Effect of Melatonin on Human Dental Papilla Cells

Abstract
:1. Introduction
2. Results
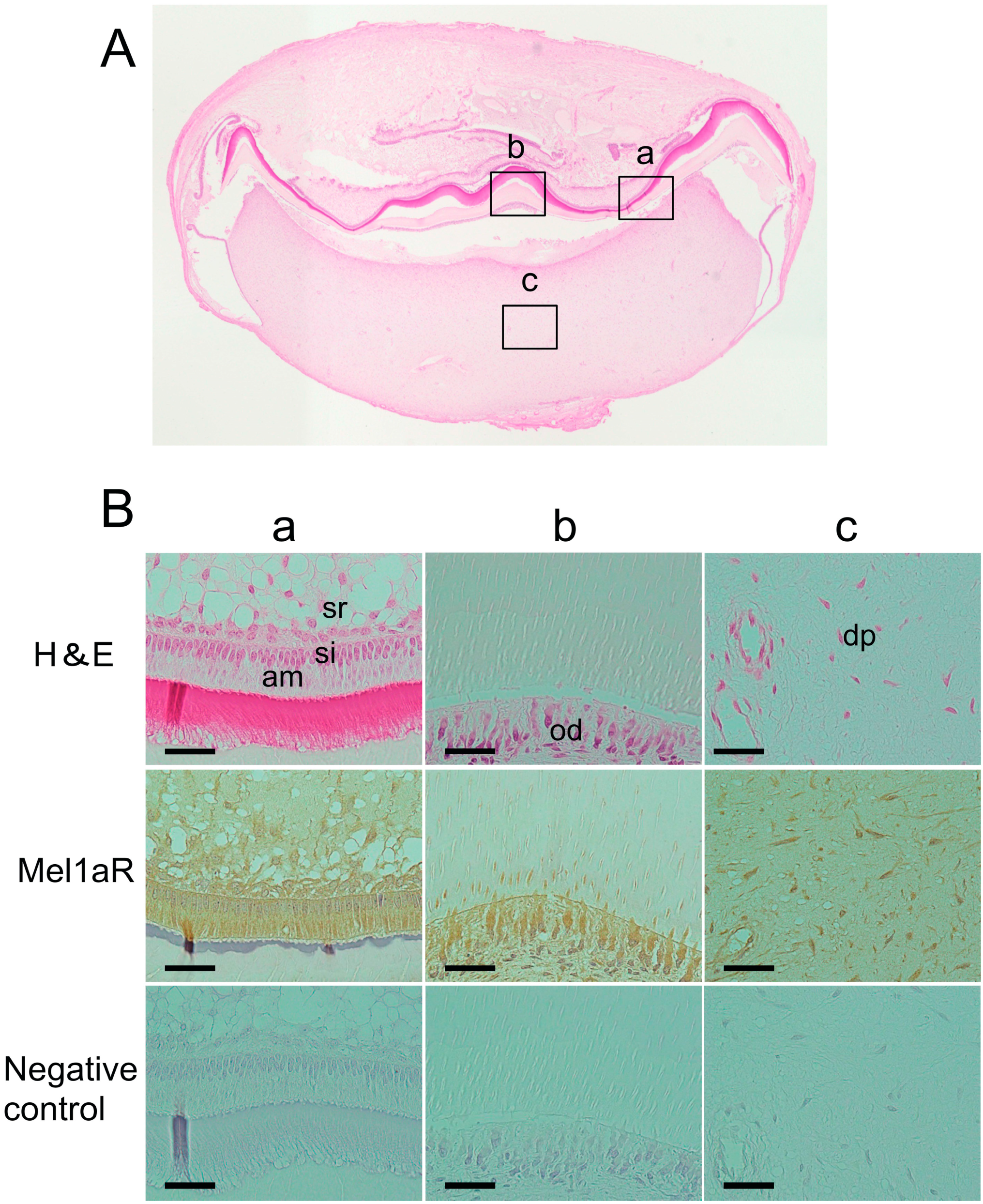
2.1. Localization of Melatonin 1a Receptor (Mel1aR) in Human Tooth Germs

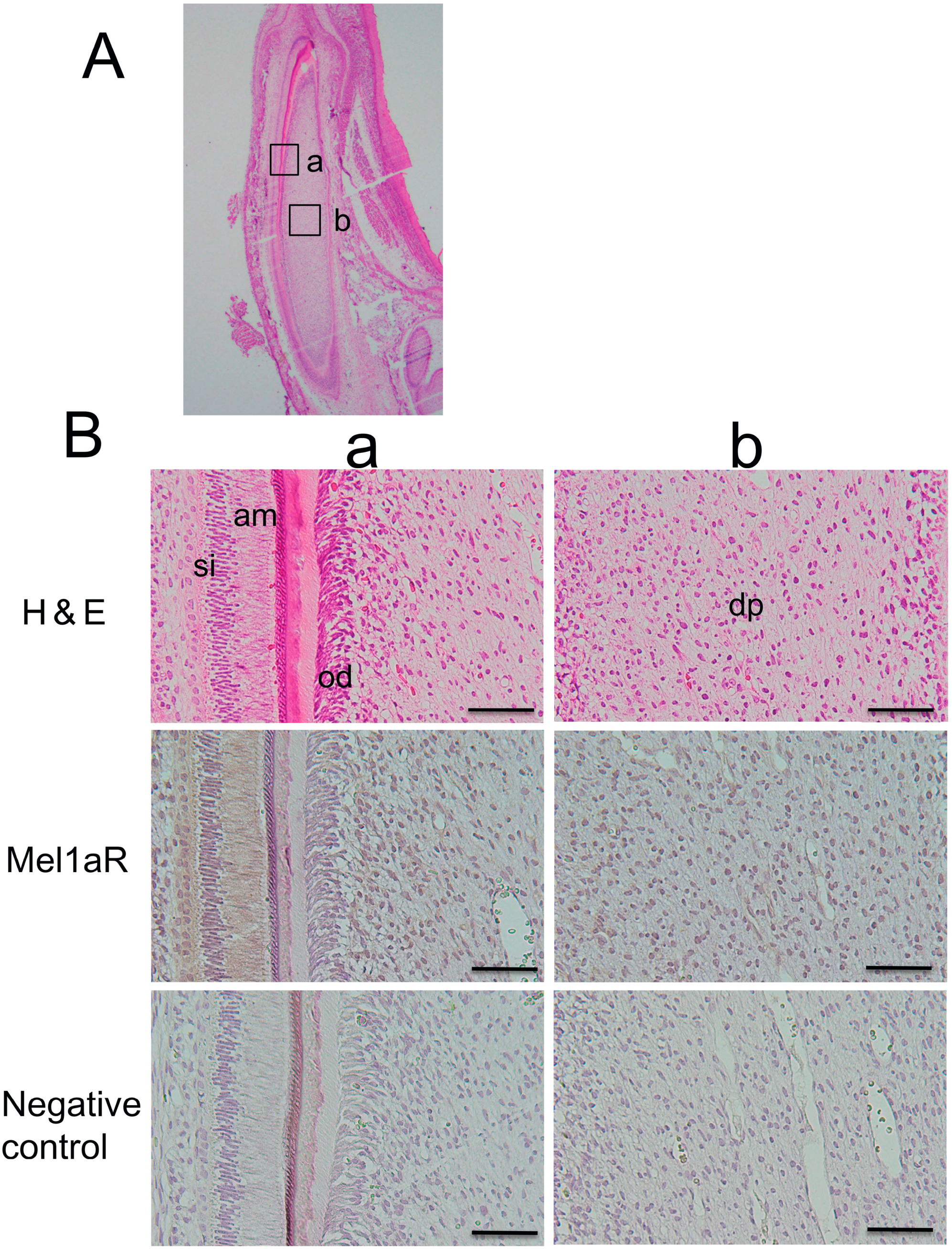
2.2. Localization of Mel1aR in Rat Tooth Germs

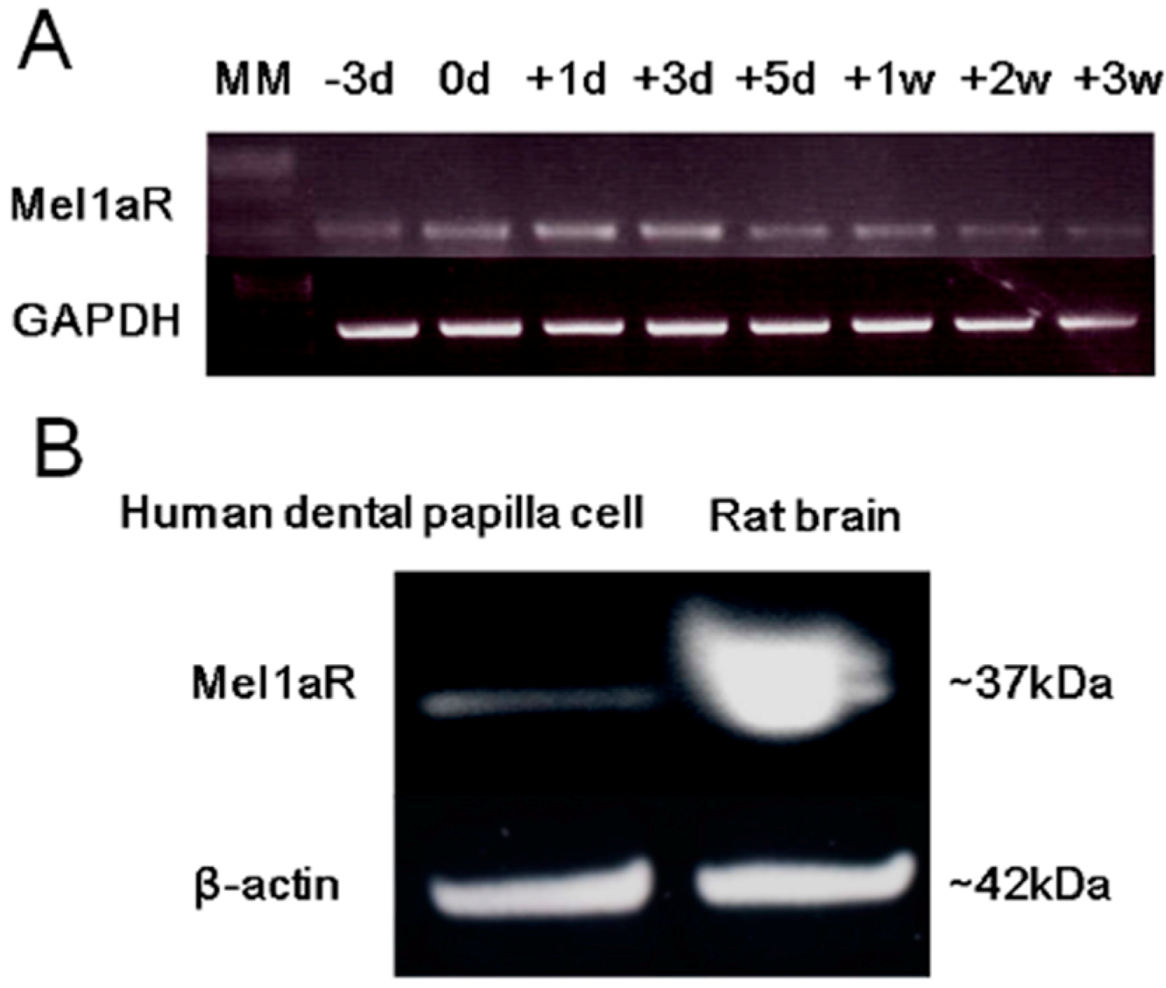
2.3. Expression of Mel1aR in Human Dental Papilla Cells

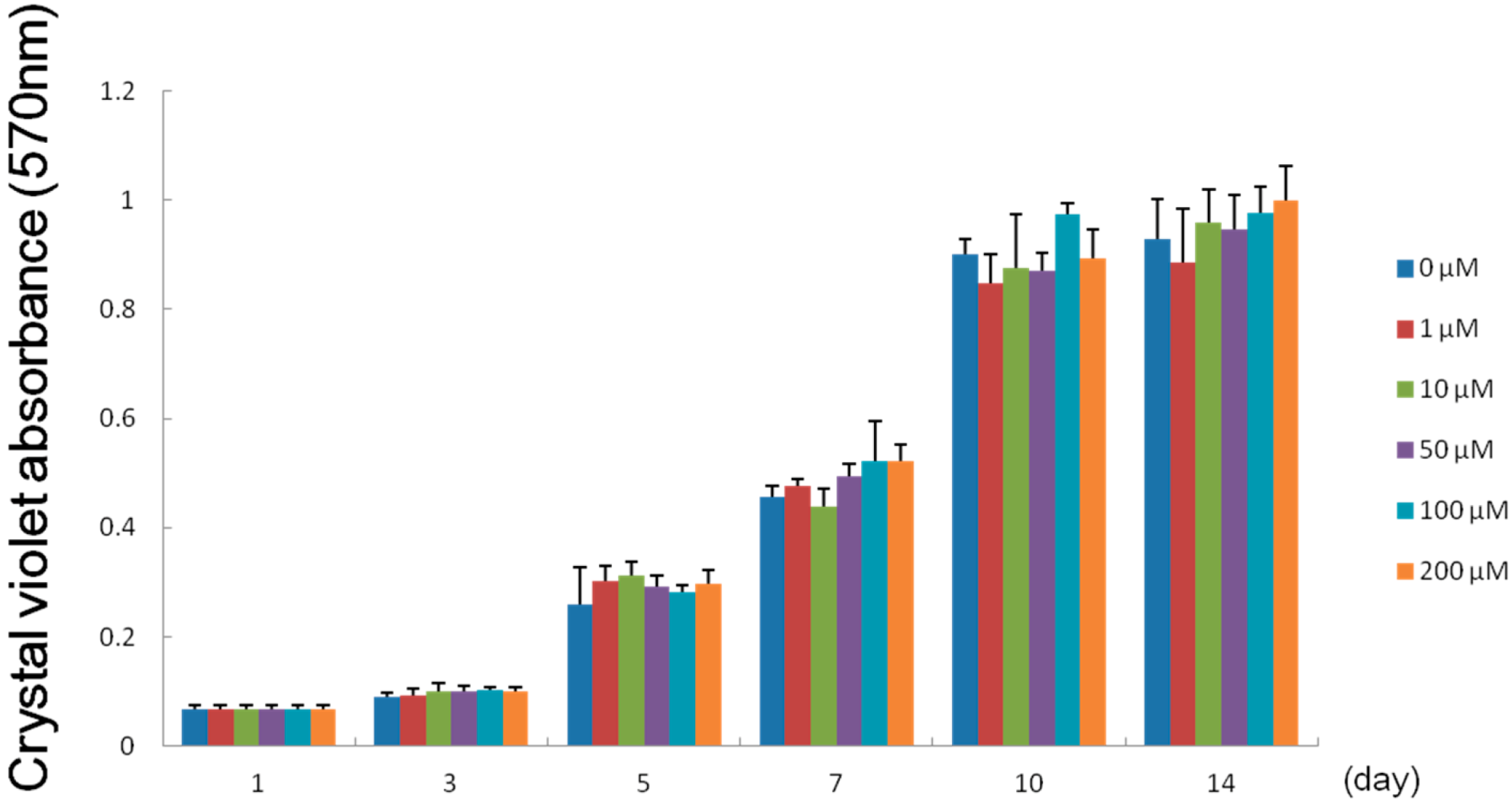
2.4. Effect of Melatonin on the Proliferation of Human Dental Papilla Cells
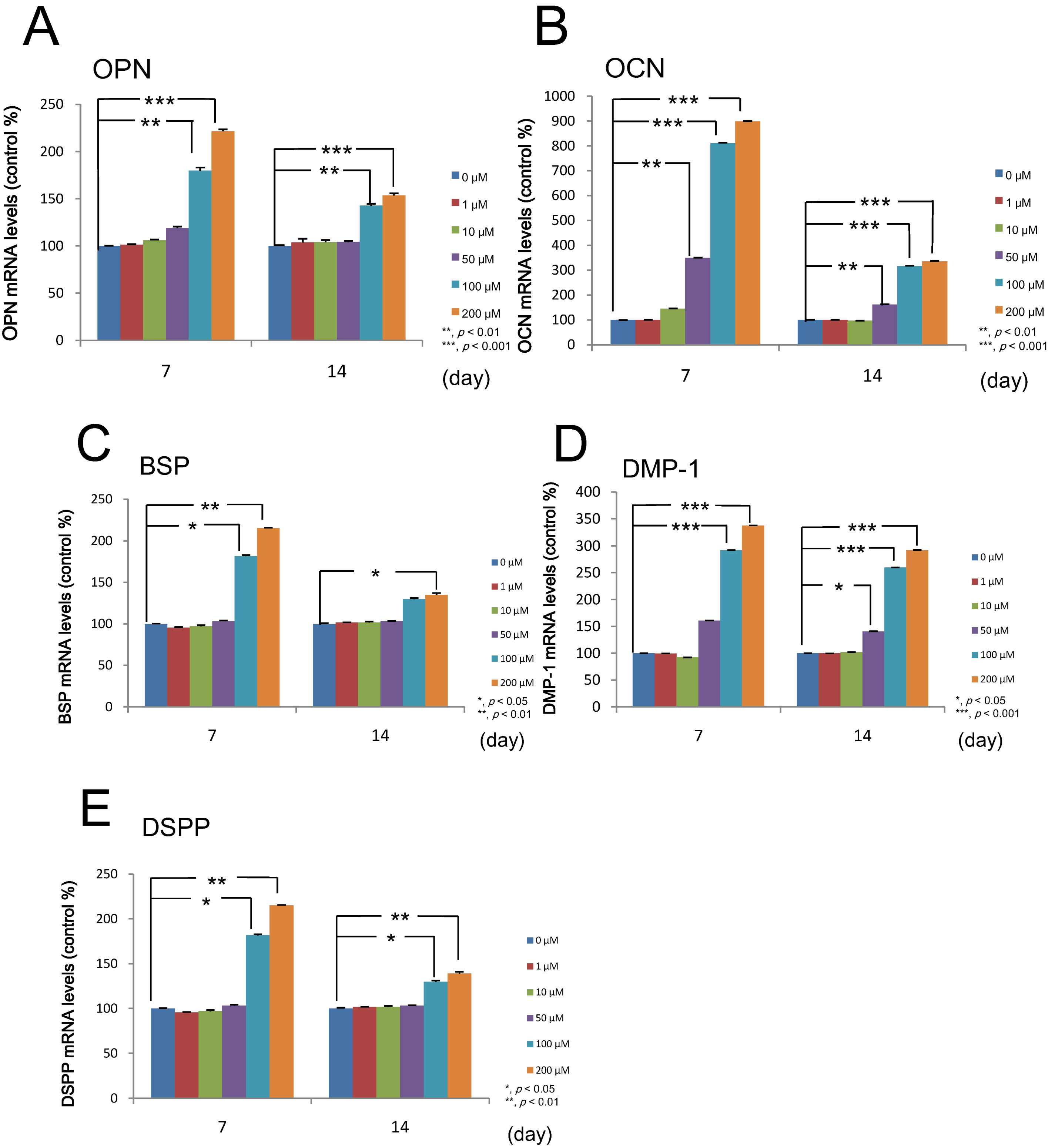
2.5. Effect of Melatonin on the Differentiation of Human Dental Papilla Cells
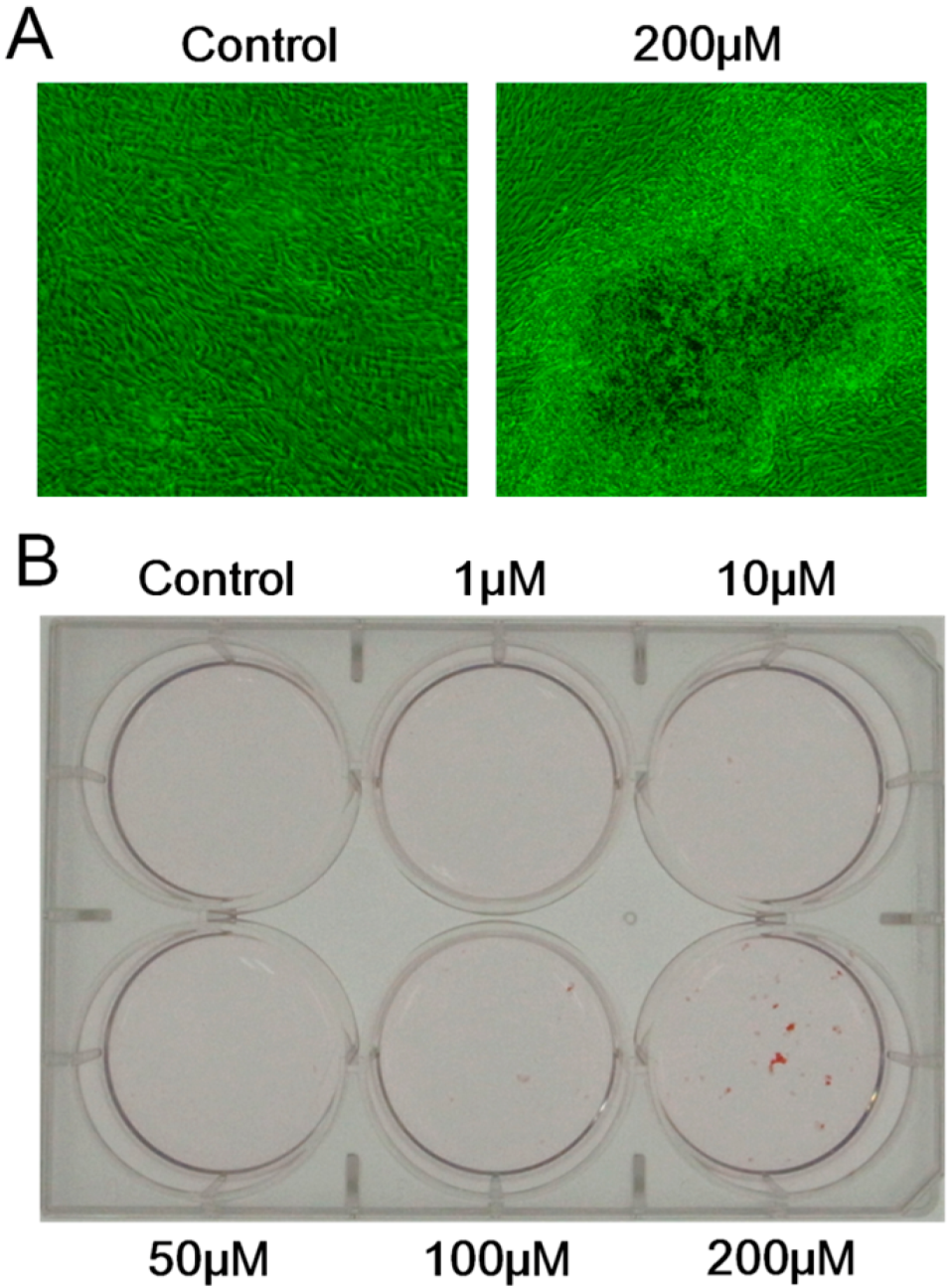
2.6. Effect of Melatonin on the Mineralized Matrix Formation by Human Dental Papilla Cells



3. Discussion
4. Experimental Section
4.1. Human Tissue Samples
4.2. Rat Tissue Samples
4.3. Immunohistochemistry
4.4. Cell Culture
4.5. Semi-Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer (GenBank Accession Number) | Sequences | Size (bp) | Annealing Temperature | Cycles |
|---|---|---|---|---|
| human Mel1aR | F: 5'-GATCCTGGTTGTCCAGGTCA-3' | 241 | 60 | 35 |
| (NM_005958) | R: 5'-CATTGAGGCAGCTGTTGAAA-3' | |||
| GAPDH | F: 5'-ACCACAGTCCATGCCATCAC-3' | 451 | 56 | 23 |
| (NM_001289746) | R: 5'-TCCACCACCCTGTTGCTGTA-3' |
4.6. Western Blot Analysis
4.7. Cell Proliferation Assay
4.8. Real-Time Reverse Transcription-Polymerase Chain Reaction
| Primers (GenBank Accession Number) | Sequences | Size (bp) |
|---|---|---|
| OPN | F: 5'-TGAAACGAGTCAGCTGGATG-3' | 162 |
| (NM_001251830) | R: 5'-TGAAATTCATGGCTGTGGAA-3' | |
| OCN | F: 5'-GTGCAGAGTCCAGCAAAGGT-3' | 175 |
| (NM_001199662) | R: 5'-TCAGCCAACTCGTCACAGTC-3' | |
| BSP | F: 5'-CAACAGCACAGAGGCAGAAA-3' | 247 |
| (NM_004967) | R: 5'-CGTACTCCCCCTCGTATTCA-3' | |
| DMP-1 | F: 5'-CAGGAGCACAGGAAAAGGAG-3' | 212 |
| (NM_004407) | R: 5'-CTGGTGGTATCTTGGGCACT-3' | |
| DSPP | F: 5'-TCACAAGGGAGAAGGGAATG-3' | 181 |
| (NM_014208) | R: 5'-TGCCATTTGCTGTGATGTTT-3' | |
| GAPDH | F: 5'-GAGTCAACGGATTTGGTCGT-3' | 261 |
| (NM_001289746) | R: 5'-TTGATTTTGGAGGGATCTCG-3' |
4.9. Alizarin Red S Staining
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocytes. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar]
- Reiter, R.J.; Oh, C.S.; Fujimori, O. Melatonin its intracellular and genomic actions. Trends. Endcrinol. Metab. 1996, 7, 22–27. [Google Scholar]
- Stehle, J.H.; Saade, A.; Rawashdeho, O.; Ackermann, K.; Jilg, A.; Sebestény, T.; Maronde, E. A survey of molecular details in the human pineal gland in the light of phylogeny, structure, function and chronobiological diseases. J. Pineal Res. 2011, 51, 17–43. [Google Scholar]
- Hamm, H.E.; Menaker, M. Retinal rhythms in chicks: Circadian variation in melatonin and serotonin N-acetyltransferase activity. Proc. Natl. Acad. Sci. USA 1980, 77, 4998–5002. [Google Scholar]
- Itoh, M.T.; Ishizuka, B.; Kuribayashi, Y.; Amemiya, A.; Sumi, Y. Melatonin, its precursors, and synthesizing enzyme activities in the human ovary. Mol. Hum. Reprod. 1999, 5, 402–408. [Google Scholar]
- Itoh, M.T.; Takahashi, N.; Abe, M.; Shimizu, K. Expression and cellular localization of melatonin-synthesizing enzymes in the rat lens. J. Pineal Res. 2007, 42, 92–96. [Google Scholar]
- Stefuji, J.; Hörtner, M.; Ghosh, M.; Schauenstein, K.; Rinner, I.; Wölfler, A.; Semmler, J.; Liebmann, P.M. Gene expression of the key enzymes of melatonin synthesis in extrapineal tissues of the rat. J. Pineal Res. 2001, 30, 243–247. [Google Scholar]
- Shimozuma, M.; Tokuyama, R.; Tatehara, S.; Umeki, H.; Ide, S.; Mishima, K.; Saito, I.; Satomura, K. Expression and cellular localization of melatonin-synthesizing enzymes in rat and human salivary glands. Histochem. Cell Biol. 2011, 135, 389–396. [Google Scholar]
- McArthur, A.J.; Hunt, A.E.; Gillette, M.U. Melatonin action and signal transduction in the rat suprachiasmatic circadian clock: Activation of protein kinase C at dusk and dawn. Endocrinology 1997, 138, 627–634. [Google Scholar]
- Dollins, A.B.; Zhdanova, I.V.; Wurtman, R.J.; Lynch, H.J.; Deng, M.H. Effect of inducing nocturnal serum melatonin concentrations in daytime on sleep, mood, body temperature, and performance. Proc. Natl. Acad. Sci. USA 1994, 91, 1824–1828. [Google Scholar]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Sanchez-Sanchez, J.J.; Kaski, J.C.; Reiter, R.J. Melatonin and circadian biology in human cardiovascular disease. J. Pineal Res. 2010, 49, 14–22. [Google Scholar]
- Kennaway, D.J.; Rowe, S.A. Melatonin binding sites and their role in seasonal reproduction. J. Reprod. Fertil. Suppl. 1995, 49, 423–435. [Google Scholar]
- Raghavendra, V.; Singh, V.; Kulkarni, S.K.; Agrewala, J.N. Melatonin enhances Th2 cell mediated immune responses: Lack of sensitivity to reversal by naltrexone or benzodiazepine receptor antagonists. Mol. Cell. Biochem. 2001, 221, 57–62. [Google Scholar]
- Guerrero, J.M.; Reiter, R.J. Melatonin-immune system relationships. Curr. Top. Med. Chem. 2002, 2, 167–179. [Google Scholar]
- León, J.; Acuña-Castroviejo, D.; Escames, D.; Tan, D.X.; Reiter, R.J. Melatonin mitigates mitochondrial malfunction. J. Pineal Res. 2005, 38, 1–9. [Google Scholar]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Sánchez-Barceló, E.J.; Cos, S.; Mediavilla, M.D.; Martínez-Campa, C.; González, A.; Alonso-González, C. Melatonin-estrogen interaction in breast cancer. J. Pineal Res. 2005, 38, 217–222. [Google Scholar]
- Martínez-Campa, C.; Alonso-González, C.; Mediavilla, M.D.; Cos, S.; González, A.; Ramos, S.; Sánchez-Barceló, E.J. Melatonin inhibits both ER activation and breast cancer cell proliferation induced by a metalloestrogen, cadmium. J. Pineal Res. 2006, 40, 291–294. [Google Scholar] [CrossRef]
- Jardim-Perassi, B.V.; Arbab, A.S.; Ferreira, L.C. Effect of melatonin on tumor growth and angiogenesis in xenograft model of breast cancer. PLoS One 2014, 9, e85311. [Google Scholar]
- Czesnikiewicz-Guzik, M.; Konturek, S.J.; Loster, B.; Wisniewska, G.; Majewski, S. Melatonin and its role in oxidative stress related diseases of oral cavity. J. Physiol. Pharmacol. 2007, 58, 5–19. [Google Scholar]
- Cutando, A.; Gómez-Moreno, G.; Arana, C.; Acuña-Castroviejo, D.; Reiter, R.J. Melatonin: Potential functions in the oral cavity. J. Periodontol. 2007, 78, 1094–1102. [Google Scholar]
- Gómez-Moreno, G.; Guardia, J.; Ferrera, M.J.; Cutando, A.; Reiter, R.J. Melatonin in diseases of the oral cavity. Oral Dis. 2010, 16, 242–247. [Google Scholar] [CrossRef]
- Cutando, A.; Aneiros-Fernández, J.; Aneiros-Cachaza, J.; Arias-Santiago, S. Melatonin and cancer: Current knowledge and its application to oral cavity tumours. J. Oral Pathol. Med. 2011, 40, 593–597. [Google Scholar]
- Roth, J.A.; Kim, B.G.; Lin, W.L.; Cho, M.I. Melatonin promotes osteoblast differentiation and bone formation. J. Biol. Chem. 1999, 274, 22041–22047. [Google Scholar] [CrossRef]
- Nakade, O.; Koyama, H.; Ariji, H.; Yajima, A.; Kaku, T. Melatonin stimulates proliferation and type I collagen synthesis in human bone cells in vitro. J. Pineal Res. 1999, 27, 106–110. [Google Scholar] [CrossRef]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via MT2 melatonin receptors and the MEK/ERK (1/2) signaling cascade. J. Pineal Res. 2006, 40, 332–342. [Google Scholar] [CrossRef]
- Satomura, K.; Tobiume, S.; Tokuyama, R.; Yamasaki, Y.; Kudoh, K.; Maeda, E.; Nagayama, M. Melatonin at pharmacological doses enhances human osteoblastic differentiation in vitro and promotes mouse cortical bone formation in vivo. J. Pineal Res. 2007, 42, 231–239. [Google Scholar] [CrossRef]
- Koyama, H.; Nakade, O.; Takada, Y.; Kaku, T.; Lau, K.H. Melatonin at pharmacologic doses increases bone mass by suppressing resorption through down-regulation of the RANKL-mediated osteoclast formation and activation. J. Bone Miner. Res. 2002, 17, 1219–1229. [Google Scholar] [CrossRef]
- Suzuki, N.; Hattori, A. Melatonin suppresses osteoclastic and osteoblastic activities in the scales of goldfish. J. Pineal Res. 2002, 33, 253–258. [Google Scholar] [CrossRef]
- Ladizesky, M.G.; Boggio, V.; Albornoz, L.E.; Castrillon, P.O.; Mautalen, C.; Cardinali, D.P. Melatonin increases oestradiol-induced bone formation in ovariectomized rats. J. Pineal Res. 2003, 34, 143–151. [Google Scholar] [CrossRef]
- Kumasaka, S.; Shimozuma, M.; Kawamoto, T.; Mishima, K.; Tokuyama, R.; Kamiya, Y.; Davaadorj, P.; Saito, I.; Satomura, K. Possible involvement of melatonin in tooth development: Expression of melatonin 1a receptor in human and mouse tooth germs. Histochem. Cell Biol. 2010, 133, 577–584. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, H.; Fan, W.; Dong, W.; Fu, S.; He, H.; Huang, F. Melatonin influences proliferation and differentiation of rat dental papilla cells in vitro and dentine formation in vivo by altering mitochondrial activity. J. Pineal Res. 2013, 54, 170–178. [Google Scholar] [CrossRef]
- Tamura, H.; Nakamura, Y.; Terron, M.P.; Flores, L.J.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and pregnancy in the human. Reprod. Toxicol. 2008, 25, 291–303. [Google Scholar]
- Karasek, H. Melatonin play a role in aging processes? J. Physiol. Pharmacol. 2007, 58, 105–113. [Google Scholar]
- Karasek, H.; Winczyk, K. Melatonin in humans. J. Physiol. Pharmacol. 2006, 57, 19–39. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tachibana, R.; Tatehara, S.; Kumasaka, S.; Tokuyama, R.; Satomura, K. Effect of Melatonin on Human Dental Papilla Cells. Int. J. Mol. Sci. 2014, 15, 17304-17317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151017304
Tachibana R, Tatehara S, Kumasaka S, Tokuyama R, Satomura K. Effect of Melatonin on Human Dental Papilla Cells. International Journal of Molecular Sciences. 2014; 15(10):17304-17317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151017304
Chicago/Turabian StyleTachibana, Ryusuke, Seiko Tatehara, Shuku Kumasaka, Reiko Tokuyama, and Kazuhito Satomura. 2014. "Effect of Melatonin on Human Dental Papilla Cells" International Journal of Molecular Sciences 15, no. 10: 17304-17317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151017304