
Fenarimol, a Pyrimidine-Type Fungicide, Inhibits Brassinosteroid Biosynthesis

Abstract
:
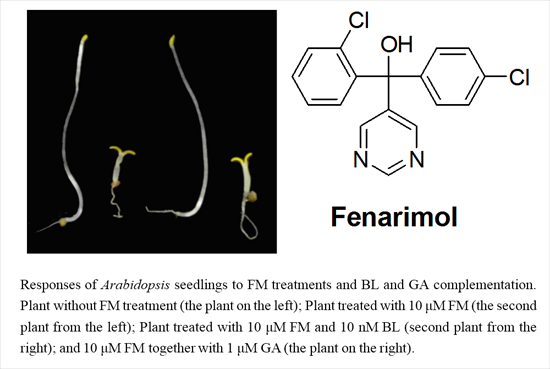{kind=link}
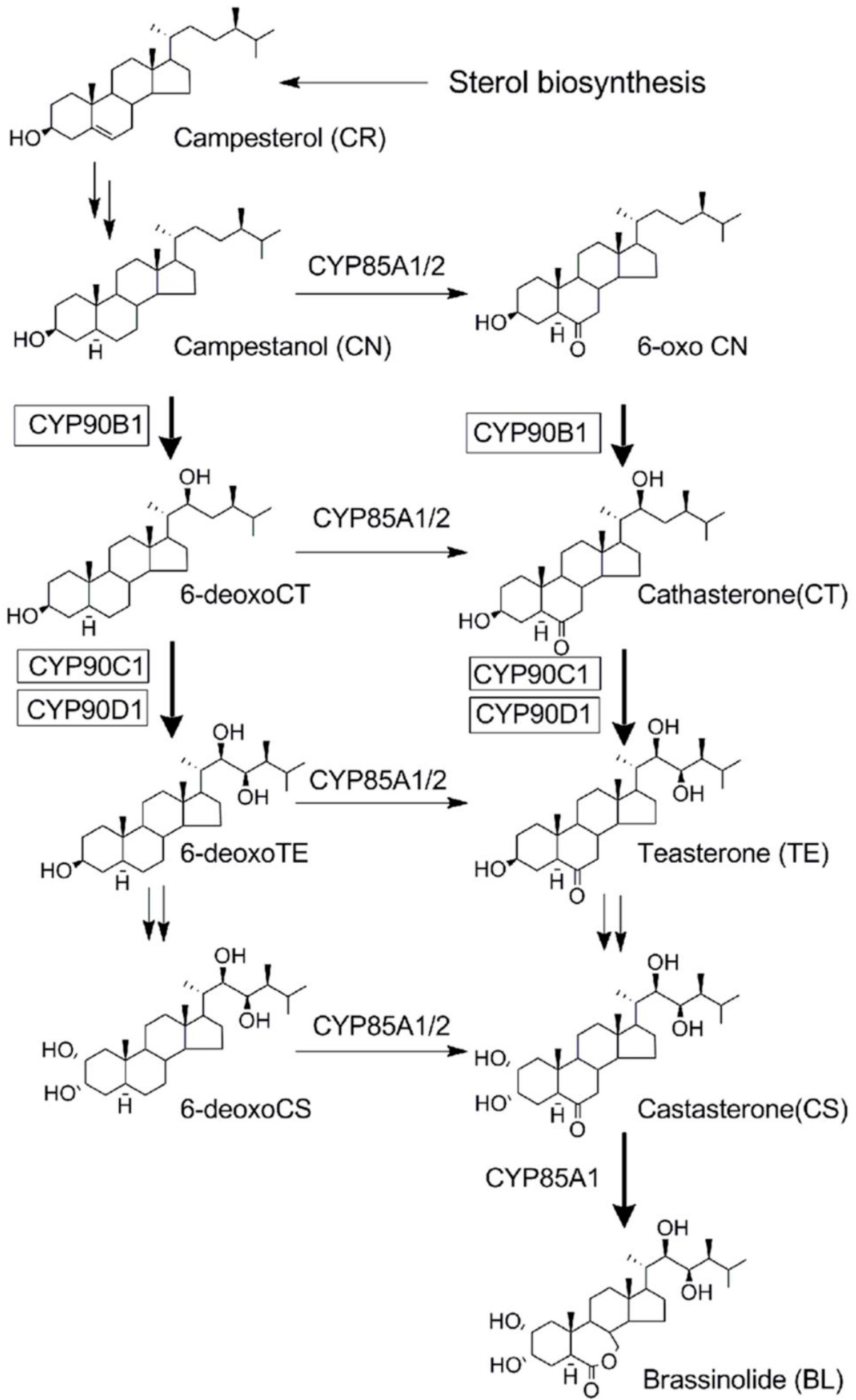{kind=link}
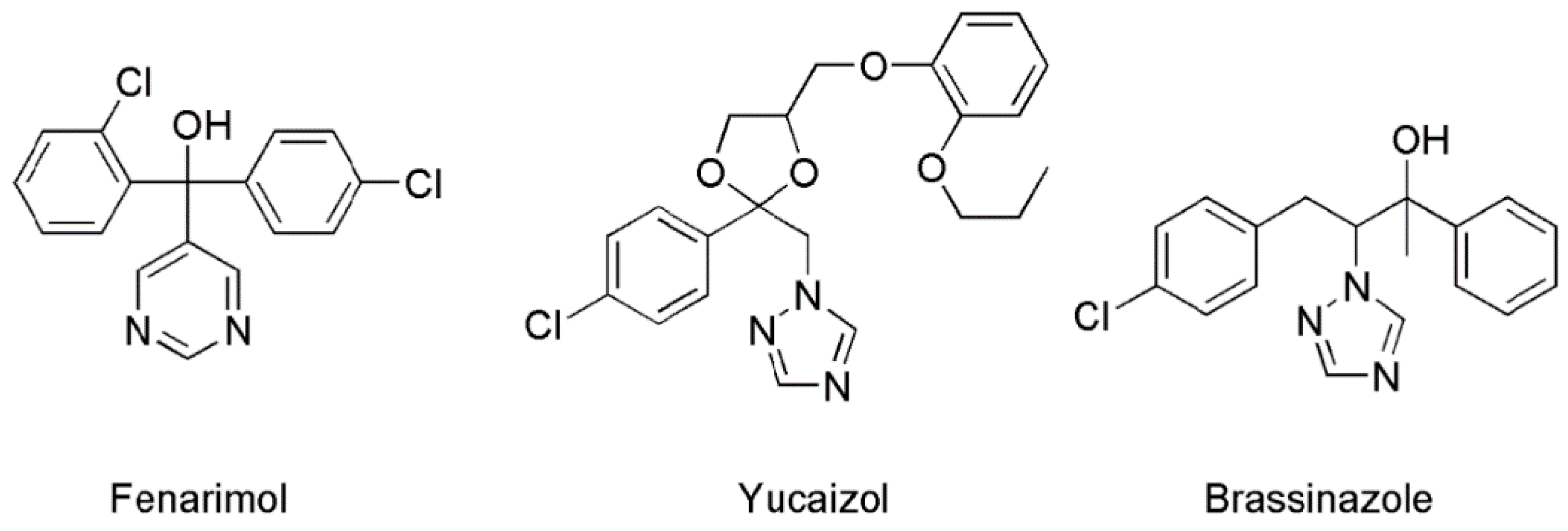{kind=link}
{kind=link}
{kind=link}
{kind=link}
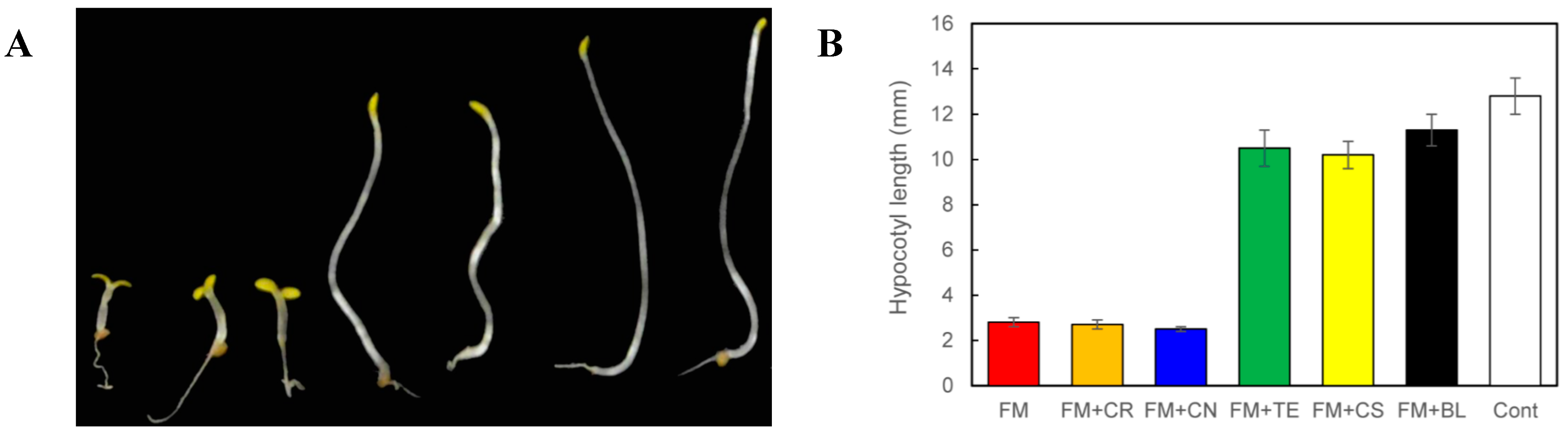{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction


2. Results
2.1. Fenarimol Induces Dwarfism and De-Etiolate Phenotypes of Arabidopsis Seedlings in the Dark


2.2. Brz-Resistant Mutant Bzr-1-1D/bil1-1D Displays Tolerance to Fenarimol Treatment

2.3. Determination of the Target Site(s) of Fenarimol on BR Biosynthesis Inhibition

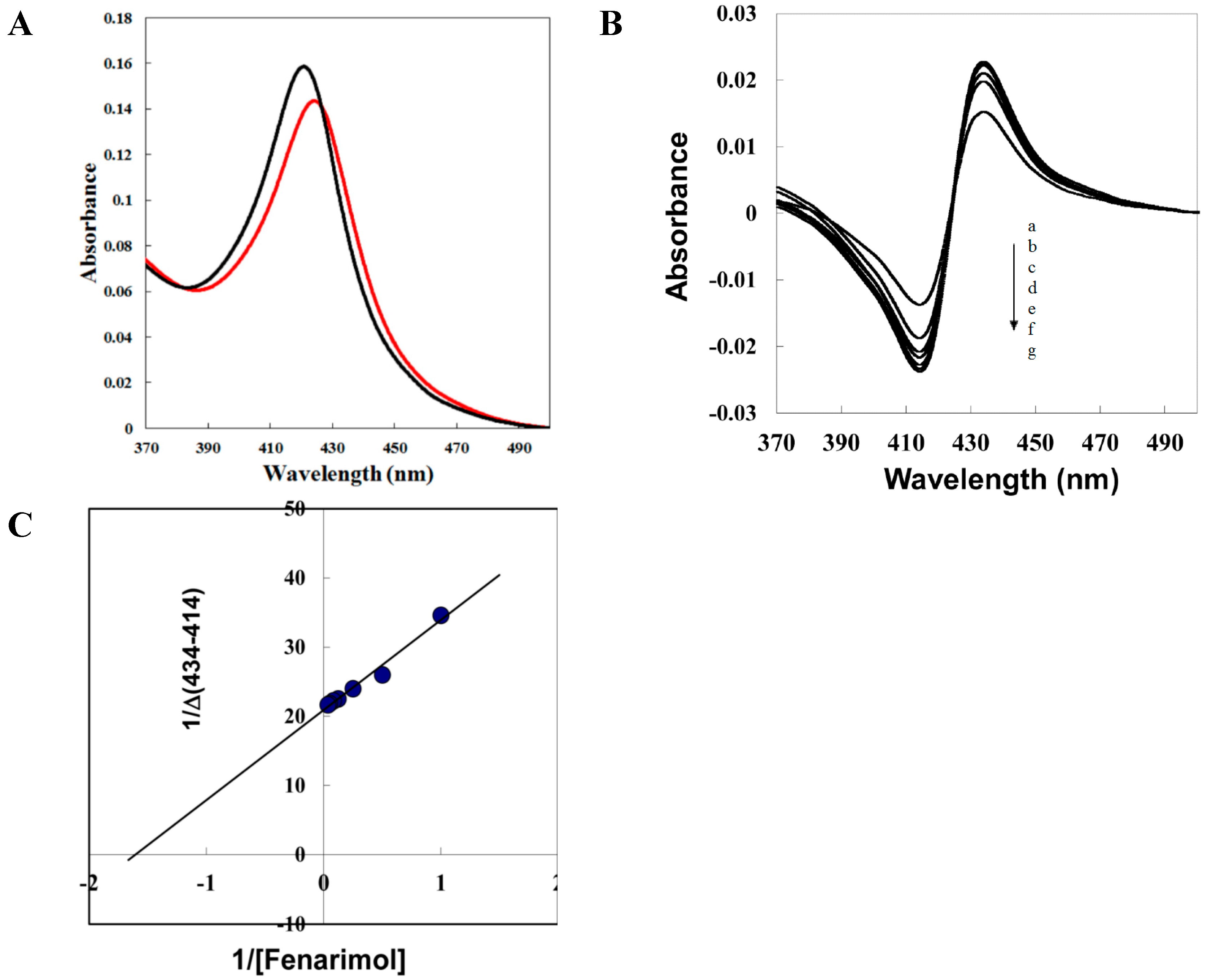
2.4. Fenarimol Binds to CYP90D1

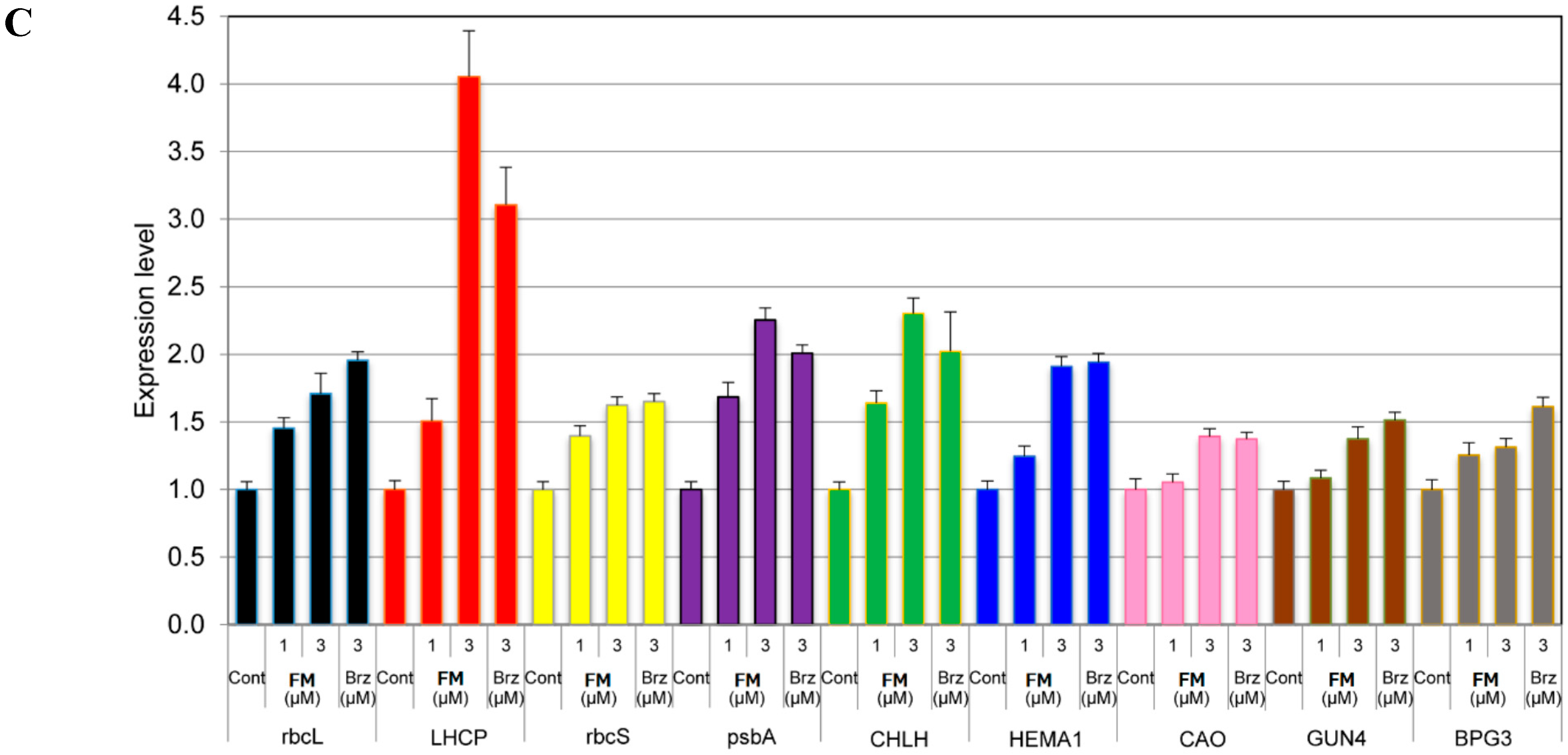
2.5. Molecular Function of Fenarimol for the Expression of BR-Responsive Genes


3. Discussion
4. Experimental Section
4.1. Chemicals
4.2. Plant Growth Conditions and BR Biosynthesis Inhibition Assay
4.3. Quantitative Real-Time PCR
4.4. Construction of CYP90D1 Expression Vectors
4.5. Expression and Purification of Recombinant CYP90D1
4.6. Binding Assay of Fenarimol to Recombinant CYP90D1
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Clouse, S.D.; Sasse, J.M. BRASSINOSTEROIDS: Essential regulators of plant growth and development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 427–451. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P. Brassinosteroid-mediated stress responses. J. Plant Growth Regul. 2003, 22, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Sasse, J.M. Physiological actions of brassinosteroids: An update. J. Plant Growth Regul. 2003, 22, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, T.; Chuck, G.S.; Fujioka, S.; Klempien, A.; Weizbauer, R; Potlurid, D.P.V.; Choe, S.; Johalf, G.S.; Schulz, B. Brassinosteroid control of sex determination in maize. Proc. Natl. Acad. Sci. USA 2011, 108, 19814–19819. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.; Dilkes, B.P.; Gregory, B.D.; Ross, A.S.; Yuan, H.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Tanaka, A.; Yoshida, S.; et al. The Arabidopsis dwarf1 mutant is defective in the conversion of 24-methylenecholesterol to campesterol in brassinosteroid biosynthesis. Plant Physiol. 1999, 119, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Ueguchi-Tanaka, M.; Umemura, K.; Uozu, S.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. Plant Cell 2003, 15, 2900–2910. [Google Scholar] [CrossRef] [PubMed]
- Bishop, G.J.; Nomura, T.; Yokota, T.; Harrison, K.; Noguchi, T; Fujioka, S.; Takatsuto, S.; Jonathan, D.G.J.; Kamiya, Y. The tomato DWARF enzyme catalyses C-6 oxidation in brassinosteroid biosynthesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1761–1766. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Nakayama, M.; Reid, J.B.; Takeuchi, Y.; Yokota, T. Blockage of brassinosteroid biosynthesis and sensitivity causes dwarfism in garden pea. Plant Physiol. 1997, 113, 31–37. [Google Scholar] [PubMed]
- Vriet, C.; Russinova, E.; Reuzeau, C. Boosting crop yields with plant steroids. Plant Cell 2012, 24, 842–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divi, U.K.; Krishna, P. Brassinosteroid: A biotechnological target for enhancing crop yield and stress tolerance. New Biotechnol. 2009, 26, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. “Green revolution” genes encode mutant gibberellin response modulators. Nature 1999, 400, 256–261. [Google Scholar] [PubMed]
- Blackwell, H.E.; Zhao, Y. Chemical genetic approaches to plant biology. Plant Physiol. 2003, 133, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Yamada, K.; Asami, T.; Yoshizawa, Y. Synthesis of novel brassinosteroid biosynthesis inhibitors based on the ketoconazole scaffold. Bioorg. Med. Chem. Lett. 2012, 22, 1625–1628. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Yoshizawa, Y.; Oh, K. Synthesis of 2RS, 4RS-1-[2-phenyl-4-[2-(2-trifluromethoxy-phenoxy)-ethyl]-1,3-dioxolan-2-yl-methyl]-1H-1,2,4-triazole derivatives as potent inhibitors of brassinosteroid biosynthesis. Molecules 2012, 17, 4460–4473. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Yajima, O.; Yoshizawa, Y.; Oh, K. Synthesis and biological evaluation of novel azole derivatives as selective potent inhibitors of brassinosteroid biosynthesis. Bioorg. Med. Chem. 2013, 21, 2451–2461. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Yamada, K.; Yoshizawa, Y. Asymmetric synthesis and effect of absolute stereochemistry of YCZ-2013, a brassinosteroid biosynthesis inhibitor. Bioorg. Med. Chem. Lett. 2013, 23, 6915–6919. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Sakurai, A. Studies on biosynthesis of brassinosteroids. Biosci. Biotechnol. Biochem. 1997, 61, 757–762. [Google Scholar]
- Choe, S.; Dilkes, B.P.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Feldmann, K.A. The DWF4 gene of Arabidopsis encodes a cytochrome P450 that mediates multiple 22-α-hydroxylation steps in brassinosteroid biosynthesis. Plant Cell 1998, 10, 231–243. [Google Scholar] [PubMed]
- Ohnishi, T.; Godza, B.; Watanabe, B.; Fujioka, S.; Hategan, L.; Ide, K.; Shibata, K.; Yokota, T.; Szekeres, M.; Mizutani, M. CYP90A1/CPD, a brassinosteroid biosynthetic cytochrome P450 of Arabidopsis, catalyzes C-3 oxidation. J. Biol. Chem. 2012, 287, 31551–31560. [Google Scholar] [CrossRef] [PubMed]
- Ohnish, T.; Szatmari, A.M.; Watanabe, B.; Fujita, S.; Bancos, S.; Koncz, C.; Lafos, M.; Shibata, K.; Yokota, T.; Sakata, K.; et al. C-23 hydroxylation by Arabidopsis CYP90C1 and CYP90D1 reveals a novel shortcut in brassinosteroid biosynthesis. Plant Cell 2006, 18, 3275–3288. [Google Scholar] [CrossRef] [PubMed]
- Castle, J.; Szekeres, M.; Jenkins, G.; Bishop, G.J. Unique and overlapping expression patterns of Arabidopsis CYP85 genes involved in brassinosteroid C-6 oxidation. Plant Mol. Biol. 2005, 57, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Jenner, P. Inhibitors of cytochrome P-450s and their mechanism of action. Drug Metab. Rev. 1981, 12, 1–117. [Google Scholar] [CrossRef] [PubMed]
- Rogerson, T.D.; Wilkinson, C.F.; Hetarski, K. Steric factors in the inhibitory interaction of imidazoles with microsomal enzymes. Biochem. Pharmacol. 1977, 26, 1039–1042. [Google Scholar] [CrossRef]
- Wang, J.M.; Asami, T.; Yoshida, S.; Murofushi, N. Biological evaluation of 5-substituted pyrimidine derivatives as inhibitors of brassinosteroid biosynthesis. Biosci. Biotechnol. Biochem. 2001, 65, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Li, J.; Choi, Y.H.; Seto, H.; Takatsuto, S.; Noguchi, T.; Watanabe, T.; Kuriyama, H.; Yokota, T.; Chory, J.; et al. The Arabidopsis deetiolated 2 mutant is blocked early in brassinosteroid biosynthesis. Plant Cell 1997, 9, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Matsumoto, T.; Yamagami, A.; Ogawa, A.; Yamada, K.; Suzuki, R.; Sawada, T.; Fujioka, S.; Yoshizawa, Y.; Nakano, T. YCZ-18 is a new brassinosteroid biosynthesis inhibitor. PLoS ONE 2015, 10, e0120812. [Google Scholar] [CrossRef] [PubMed]
- Asami, T.; Min, Y.K.; Nagata, N.; Yamagishi, K.; Takatsuto, S.; Fujioka, S.; Murofushi, N.; Yamaguchi, I.; Yoshida, S. Characterization of brassinazole, a triazole-type brassinosteroid biosynthesis inhibitor. Plant Physiol. 2000, 123, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Nakano, T.; Gendron, J.; He, J.; Chen, M.; Vafeados, D.; Yang, Y.; Fujioka, S.; Yoshida, S.; Asami, T.; et al. Nuclear-localized BZR1 mediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis. Dev. Cell 2002, 2, 505–513. [Google Scholar] [CrossRef]
- Asami, T.; Nakano, T.; Nakashita, H.; Sekimata, K.; Shimada, Y.; Yoshida, S. The influence of chemical genetics on plant science: Shedding light on functions and mechanism of action of brassinosteroids using biosynthesis inhibitors. J. Plant Growth Regul. 2003, 22, 336–349. [Google Scholar] [PubMed]
- Felpin, F.X.; Fouquet, E.A. Useful, reliable and safer protocol for hydrogenation and the hydrogenolysis of O-benzyl groups: The in situ preparation of an active Pd0/C catalyst with well-defined properties. Chem. Eur. J. 2010, 16, 12440–12445. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Purugganan, M.M.; Polisensky, D.H.; Antosiewicz, D.M.; Fry, S.C.; Braam, J. Arabidopsis TCH4, regulated by hormones and the environment, encodes a xyloglucan endotransglycosylase. Plant Cell 1995, 7, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Shimada, Y.; Goda, H.; Fujiwara, M.T.; Asami, T.; Yoshida, S. AXR1 is involved in BR-mediated elongation and SAUR-AC1 gene expression in Arabidopsis. FEBS Lett. 2003, 553, 28–32. [Google Scholar] [CrossRef]
- Goda, H.; Shimada, Y.; Asami, T.; Fujioka, S.; Yoshida, S. Microarray analysis of brassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 2002, 130, 1319–1334. [Google Scholar] [CrossRef] [PubMed]
- Shimada, S.; Komatsu, T.; Yamagami, A.; Nakazawa, M.; Matsui, M.; Kawaide, H.; Natsume, M.; Osada, H.; Asami, T.; Nakano, T. Formation and dissociation of the BSS1 protein complex regulates plant development via brassinosteroid signaling. Plant Cell 2015, 27, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Chory, J.; Nagpal, P.; Peto, C.A. Phenotypic and genetic analysis of det2, a new mutant that affects light-regulated seedling development in Arabidopsis. Plant Cell 1991, 3, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Krebbers, E.; Seurinck, J.; Herdies, L.; Cashmore, A.R.; Timko, M.P. Four genes in two diverged subfamilies encode the ribulose-1,5-bisphosphate carboxylase small subunit polypeptides of Arabidopsis thaliana. Plant Mol. Biol. 1988, 11, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Leutwiler, L.S.; Meyerowitz, E.M.; Tobin, E.M. Structure and expression of three light-harvesting chlorophyll a/b-binding protein genes in Arabidopsis thaliana. Nucleic Acids Res. 1986, 14, 4051–4064. [Google Scholar] [CrossRef] [PubMed]
- Fromm, H.; Devic, M.; Fluhr, R.; Edelman, M. Control of psbA gene expression: in mature Spirodela chloroplasts light regulation of 32-KD protein synthesis is independent of transcript level. EMBO J. 1985, 4, 291–295. [Google Scholar] [PubMed]
- Kim, G.T.; Tsukaya, H.; Uchimiya, H. The ROTUNDIFOLIA3 gene of Arabidopsis thaliana encodes a new member of the cytochrome P-450 family that is required for the regulated polar elongation of leaf cells. Genes Dev. 1998, 12, 2381–2391. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.T.; Fujioka, S.; Kozuka, T.; Tax, F.E.; Takatsuto, S.; Yoshida, S. CYP90C1 and CYP90D1 are involved in different steps in the brassinosteroid biosynthesis pathway in Arabidopsis thaliana. Plant J. 2005, 41, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Asami, T.; Mizutani, M.; Fujioka, S.; Goda, H.; Min, Y.K.; Shimada, Y.; Nakano, T.; Takatsuto, S.; Matsuyama, T.; Nagata, N.; et al. Selective interaction of triazole derivatives with DWF4, a cytochrome p450 monooxygenase of the brassinosteroid biosynthetic pathway, correlates with brassinosteroid deficiency in planta. J. Biol. Chem. 2001, 276, 25687–25691. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, A.; Nakazawa, M.; Matsui, M.; Tujimoto, M.; Sakuta, M.; Asami, T.; Nakano, T. Chemical genetics reveal the novel transmembrane protein BIL4, which mediates plant cell elongation in brassinosteroid signaling. Biosci. Biotechnol. Biochem. 2009, 73, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Carninci, P.; Nishiyama, Y.; Hayashizak, Y.; Shinozaki, K. High-efficiency cloning of Arabidopsis full-length cDNA by biotinylated CAP trapper. Plant J. 1998, 15, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Narusaka, M.; Kamiya, A.; Ishida, J.; Satou, M.; Tetsuya, S.; Maiko, N.; Akiko, E.; Kenji, A.; Youko, O.; et al. Functional annotation of a full-length Arabidopsis cDNA collection. Science 2002, 296, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Kojima, C. pCold-GST vector: A novel cold-shock vector containing GST tag for soluble protein production. Protein Expr. Purif. 2008, 62, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Asami, T.; Matsui, K.; Howe, G.A.; Murofushi, N. Characterization of novel imidazole derivative, JM-8686, a potent inhibitor of allene oxide synthase. FEBS Lett. 2006, 580, 5791–5796. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, K.; Matsumoto, T.; Yamagami, A.; Hoshi, T.; Nakano, T.; Yoshizawa, Y. Fenarimol, a Pyrimidine-Type Fungicide, Inhibits Brassinosteroid Biosynthesis. Int. J. Mol. Sci. 2015, 16, 17273-17288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160817273
Oh K, Matsumoto T, Yamagami A, Hoshi T, Nakano T, Yoshizawa Y. Fenarimol, a Pyrimidine-Type Fungicide, Inhibits Brassinosteroid Biosynthesis. International Journal of Molecular Sciences. 2015; 16(8):17273-17288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160817273
Chicago/Turabian StyleOh, Keimei, Tadashi Matsumoto, Ayumi Yamagami, Tomoki Hoshi, Takeshi Nakano, and Yuko Yoshizawa. 2015. "Fenarimol, a Pyrimidine-Type Fungicide, Inhibits Brassinosteroid Biosynthesis" International Journal of Molecular Sciences 16, no. 8: 17273-17288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160817273