghum 10-member kafirin gene cluster in maize endosperm. Nucleic Acids Res 2004, 32, e189. [Google Scholar]
- Huang, S.; Jin, L.; Du, J.; Li, H.; Zhao, Q.; Ou, G.; Ao, G.; Yuan, M. SB401, a pollen-specific protein from Solanum berthaultii, binds to and bundles microtubules and F-actin. Plant J 2007, 51, 406–418. [Google Scholar]
- Noble, M.; Lewis, S.; Cowan, N.J. The microtubule binding domain of microtubule-associated protein MAP1B contains a repeated sequence motif unrelated to that of MAP2 and Tau. J. Cell Biol 1989, 109, 3367–3376. [Google Scholar]
- Wiche, G.; Oberkanins, C.; Himmler, A. Molecular structure and function of microtubule-associated proteins. Int. Rev. Cytol 1991, 124, 217–273. [Google Scholar]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci 2005, 88, 424–438. [Google Scholar]
- Stepien, P.; Klobus, G. Water relations and photosynthesi in Cucumis sativus L. leaves under salt stress. Biologia Plantarum 2006, 50, 610–616. [Google Scholar]
- Zhang, H.; Yang, Y.; Zhang, Z.; Chen, J.; Wang, X.C.; Huang, R. Expression of the ethylene response factor gene TSRF1 enhances abscisic acid responses during seedling development in tobacco. Planta 2008, 228, 777–787. [Google Scholar]
- Savoure, A.; Jaoua, S.; Hua, X.J.; Ardiles, W.; van Montagu, M.; Verbruggen, N. Isolation, characterization, and chromosomal location of a gene encoding the DELTA-1-pyrroline-5- carboxylate synthetase in Arabidopsis thaliana. FEBS Lett 1995, 372, 13–19. [Google Scholar]
- Yoshiba, Y.; Kiyosue, T.; Katagiri, T.; Ueda, H.; Mizoguchi, T.; Yamaguchi-Shinozaki, K.; Wada, K.; Harada, Y.; Shinozaki, K. Correlation between the induction of a gene for delta 1-pyrroline-5-carboxylate synthase and the accumulation of proline in Arabidopsis thaliana under osmotic stress. Plant J 1995, 7, 751–760. [Google Scholar]
- Zhang, C.S.; Lu, Q.; Verma, D.P.S. Removal of feedback inhibition of delta 1-pyrroline-5- carboxylate synthase, a bifunctional enzyme catalyzing the first two steps of proline biosynthesis in plants. J. Biol. Chem 1995, 270, 20491–20496. [Google Scholar]
- Strizhov, N.; Abraham, E.; Okresz, L.; Blickling, S.; Zilberstein, A.; Schell, J.; Koncz, C.; Szabados, L. Differential expression of two P5CS genes controlling proline accumulation during salt-stress requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis. Plant J 1997, 12, 557–569. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed; Cold Spring Harbor Press: New York, NY, USA, 1989. [Google Scholar]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar]
- Walker, N.J. A technique whose time has come. Science 2002, 296, 557. [Google Scholar]
- geNorm v3.5 software. Available online: http://medgen.ugent.be/~jvdesomp/genorm/ (accessed on 7 May 2002).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratio of pTSSB/pHpt | No. of transformed calli | No. of hygromycinresistant events recovered | No. of regenerated plants | PCR analysis | Co-transformation efficiency (%) |
|---|---|---|---|---|---|
| SBgLR (+) TSRF1 (+) Hpt (+) | |||||
| 1.5:1 | 2411 | 224 | 114 | 26 | 1.08 a |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, M.; Liu, C.; Li, S.; Zhu, D.; Zhao, Q.; Yu, J. Improved Nutritive Quality and Salt Resistance in Transgenic Maize by Simultaneously Overexpression of a Natural Lysine-Rich Protein Gene, SBgLR, and an ERF Transcription Factor Gene, TSRF1. Int. J. Mol. Sci. 2013, 14, 9459-9474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14059459
Wang M, Liu C, Li S, Zhu D, Zhao Q, Yu J. Improved Nutritive Quality and Salt Resistance in Transgenic Maize by Simultaneously Overexpression of a Natural Lysine-Rich Protein Gene, SBgLR, and an ERF Transcription Factor Gene, TSRF1. International Journal of Molecular Sciences. 2013; 14(5):9459-9474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14059459
Chicago/Turabian StyleWang, Meizhen, Chen Liu, Shixue Li, Dengyun Zhu, Qian Zhao, and Jingjuan Yu. 2013. "Improved Nutritive Quality and Salt Resistance in Transgenic Maize by Simultaneously Overexpression of a Natural Lysine-Rich Protein Gene, SBgLR, and an ERF Transcription Factor Gene, TSRF1" International Journal of Molecular Sciences 14, no. 5: 9459-9474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14059459