Acidosis Decreases c-Myc Oncogene Expression in Human Lymphoma Cells: A Role for the Proton-Sensing G Protein-Coupled Receptor TDAG8

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. c-Myc Protein Is Downregulated by Acidic pH Treatment in U937 Lymphoma Cells
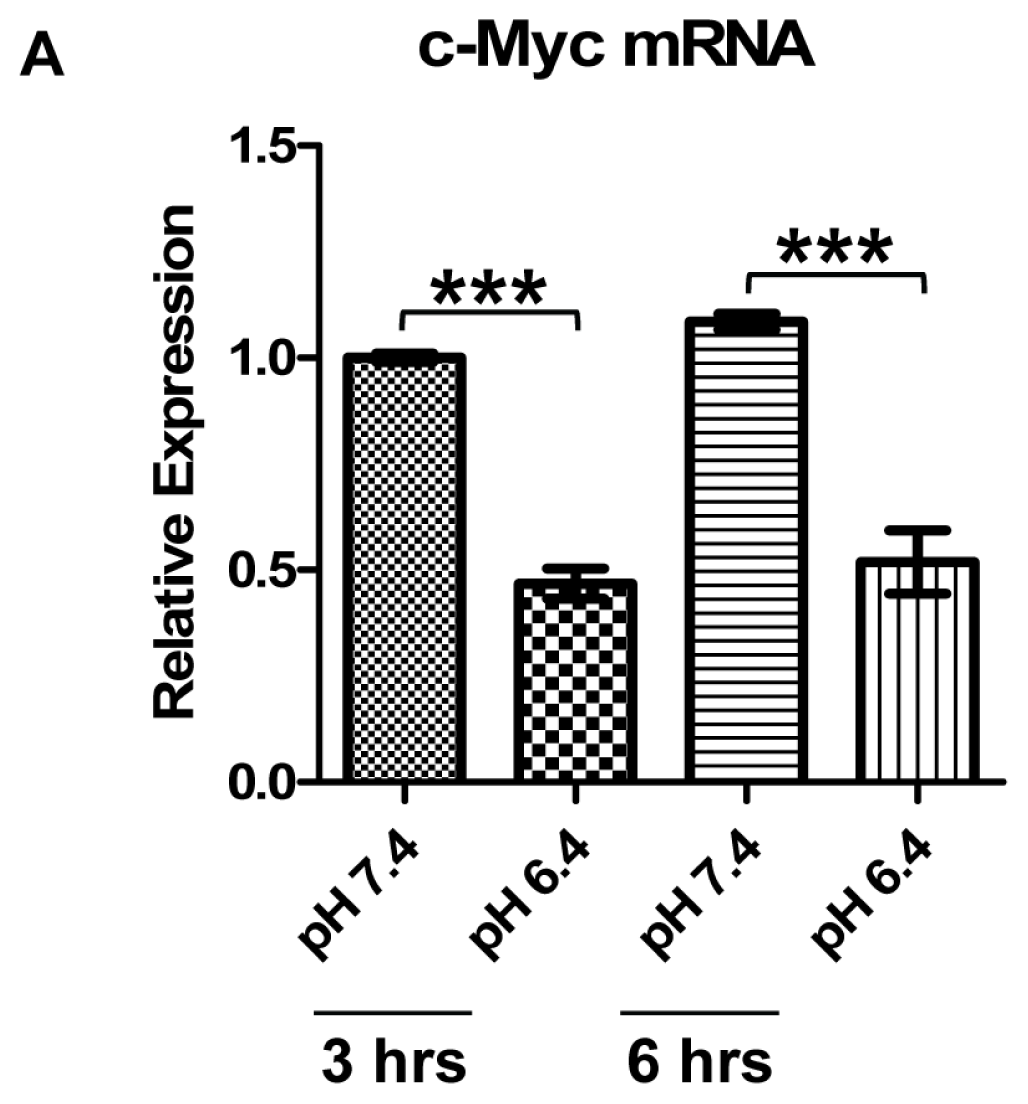
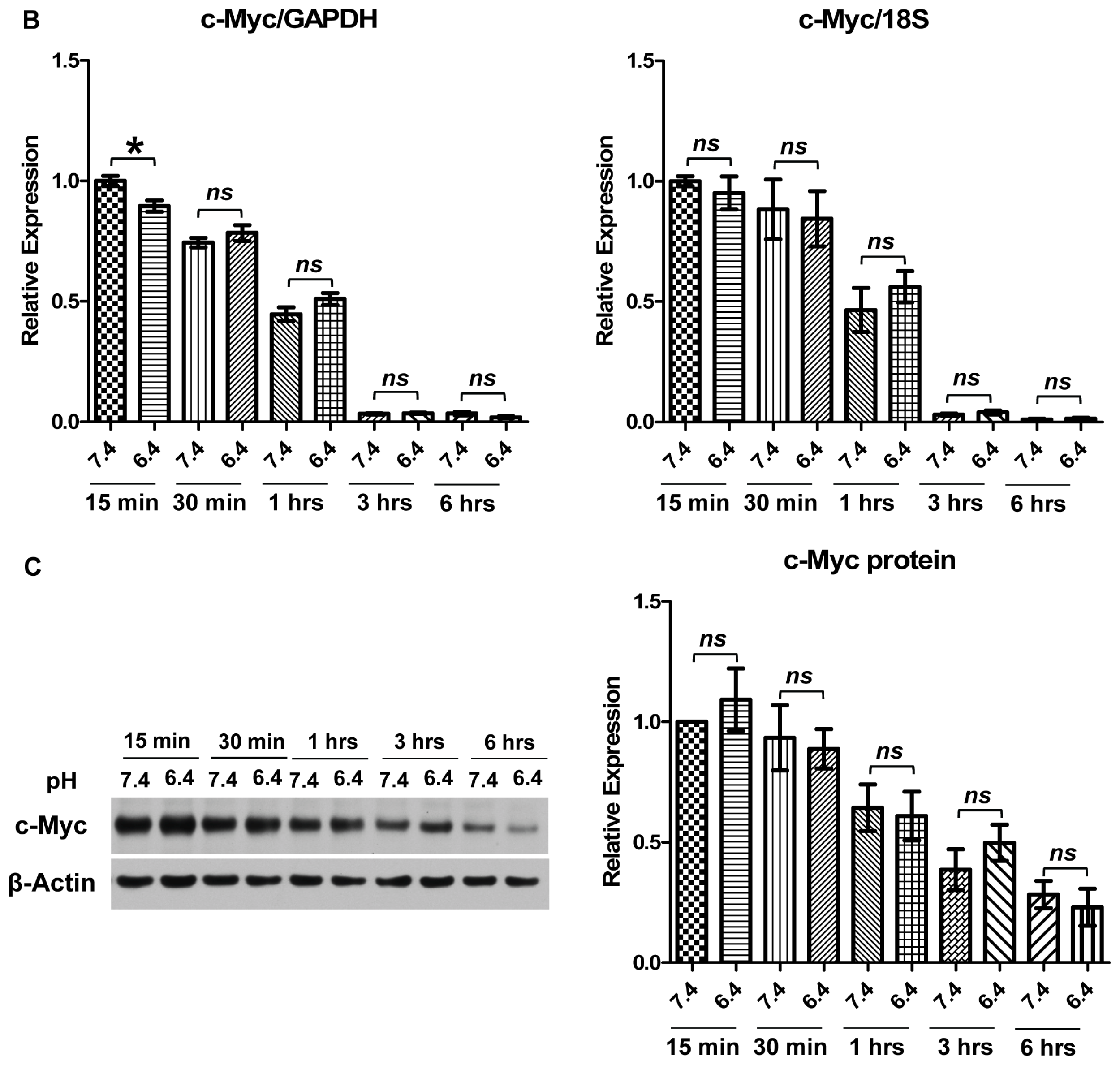
2.2. Downregulation of c-Myc Protein by Acidosis Is Due to Reduced c-Myc Transcriptional Level, but not mRNA or Protein Stability, in U937 Lymphoma Cells
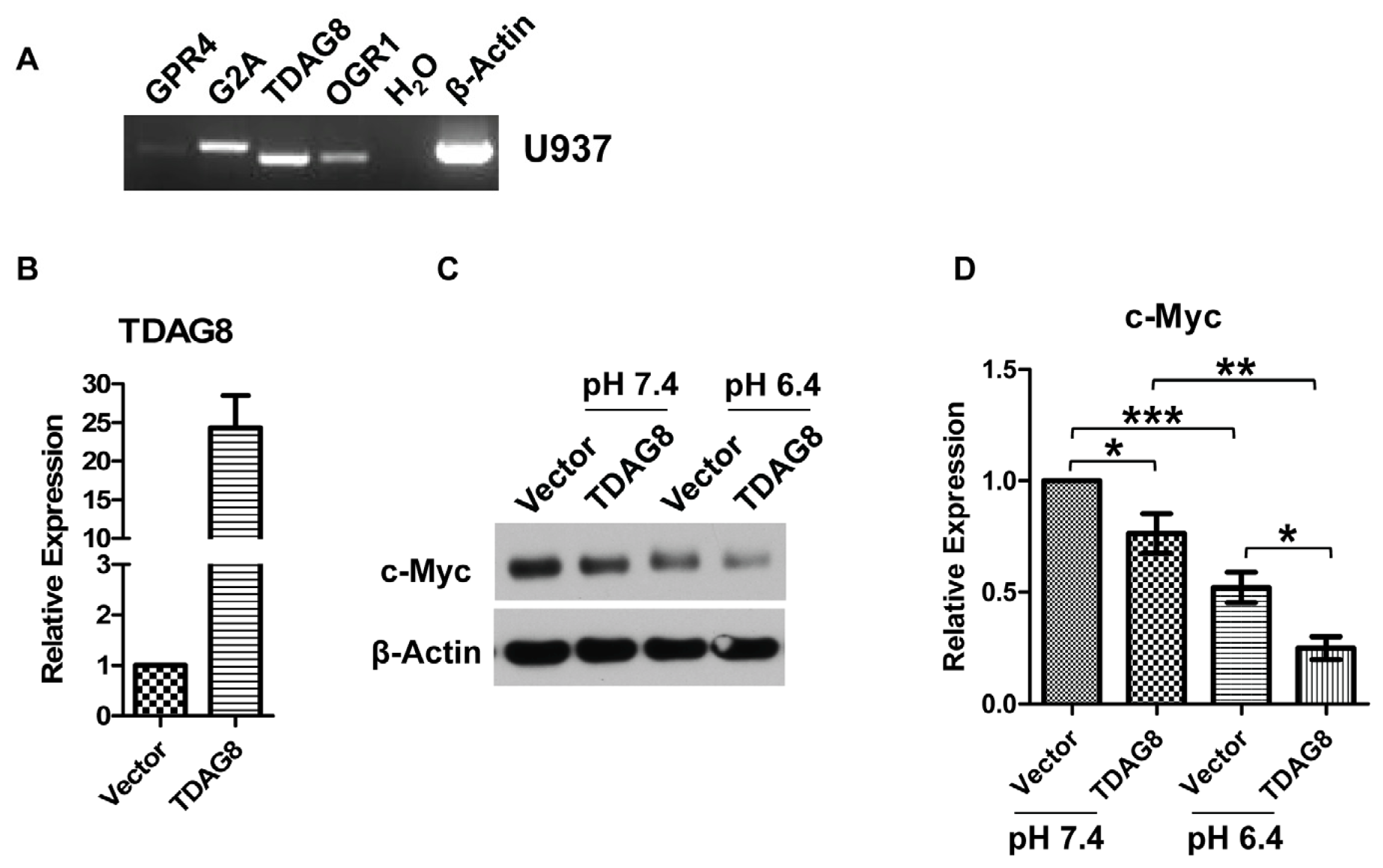
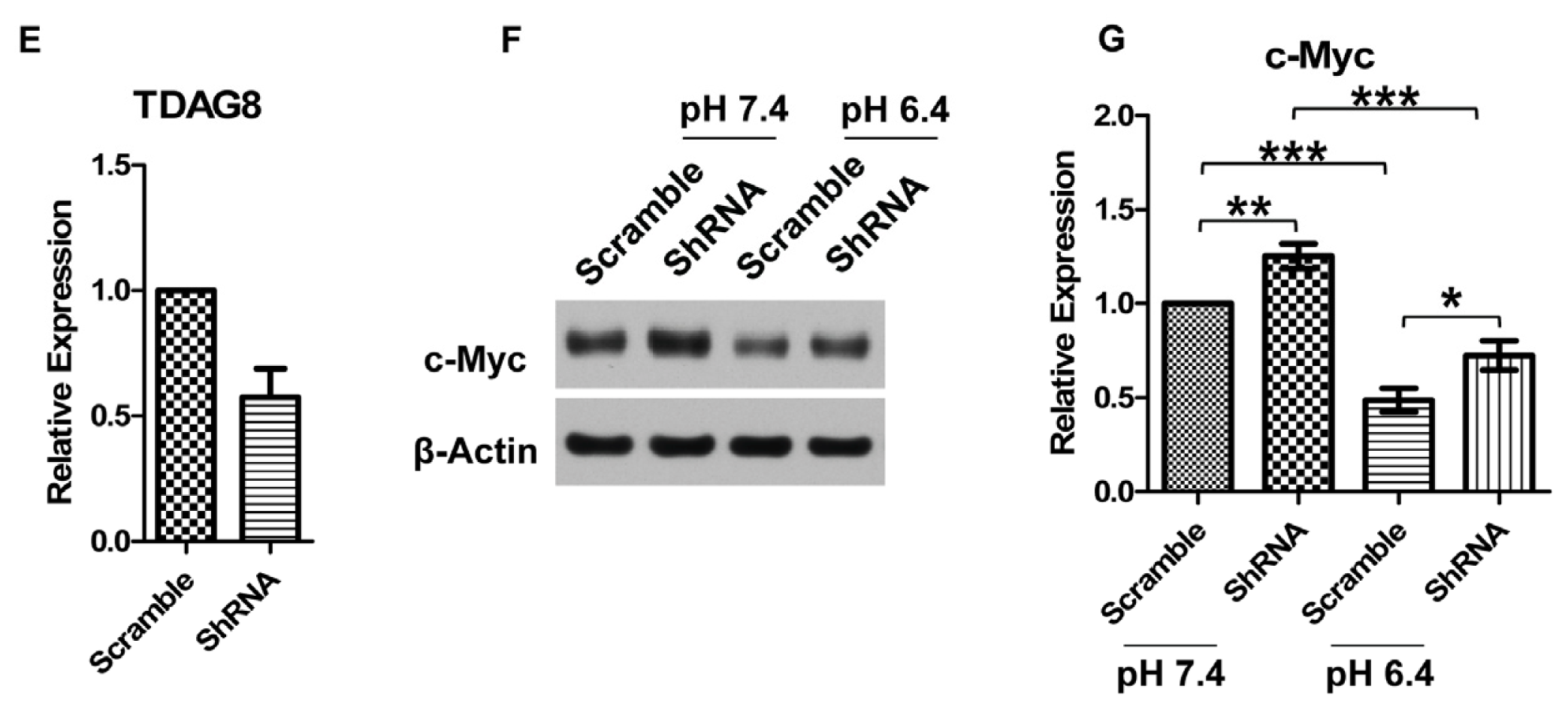
2.3. c-Myc Protein Is Decreased by TDAG8 Overexpression and Increased by TDAG8 Knockdown in U937 Lymphoma Cells
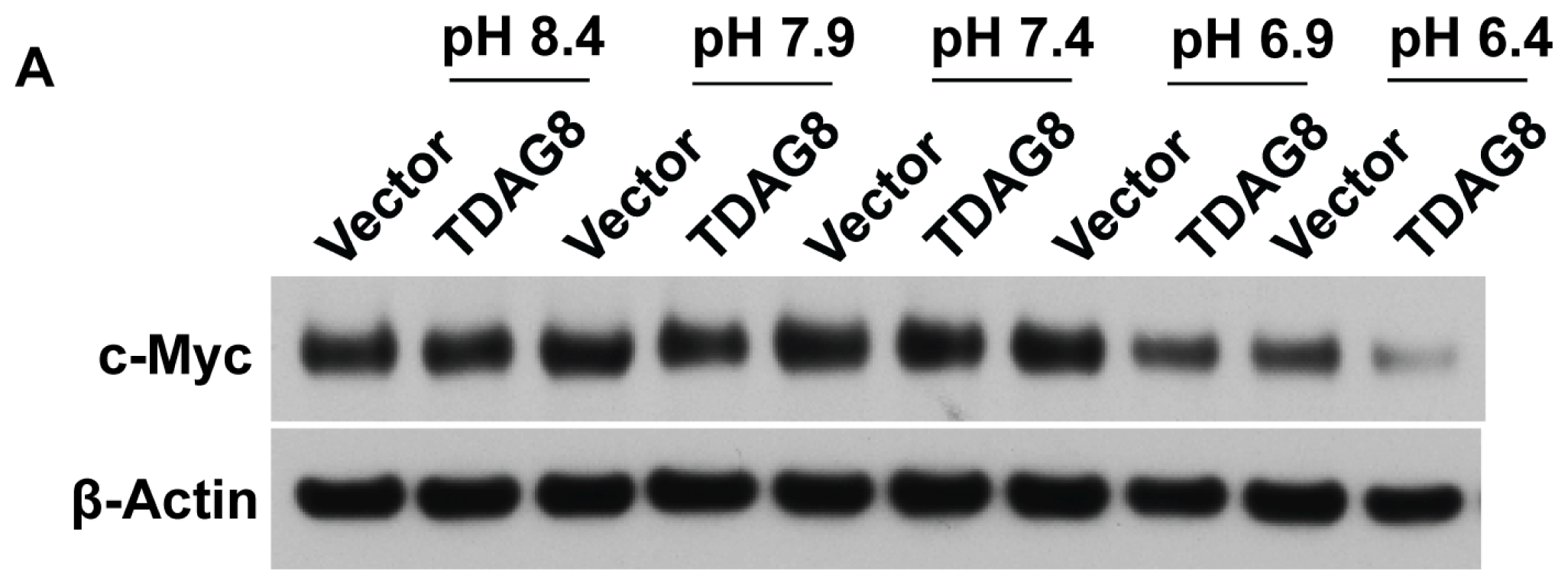
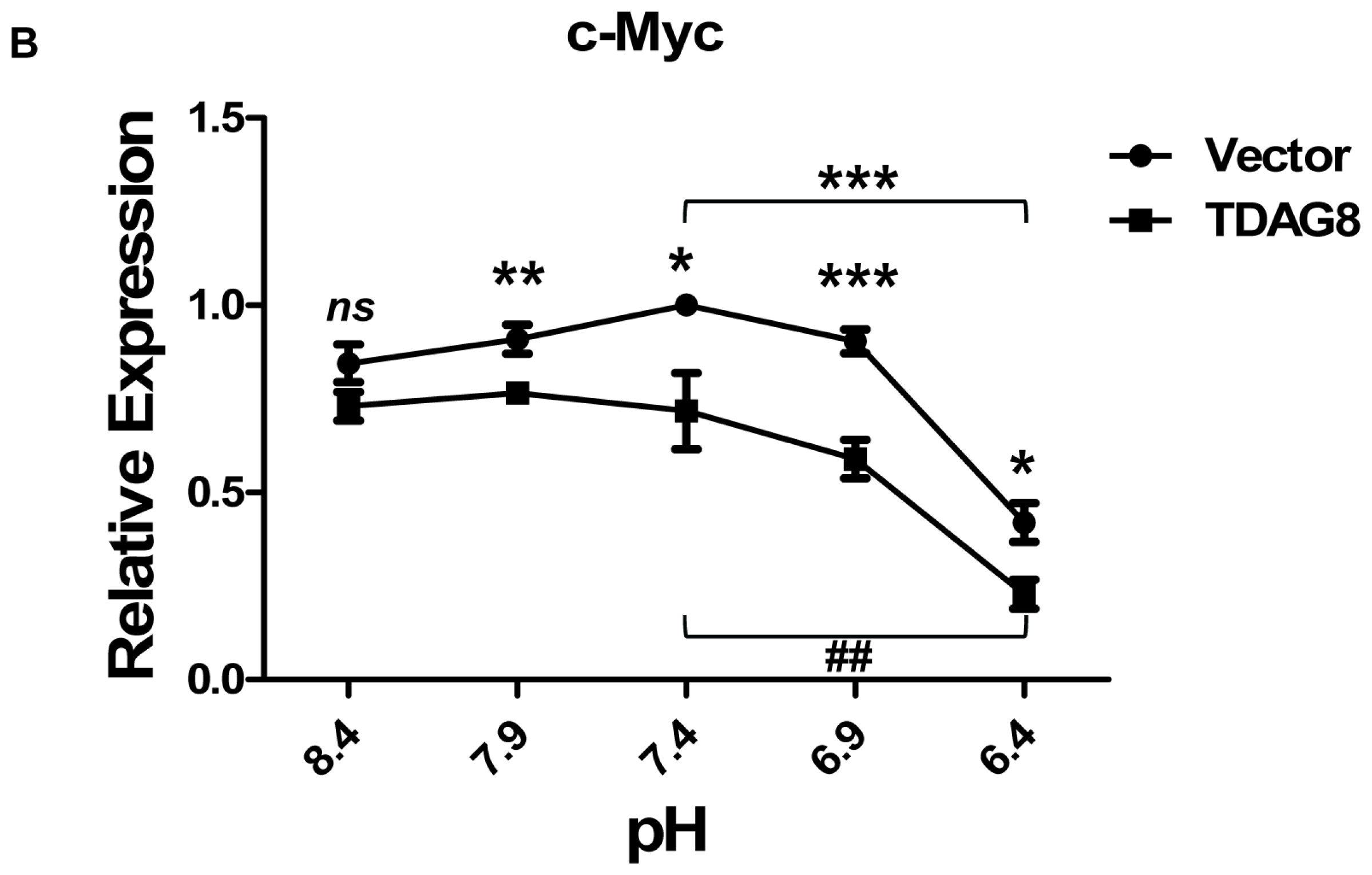
2.4. TDAG8 Downregulates c-Myc Protein in a pH-Dependent Manner
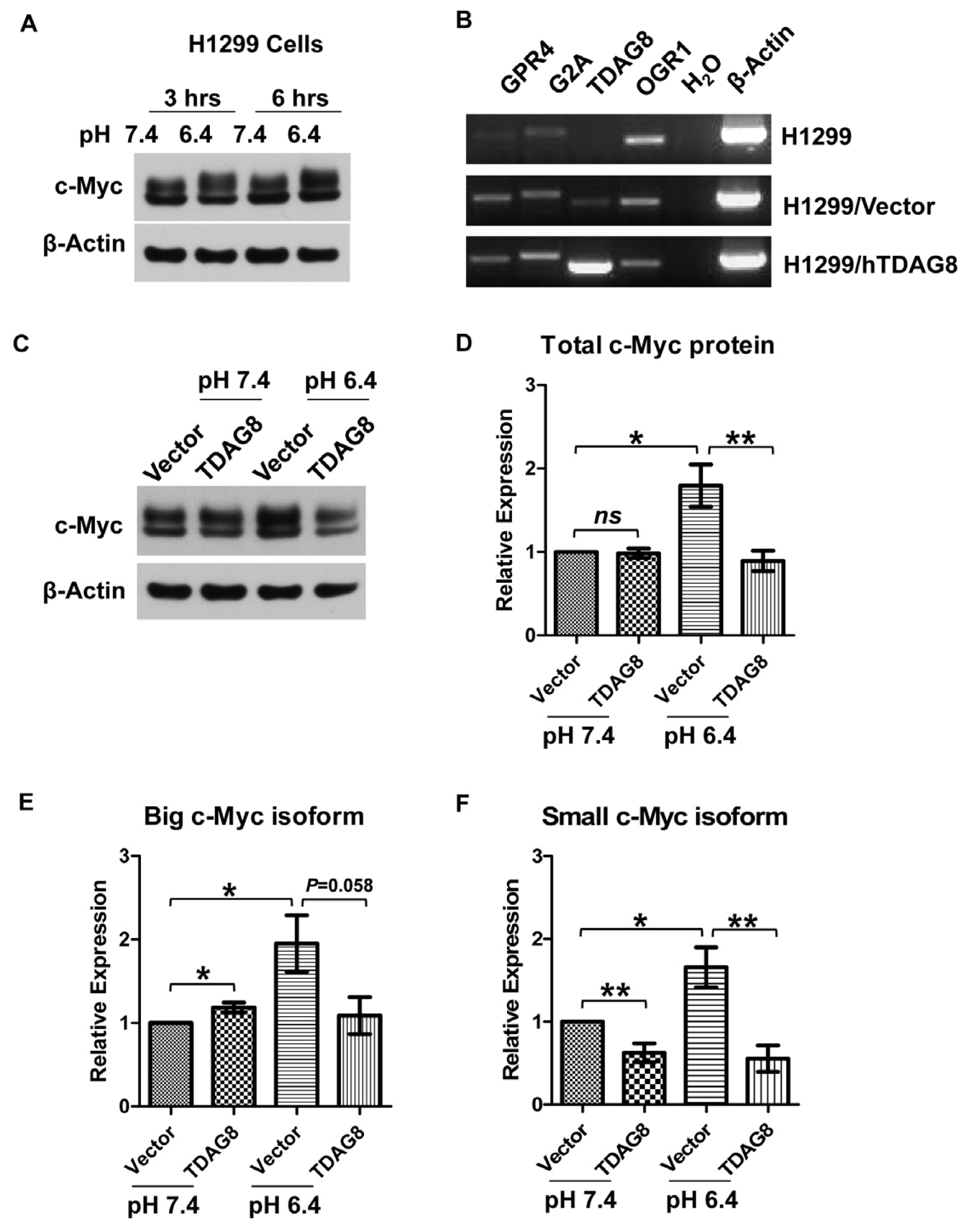
2.5. The Protein Level of c-Myc Is Moderately Increased by Acidosis in H1299 Lung Cancer Cells, but This Increase Is Completely Inhibited by Ectopic TDAG8 Overexpression
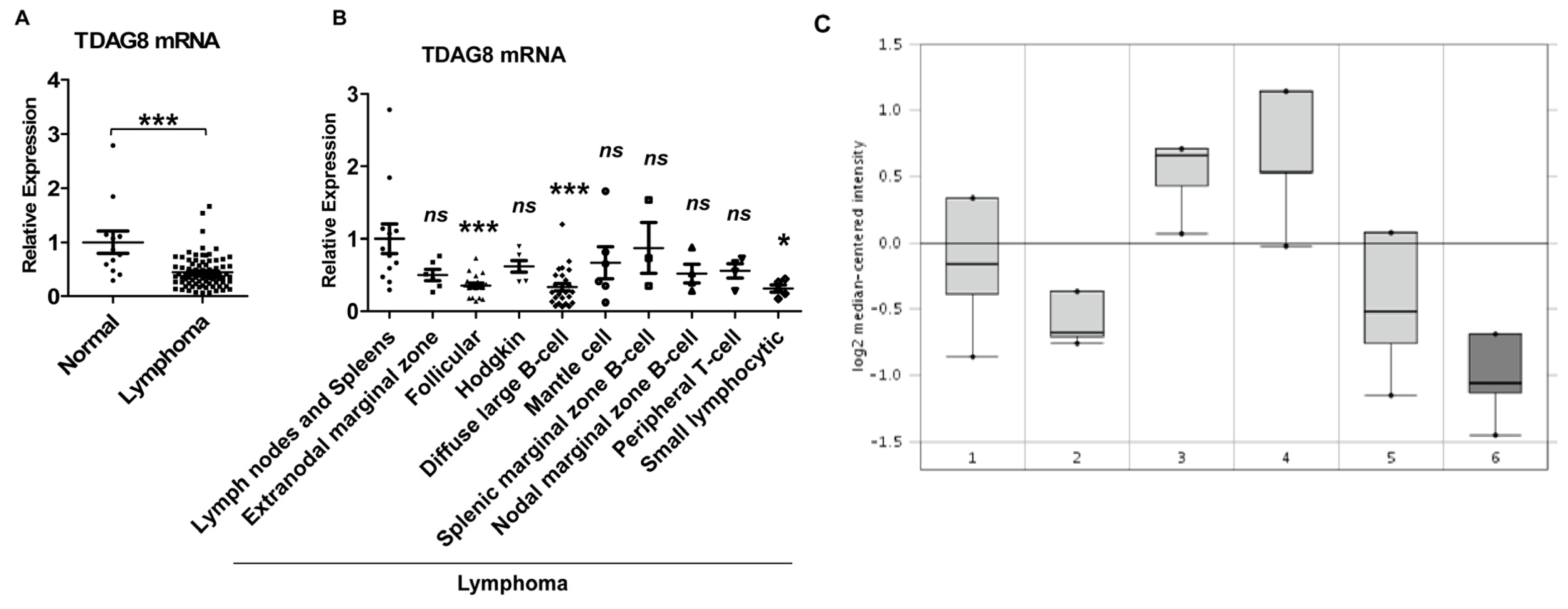
2.6. Expression of TDAG8 Transcripts Is Significantly Reduced in Human Lymphomas
2.7. The Level of TDAG8 mRNA Is Downregulated by Acidosis in U937 Lymphoma Cells
3. Discussion
4. Experimental Section
4.1. Cell Culture and Treatment
4.2. Western Blotting
4.3. RNA Extraction, RT-PCR (Reverse Transcriptase-Polymerase Chain Reaction) and Quantitative Real-Time RT-PCR
4.4. Plasmid Constructs
4.5. Cell Transfection and Transduction
4.6. Oncomine Database Mining
4.7. Statistical Analysis
5. Conclusions










Acknowledgments
Conflicts of Interest
References
- Albihn, A.; Johnsen, J.I.; Henriksson, M.A. MYC in oncogenesis and as a target for cancer therapies. Adv. Cancer Res 2010, 107, 163–224. [Google Scholar]
- Brodeur, G.M.; Seeger, R.C.; Schwab, M.; Varmus, H.E.; Bishop, J.M. Amplification of N-myc in untreated human neuroblastomas correlates with advanced disease stage. Science 1984, 224, 1121–1124. [Google Scholar]
- Nau, M.M.; Brooks, B.J.; Battey, J.; Sausville, E.; Gazdar, A.F.; Kirsch, I.R.; McBride, O.W.; Bertness, V.; Hollis, G.F.; Minna, J.D. l-myc, a new myc-related gene amplified and expressed in human small cell lung cancer. Nature 1985, 318, 69–73. [Google Scholar]
- Dang, C.V. MYC on the path to cancer. Cell 2012, 149, 22–35. [Google Scholar]
- Meyer, N.; Penn, L.Z. Reflecting on 25 years with MYC. Nat. Rev. Cancer 2008, 8, 976–990. [Google Scholar]
- Hurlin, P.J.; Dezfouli, S. Functions of myc:max in the control of cell proliferation and tumorigenesis. Int. Rev. Cytol 2004, 238, 183–226. [Google Scholar]
- Steiner, P.; Philipp, A.; Lukas, J.; Godden-Kent, D.; Pagano, M.; Mittnacht, S.; Bartek, J.; Eilers, M. Identification of a Myc-dependent step during the formation of active G1 cyclin-cdk complexes. EMBO J 1995, 14, 4814–4826. [Google Scholar]
- Zajac-Kaye, M. Myc oncogene: A key component in cell cycle regulation and its implication for lung cancer. Lung Cancer 2001, 34, S43–S46. [Google Scholar]
- Arabi, A.; Wu, S.; Ridderstrale, K.; Bierhoff, H.; Shiue, C.; Fatyol, K.; Fahlen, S.; Hydbring, P.; Soderberg, O.; Grummt, I.; et al. c-Myc associates with ribosomal DNA and activates RNA polymerase I transcription. Nat. Cell Biol 2005, 7, 303–310. [Google Scholar]
- Grandori, C.; Gomez-Roman, N.; Felton-Edkins, Z.A.; Ngouenet, C.; Galloway, D.A.; Eisenman, R.N.; White, R.J. c-Myc binds to human ribosomal DNA and stimulates transcription of rRNA genes by RNA polymerase I. Nat. Cell Biol 2005, 7, 311–318. [Google Scholar]
- Mai, S.; Mushinski, J.F. c-Myc-induced genomic instability. J. Environ. Pathol. Toxicol. Oncol 2003, 22, 179–199. [Google Scholar]
- Schmidt, E.V. The role of c-myc in regulation of translation initiation. Oncogene 2004, 23, 3217–3221. [Google Scholar]
- Wade, M.; Wahl, G.M. c-Myc, genome instability, and tumorigenesis: The devil is in the details. Curr. Top. Microbiol. Immunol 2006, 302, 169–203. [Google Scholar]
- Pelengaris, S.; Khan, M.; Evan, G. c-MYC: More than just a matter of life and death. Nat. Rev. Cancer 2002, 2, 764–776. [Google Scholar]
- Conacci-Sorrell, M.; Ngouenet, C.; Eisenman, R.N. Myc-nick: A cytoplasmic cleavage product of Myc that promotes alpha-tubulin acetylation and cell differentiation. Cell 2010, 142, 480–493. [Google Scholar]
- Hann, S.R. Role of post-translational modifications in regulating c-Myc proteolysis, transcriptional activity and biological function. Semin. Cancer Biol 2006, 16, 288–302. [Google Scholar]
- Wierstra, I.; Alves, J. The c-myc promoter: Still MysterY and challenge. Adv. Cancer Res 2008, 99, 113–333. [Google Scholar]
- Evelhoch, J.L. pH and therapy of human cancers. Novartis Found. Symp 2001, 240, 68–80, discussion 80–84, 152–153. [Google Scholar]
- Mortensen, B.T.; Jensen, P.O.; Helledie, N.; Iversen, P.O.; Ralfkiaer, E.; Larsen, J.K.; Madsen, M.T. Changing bone marrow micro-environment during development of acute myeloid leukaemia in rats. Br. J. Haematol 1998, 102, 458–464. [Google Scholar]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar]
- Yang, L.V.; Castellone, R.D.; Dong, L. Targeting Tumor Microenvironments for Cancer Prevention and Therapy. In Cancer Prevention—From Mechanisms to Translational Benefits; Georgakilas, A.G., Ed.; InTech: Rijeka, Croatia, 2012; pp. 3–40. [Google Scholar]
- Ohtsubo, T.; Wang, X.; Takahashi, A.; Ohnishi, K.; Saito, H.; Song, C.W.; Ohnishi, T. p53-dependent induction of WAF1 by a low-pH culture condition in human glioblastoma cells. Cancer Res 1997, 57, 3910–3913. [Google Scholar]
- Putney, L.K.; Barber, D.L. Na-H exchange-dependent increase in intracellular pH times G2/M entry and transition. J. Biol. Chem 2003, 278, 44645–44649. [Google Scholar]
- Smallbone, K.; Maini, P.K.; Gatenby, R.A. Episodic, transient systemic acidosis delays evolution of the malignant phenotype: Possible mechanism for cancer prevention by increased physical activity. Biol. Direct 2010, 5, 22. [Google Scholar] [Green Version]
- Webb, B.A.; Chimenti, M.; Jacobson, M.P.; Barber, D.L. Dysregulated pH: A perfect storm for cancer progression. Nat. Rev. Cancer 2011, 11, 671–677. [Google Scholar]
- Williams, A.C.; Collard, T.J.; Paraskeva, C. An acidic environment leads to p53 dependent induction of apoptosis in human adenoma and carcinoma cell lines: implications for clonal selection during colorectal carcinogenesis. Oncogene 1999, 18, 3199–3204. [Google Scholar]
- Chen, J.L.; Lucas, J.E.; Schroeder, T.; Mori, S.; Wu, J.; Nevins, J.; Dewhirst, M.; West, M.; Chi, J.T. The genomic analysis of lactic acidosis and acidosis response in human cancers. PLoS Genet 2008, 4, e1000293. [Google Scholar]
- Cairns, R.; Papandreou, I.; Denko, N. Overcoming physiologic barriers to cancer treatment by molecularly targeting the tumor microenvironment. Mol. Cancer Res 2006, 4, 61–70. [Google Scholar]
- Gatenby, R.A.; Gawlinski, E.T.; Gmitro, A.F.; Kaylor, B.; Gillies, R.J. Acid-mediated tumor invasion: A multidisciplinary study. Cancer Res 2006, 66, 5216–5223. [Google Scholar]
- Gatenby, R.A.; Gillies, R.J. Why do cancers have high aerobic glycolysis? Nat. Rev. Cancer 2004, 4, 891–899. [Google Scholar]
- Marino, M.L.; Pellegrini, P.; di Lernia, G.; Djavaheri-Mergny, M.; Brnjic, S.; Zhang, X.; Hagg, M.; Linder, S.; Fais, S.; Codogno, P.; et al. Autophagy is a protective mechanism for human melanoma cells under acidic stress. J. Biol. Chem 2012, 287, 30664–30676. [Google Scholar]
- Wojtkowiak, J.W.; Rothberg, J.M.; Kumar, V.; Schramm, K.J.; Haller, E.; Proemsey, J.B.; Lloyd, M.C.; Sloane, B.F.; Gillies, R.J. Chronic autophagy is a cellular adaptation to tumor acidic pH microenvironments. Cancer Res 2012, 72, 3938–3947. [Google Scholar]
- Bourguignon, L.Y.; Singleton, P.A.; Diedrich, F.; Stern, R.; Gilad, E. CD44 interaction with Na+-H+ exchanger (NHE1) creates acidic microenvironments leading to hyaluronidase-2 and cathepsin B activation and breast tumor cell invasion. J. Biol. Chem 2004, 279, 26991–27007. [Google Scholar]
- Johnson, L.L.; Pavlovsky, A.G.; Johnson, A.R.; Janowicz, J.A.; Man, C.F.; Ortwine, D.F.; Purchase, C.F., II; White, A.D.; Hupe, D.J. A rationalization of the acidic pH dependence for stromelysin-1 (Matrix metalloproteinase-3) catalysis and inhibition. J. Biol. Chem 2000, 275, 11026–11033. [Google Scholar]
- Ishii, S.; Kihara, Y.; Shimizu, T. Identification of T cell death-associated gene 8 (TDAG8) as a novel acid sensing G-protein-coupled receptor. J. Biol. Chem 2005, 280, 9083–9087. [Google Scholar]
- Liu, J.P.; Nakakura, T.; Tomura, H.; Tobo, M.; Mogi, C.; Wang, J.Q.; He, X.D.; Takano, M.; Damirin, A.; Komachi, M.; et al. Each one of certain histidine residues in G-protein-coupled receptor GPR4 is critical for extracellular proton-induced stimulation of multiple G-protein-signaling pathways. Pharmacol. Res 2010, 61, 499–505. [Google Scholar]
- Ludwig, M.G.; Vanek, M.; Guerini, D.; Gasser, J.A.; Jones, C.E.; Junker, U.; Hofstetter, H.; Wolf, R.M.; Seuwen, K. Proton-sensing G-protein-coupled receptors. Nature 2003, 425, 93–98. [Google Scholar]
- Murakami, N.; Yokomizo, T.; Okuno, T.; Shimizu, T. G2A is a proton-sensing G-protein-coupled receptor antagonized by lysophosphatidylcholine. J. Biol. Chem 2004, 279, 42484–42491. [Google Scholar]
- Radu, C.G.; Nijagal, A.; McLaughlin, J.; Wang, L.; Witte, O.N. Differential proton sensitivity of related G protein-coupled receptors T cell death-associated gene 8 and G2A expressed in immune cells. Proc. Natl. Acad. Sci. USA 2005, 102, 1632–1637. [Google Scholar]
- Tobo, M.; Tomura, H.; Mogi, C.; Wang, J.Q.; Liu, J.P.; Komachi, M.; Damirin, A.; Kimura, T.; Murata, N.; Kurose, H.; et al. Previously postulated “ligand-independent” signaling of GPR4 is mediated through proton-sensing mechanisms. Cell Signal 2007, 19, 1745–1753. [Google Scholar]
- Wang, J.Q.; Kon, J.; Mogi, C.; Tobo, M.; Damirin, A.; Sato, K.; Komachi, M.; Malchinkhuu, E.; Murata, N.; Kimura, T.; et al. TDAG8 is a proton-sensing and psychosine-sensitive G-protein-coupled receptor. J. Biol. Chem 2004, 279, 45626–45633. [Google Scholar]
- Yang, L.V.; Radu, C.G.; Roy, M.; Lee, S.; McLaughlin, J.; Teitell, M.A.; Iruela-Arispe, M.L.; Witte, O.N. Vascular abnormalities in mice deficient for the G protein-coupled receptor GPR4 that functions as a pH sensor. Mol. Cell Biol 2007, 27, 1334–1347. [Google Scholar]
- Choi, J.W.; Lee, S.Y.; Choi, Y. Identification of a putative G protein-coupled receptor induced during activation-induced apoptosis of T cells. Cell. Immunol 1996, 168, 78–84. [Google Scholar]
- Kyaw, H.; Zeng, Z.; Su, K.; Fan, P.; Shell, B.K.; Carter, K.C.; Li, Y. Cloning, characterization, and mapping of human homolog of mouse T-cell death-associated gene. DNA Cell Biol 1998, 17, 493–500. [Google Scholar]
- Malone, M.H.; Wang, Z.; Distelhorst, C.W. The glucocorticoid-induced gene tdag8 encodes a pro-apoptotic G protein-coupled receptor whose activation promotes glucocorticoid-induced apoptosis. J. Biol. Chem 2004, 279, 52850–52859. [Google Scholar]
- Radu, C.G.; Cheng, D.; Nijagal, A.; Riedinger, M.; McLaughlin, J.; Yang, L.V.; Johnson, J.; Witte, O.N. Normal immune development and glucocorticoid-induced thymocyte apoptosis in mice deficient for the T-cell death-associated gene 8 receptor. Mol. Cell. Biol 2006, 26, 668–677. [Google Scholar]
- Tosa, N.; Murakami, M.; Jia, W.Y.; Yokoyama, M.; Masunaga, T.; Iwabuchi, C.; Inobe, M.; Iwabuchi, K.; Miyazaki, T.; Onoe, K.; et al. Critical function of T cell death-associated gene 8 in glucocorticoid-induced thymocyte apoptosis. Int. Immunol 2003, 15, 741–749. [Google Scholar]
- Ihara, Y.; Kihara, Y.; Hamano, F.; Yanagida, K.; Morishita, Y.; Kunita, A.; Yamori, T.; Fukayama, M.; Aburatani, H.; Shimizu, T.; et al. The G protein-coupled receptor T-cell death-associated gene 8 (TDAG8) facilitates tumor development by serving as an extracellular pH sensor. Proc. Natl. Acad. Sci. USA 2010, 107, 17309–17314. [Google Scholar]
- Ryder, C.; McColl, K.; Zhong, F.; Distelhorst, C.W. Acidosis promotes Bcl-2 family-mediated evasion of apoptosis: Involvement of acid-sensing G protein-coupled receptor Gpr65 signaling to Mek/Erk. J. Biol. Chem 2012, 287, 27863–27875. [Google Scholar]
- Weng, Z.; Fluckiger, A.C.; Nisitani, S.; Wahl, M.I.; Le, L.Q.; Hunter, C.A.; Fernal, A.A.; le Beau, M.M.; Witte, O.N. A DNA damage and stress inducible G protein-coupled receptor blocks cells in G2/M. Proc. Natl. Acad. Sci. USA 1998, 95, 12334–12339. [Google Scholar]
- Fukumura, D.; Jain, R.K. Tumor microvasculature and microenvironment: targets for anti-angiogenesis and normalization. Microvasc. Res 2007, 74, 72–84. [Google Scholar]
- Ansell, S.M.; Armitage, J.O. Positron emission tomographic scans in lymphoma: Convention and controversy. Mayo Clin. Proc 2012, 87, 571–580. [Google Scholar]
- Buerkle, A.; Weber, W.A. Imaging of tumor glucose utilization with positron emission tomography. Cancer Metastasis. Rev 2008, 27, 545–554. [Google Scholar]
- Elhomsy, G.C.; Eranki, V.; Albert, S.G.; Fesler, M.J.; Parker, S.M.; Michael, A.G.; Griffing, G.T. Hyper-warburgism a cause of asymptomatic hypoglycemia with lactic acidosis in a patient with non-Hodgkin’s lymphoma. J. Clin. Endocrinol. Metab 2012, 97, 4311–4316. [Google Scholar]
- Ruiz, J.P.; Singh, A.K.; Hart, P. Type B lactic acidosis secondary to malignancy: case report, review of published cases, insights into pathogenesis, and prospects for therapy. ScientificWorldJournal 2011, 11, 1316–1324. [Google Scholar]
- Zanke, B.W.; Lee, C.; Arab, S.; Tannock, I.F. Death of tumor cells after intracellular acidification is dependent on stress-activated protein kinases (SAPK/JNK) pathway activation and cannot be inhibited by Bcl-2 expression or interleukin 1beta-converting enzyme inhibition. Cancer Res 1998, 58, 2801–2808. [Google Scholar]
- Morita, T. Low pH leads to sister-chromatid exchanges and chromosomal aberrations, and its clastogenicity is S-dependent. Mutat. Res 1995, 334, 301–308. [Google Scholar]
- Sin, W.C.; Zhang, Y.; Zhong, W.; Adhikarakunnathu, S.; Powers, S.; Hoey, T.; An, S.; Yang, J. G protein-coupled receptors GPR4 and TDAG8 are oncogenic and overexpressed in human cancers. Oncogene 2004, 23, 6299–6303. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar]
- He, X.D.; Tobo, M.; Mogi, C.; Nakakura, T.; Komachi, M.; Murata, N.; Takano, M.; Tomura, H.; Sato, K.; Okajima, F. Involvement of proton-sensing receptor TDAG8 in the anti-inflammatory actions of dexamethasone in peritoneal macrophages. Biochem. Biophys. Res. Commun 2011, 415, 627–631. [Google Scholar]
- Guo, J.; Parise, R.A.; Joseph, E.; Egorin, M.J.; Lazo, J.S.; Prochownik, E.V.; Eiseman, J.L. Efficacy, pharmacokinetics, tisssue distribution, and metabolism of the Myc-Max disruptor, 10058-F4 [Z,E]-5-[4-ethylbenzylidine]-2-thioxothiazolidin-4-one, in mice. Cancer Chemother. Pharmacol 2009, 63, 615–625. [Google Scholar]
- Lin, C.P.; Liu, J.D.; Chow, J.M.; Liu, C.R.; Liu, H.E. Small-molecule c-Myc inhibitor, 10058-F4, inhibits proliferation, downregulates human telomerase reverse transcriptase and enhances chemosensitivity in human hepatocellular carcinoma cells. Anticancer Drugs 2007, 18, 161–170. [Google Scholar]
- Yin, X.; Giap, C.; Lazo, J.S.; Prochownik, E.V. Low molecular weight inhibitors of Myc-Max interaction and function. Oncogene 2003, 22, 6151–6159. [Google Scholar]
- Delmore, J.E.; Issa, G.C.; Lemieux, M.E.; Rahl, P.B.; Shi, J.; Jacobs, H.M.; Kastritis, E.; Gilpatrick, T.; Paranal, R.M.; Qi, J.; et al. BET bromodomain inhibition as a therapeutic strategy to target c-Myc. Cell 2011, 146, 904–917. [Google Scholar]
- Mertz, J.A.; Conery, A.R.; Bryant, B.M.; Sandy, P.; Balasubramanian, S.; Mele, D.A.; Bergeron, L.; Sims, R.J., III. Targeting MYC dependence in cancer by inhibiting BET bromodomains. Proc. Natl. Acad. Sci. USA 2011, 108, 16669–16674. [Google Scholar]
- Zuber, J.; Shi, J.; Wang, E.; Rappaport, A.R.; Herrmann, H.; Sison, E.A.; Magoon, D.; Qi, J.; Blatt, K.; Wunderlich, M.; et al. RNAi screen identifies Brd4 as a therapeutic target in acute myeloid leukaemia. Nature 2011, 478, 524–528. [Google Scholar]
- Lundstrom, K. Latest development in drug discovery on G protein-coupled receptors. Curr. Protein Pept. Sci 2006, 7, 465–470. [Google Scholar]
- Onozawa, Y.; Fujita, Y.; Kuwabara, H.; Nagasaki, M.; Komai, T.; Oda, T. Activation of T cell death-associated gene 8 regulates the cytokine production of T cells and macrophages in vitro. Eur. J. Pharmacol. 2012, 683, 325–331. [Google Scholar]
- Li, Z.; Leffler, N.R.; Yang, L.V. Department of Oncology, East Carolina University; Unpublished data; 2013.
- Castellone, R.D.; Leffler, N.R.; Dong, L.; Yang, L.V. Inhibition of tumor cell migration and metastasis by the proton-sensing GPR4 receptor. Cancer Lett 2011, 312, 197–208. [Google Scholar]
- Chen, A.; Dong, L.; Leffler, N.R.; Asch, A.S.; Witte, O.N.; Yang, L.V. Activation of GPR4 by acidosis increases endothelial cell adhesion through the cAMP/Epac pathway. PLoS One 2011, 6, e27586. [Google Scholar]
- Dong, L.; Li, Z.; Leffler, N.R.; Asch, A.S.; Chi, J.T.; Yang, L.V. Acidosis activation of the proton-sensing GPR4 receptor stimulates vascular endothelial cell inflammatory responses revealed by transcriptome analysis. PLoS One 2013, 8, e61991. [Google Scholar]
- Zhang, Y.; Feng, Y.; Justus, C.R.; Jiang, W.; Li, Z.; Lu, J.Q.; Brock, R.S.; McPeek, M.K.; Weidner, D.A.; Yang, L.V.; et al. Comparative study of 3D morphology and functions on genetically engineered mouse melanoma cells. Integr. Biol. (Camb.) 2012, 4, 1428–1436. [Google Scholar]
- Yang, L.V.; Radu, C.G.; Wang, L.; Riedinger, M.; Witte, O.N. Gi-independent macrophage chemotaxis to lysophosphatidylcholine via the immunoregulatory GPCR G2A. Blood 2005, 105, 1127–1134. [Google Scholar]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Z.; Dong, L.; Dean, E.; Yang, L.V. Acidosis Decreases c-Myc Oncogene Expression in Human Lymphoma Cells: A Role for the Proton-Sensing G Protein-Coupled Receptor TDAG8. Int. J. Mol. Sci. 2013, 14, 20236-20255. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms141020236
Li Z, Dong L, Dean E, Yang LV. Acidosis Decreases c-Myc Oncogene Expression in Human Lymphoma Cells: A Role for the Proton-Sensing G Protein-Coupled Receptor TDAG8. International Journal of Molecular Sciences. 2013; 14(10):20236-20255. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms141020236
Chicago/Turabian StyleLi, Zhigang, Lixue Dong, Eric Dean, and Li V. Yang. 2013. "Acidosis Decreases c-Myc Oncogene Expression in Human Lymphoma Cells: A Role for the Proton-Sensing G Protein-Coupled Receptor TDAG8" International Journal of Molecular Sciences 14, no. 10: 20236-20255. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms141020236