
Impacts of Alternative Splicing Events on the Differentiation of Adipocytes

Abstract
:1. Introduction
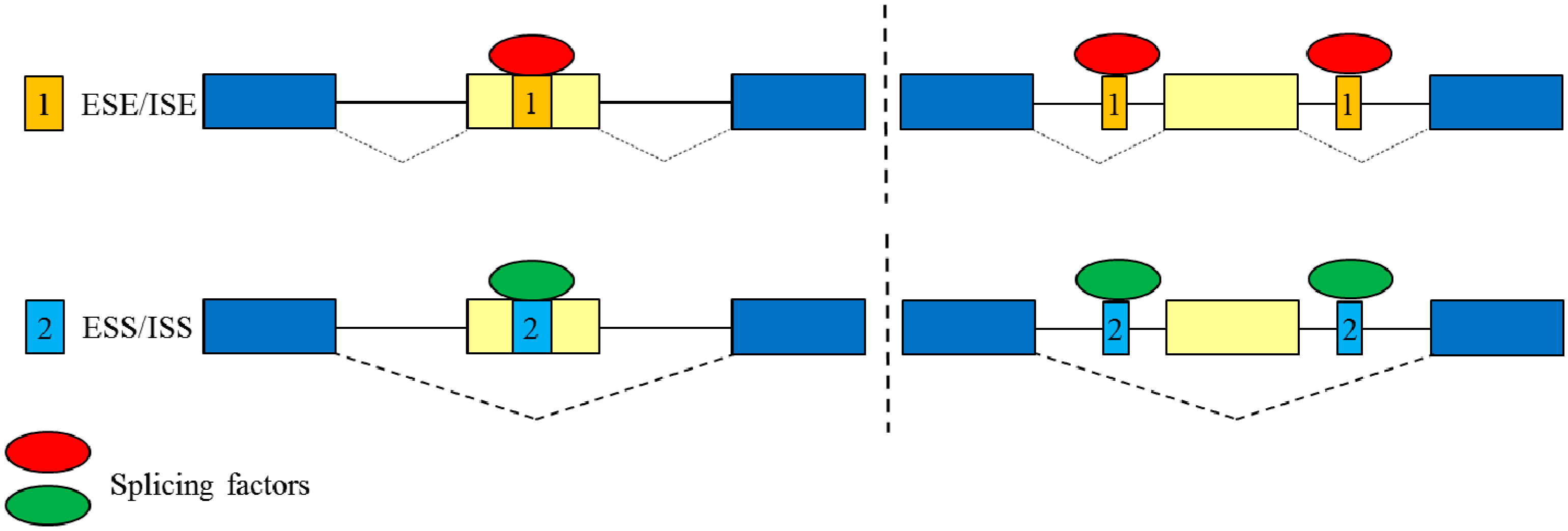
2. Overview of Alternative Splicing

3. Overview of Adipocyte Differentiation
3.1. White Adipocytes
3.2. Brown Adipocytes
4. Impacts of Alternative Splicing Events on Adipogenesis
4.1. Adipogenesis-Related Splicing Factors
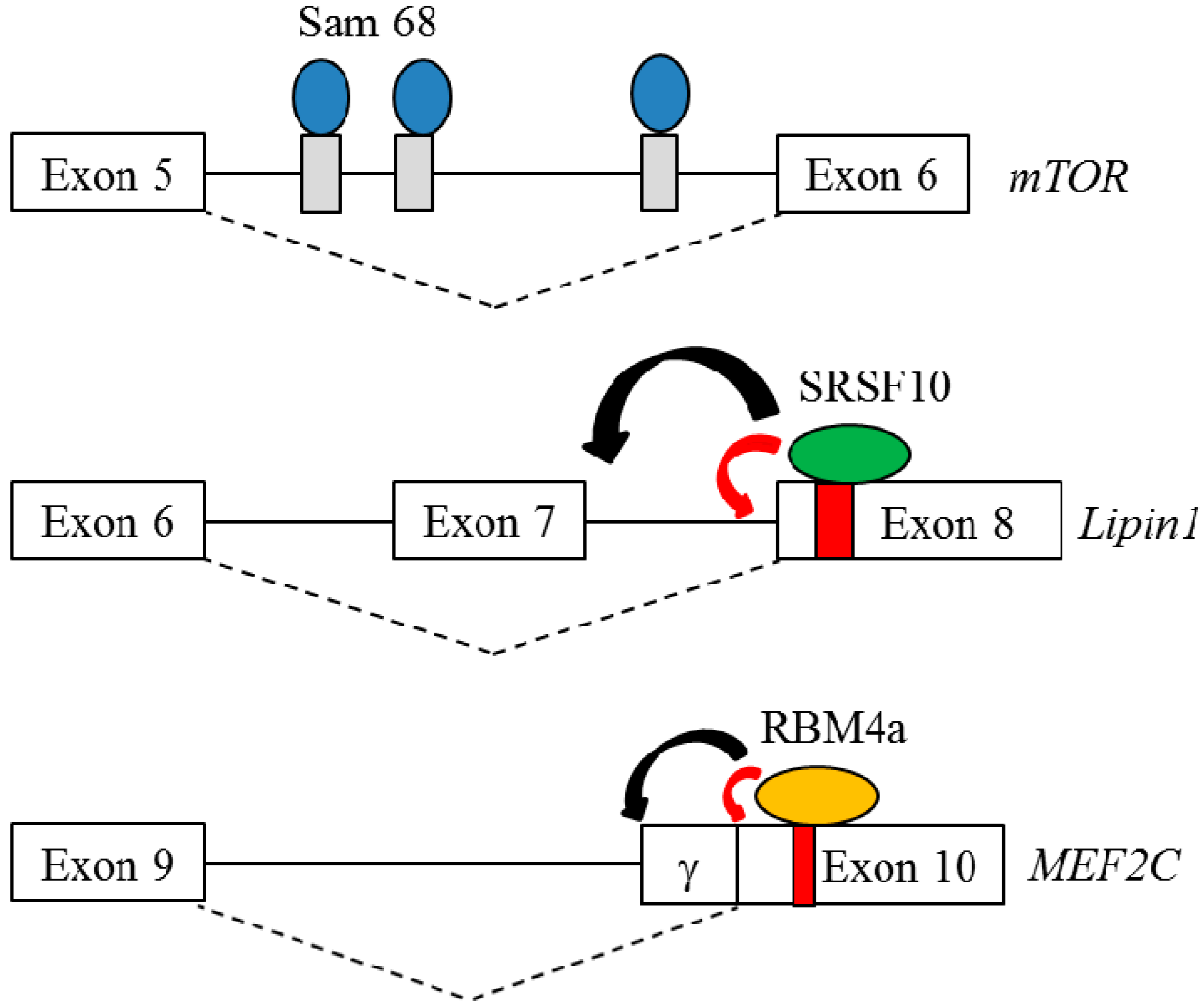
4.1.1. Src-Associated Substrate During Mitosis of 68 kDa (Sam68)

4.1.2. Serine/Arginine-Rich Splicing Factor 10 (SRSF10)
4.1.3. Fat Mass and Obesity-Associated Protein (FTO) and SRSF2
4.1.4. Zinc Finger Protein 638 (ZNF638)
4.1.5. RNA-Binding Motif Protein 4a (RBM4a)

{kind=link}
{kind=link}
{kind=link}
| Splicing Factor or Regulator | AS Event | Adipocyte | Biological Signatures | References |
|---|---|---|---|---|
| Sam68 | mTOR | WAT | Promote adipogenesis | [41,42] |
| SRSF10 | Lipin1 | WAT | Differentiation/Lipid storage | [48,51,52] |
| SRp40 | PPARγ | Pre-adipocyte | Promote adipogenesis | [64,65] |
| SRSF2 & FTO | RUNX1T1 | WAT | Promote adipogenesis | [54,55] |
| RBM4a | PPARg, Pref-1 INSR, MEF2C | BAT | Enhance differentiation and metabolismof BAs | [12,13,62,63] |
4.2. Adipocyte-Related Alternative Splicing Events
4.2.1. Nuclear Receptor Co-Repressor 1 (NCoR)
4.2.2. Protein Kinase Cδ (PKCδ)
4.2.3. Cholesteryl Ester Transfer Protein (CETP)
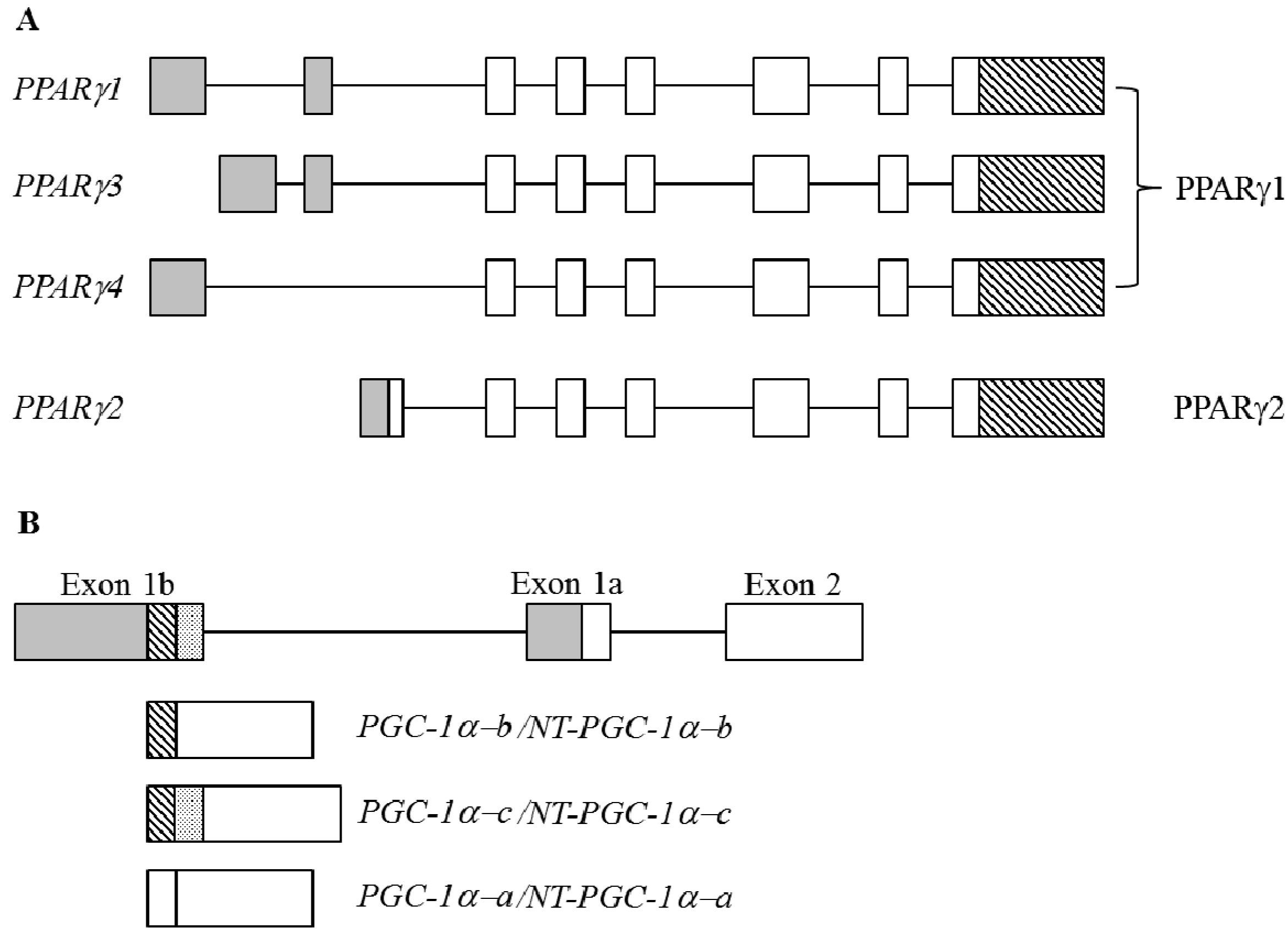
4.2.4. Peroxisome Proliferator-Activated Receptor γ (PPARγ)
4.2.5. PPARγ Coactivator 1α (PGC-1α)

4.2.6. Mitochondrial Oxodicarboxylate Carrier (ODC)
| Gene | AS Region | Adipocyte Isoform | Biological Signatures | References |
|---|---|---|---|---|
| mTOR | Intron 5 | Excluded | Promote white adipogenesis | [42] |
| Lipin 1 | Exon 7 | Included/Excluded | Differentiation/Lipid storage | [48,51,52] |
| INSR | Exon 11 | Included | Promote brown adipogenesis | [13,62] |
| MEF2C | Alternative 3′ splice site of Exon 10 | Distal | Promote brown adipogenesis | [63] |
| NcoR | Exon 37b | Included/Excluded | Proliferation/Differentiation | [66,67,68,69,70,71,72,73,74,75,76,77] |
| PKCδ | Alternative 5′ splice site of Exon 9 | Proximal | Promote adipogenesis | [30,78,79,80,81,82,83,84,85] |
| CETP | Exon 9 | Included | Promote lipid metabolism | [86,87,88,89,90,91] |
| RUNX1T1 | Exon 6 | Included | Promote white adipogenesis | [54] |
| ODC | Exon 1, 2 and 3 | Selection of exon 2 and 3 | Promote lipid accumulation | [105,106,107] |
5. High-Throughput Methods Applied to Tissue-Specific Alternative Splicing
6. Conclusions and Perspectives
Conflicts of Interest
References
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Quesnel-Vallières, M.; Irimia, M.; Cordes, S.P.; Blencowe, B.J. Essential roles for the splicing regulator nSR100/SRRM4 during nervous system development. Genes Dev. 2015, 29, 746–759. [Google Scholar]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Matera, A.G.; Wang, Z. A day in the life of the spliceosome. Nat. Rev. Mol. Cell Biol. 2014, 15, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Fan, J.; Wang, J.; Yang, S.; Cong, Q.; Wang, R.; Lv, Q.; Liu, R.; Ning, G. High levels of chorionic gonadotrophin attenuate insulin sensitivity and promote inflammation in adipocytes. J. Mol. Endocrinol. 2015, 54, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Al-Daghri, N.M.; Al-Attas, O.S.; Krishnaswamy, S.; Mohammed, A.K.; Alenad, A.M.; Chrousos, G.P.; Alokail, M.S. Association of Type 2 Diabetes Mellitus related SNP genotypes with altered serum adipokine levels and metabolic syndrome phenotypes. Int. J. Clin. Exp. Med. 2015, 8, 4464–4471. [Google Scholar] [PubMed]
- Giordano, A.; Smorlesi, A.; Frontini, A.; Barbatelli, G.; Cinti, S. White, brown and pink adipocytes: The extraordinary plasticity of the adipose organ. Eur. J. Endocrinol. 2014, 170, R159–R171. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, T.; Kajita, K.; Yamada, K.; Miura, A.; Kanoh, Y.; Ishizawa, M.; Wada, H.; Itaya, S.; Yamamoto, M.; Yasuda, K.; et al. Insulin regulated PKC isoform mRNA in rat adipocytes. Diabetes Res. Clin. Pract. 1996, 33, 159–167. [Google Scholar] [CrossRef]
- Bradbury, M.W.; Berk, P.D. Mitochondrial aspartate aminotransferase: Direction of a single protein with two distinct functions to two subcellular sites does not require alternative splicing of the mRNA. Biochem. J. 2000, 345, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Mei, B.; Zhao, L.; Chen, L.; Sul, H.S. Only the large soluble form of preadipocyte factor-1 (Pref-1), but not the small soluble and membrane forms, inhibits adipocyte differentiation: Role of alternative splicing. Biochem. J. 2002, 364, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Entingh, A.J.; Taniguchi, C.M.; Kahn, C.R. Bi-directional regulation of brown fat adipogenesis by the insulin receptor. J. Biol. Chem. 2003, 278, 33377–33383. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Moore, M.J. The spliceosome: Disorder and dynamics defined. Curr. Opin. Struct. Biol. 2014, 24, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Papasaikas, P.; Tejedor, J.R.; Vigevani, L.; Valcárcel, J. Functional splicing network reveals extensive regulatory potential of the core spliceosomal machinery. Mol. Cell 2015, 57, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Cyphert, T.J.; Suchanek, A.L.; Griffith, B.N.; Salati, L.M. Starvation actively inhibits splicing of glucose-6-phosphate dehydrogenase mRNA via a bifunctional ESE/ESS element bound by hnRNP K. Biochim. Biophys. Acta 2013, 1829, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Erkelenz, S.; Mueller, W.F.; Evans, M.S.; Busch, A.; Schöneweis, K.; Hertel, K.J.; Schaal, H. Position-dependent splicing activation and repression by SR and hnRNP proteins rely on common mechanisms. RNA 2013, 19, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Chiellini, C.; Cochet, O.; Negroni, L.; Samson, M.; Poggi, M.; Ailhaud, G.; Alessi, M.C.; Dani, C.; Amri, E.Z. Characterization of human mesenchymal stem cell secretome at early steps of adipocyte and osteoblast differentiation. BMC Mol. Biol. 2008, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Bowers, R.R.; Lane, M.D. A role for bone morphogenetic protein-4 in adipocyte development. Cell Cycle 2007, 6, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Li, X.; Tang, Q.Q. Transcriptional regulation of adipocyte differentiation: A central role for CCAAT/enhancer-binding protein (C/EBP) β. J. Biol. Chem. 2015, 290, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Guo, B.; Chatterjee, S.; Chen, M.H.; Nelson, D.; Yechoor, V.K.; Ma, K. The adipocyte clock controls brown adipogenesis through the TGF-β and BMP signaling pathways. J. Cell Sci. 2015, 128, 1835–1847. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhou, Y.; Hu, X.; Peng, X.; Wei, H.; Peng, J.; Jiang, S. Activation of PPARγ2 by PPARγ1 through a functional PPRE in transdifferentiation of myoblasts to adipocytes induced by EPA. Cell Cycle 2015, 14, 1830–1841. [Google Scholar] [CrossRef] [PubMed]
- Majka, S.M.; Fox, K.E.; Psilas, J.C.; Helm, K.M.; Childs, C.R.; Acosta, A.S.; Janssen, R.C.; Friedman, J.E.; Woessner, B.T.; Shade, T.R. De novo generation of white adipocytes from the myeloid lineage via mesenchymal intermediates is age, adipose depot, and gender specific. Proc. Natl. Acad. Sci. USA 2010, 107, 14781–14786. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.J.; Ishibashi, J.; Wang, W.; Lim, H.W.; Goyama, S.; Sato, T.; Kurokawa, M.; Won, K.J.; Seale, P. Prdm16 is required for the maintenance of brown adipocyte identity and function in adult mice. Cell Metab. 2014, 19, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.W.; Tang, Y.; Li, X.; Liu, Y.; Zhang, Y.Y.; Huang, H.Y.; Xue, R.D.; Yu, H.Y.; Guo, L.; Gao, H.D. BMP4-mediated brown fat-like changes in white adipose tissue alter glucose and energy homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Gaya, M.; Repetto, V.; Toneatto, J.; Anesini, C.; Piwien-Pilipuk, G.; Moreno, S. Antiadipogenic effect of carnosic acid, a natural compound present in Rosmarinus officinalis, is exerted through the C/EBPs and PPARγ pathways at the onset of the differentiation program. Biochim. Biophys. Acta 2013, 1830, 3796–3806. [Google Scholar] [CrossRef] [PubMed]
- Lechner, S.; Mitterberger, M.C.; Mattesich, M.; Zwerschke, W. Role of C/EBPβ-LAP and C/EBPβ-LIP in early adipogenic differentiation of human white adipose-derived progenitors and at later stages in immature adipocytes. Differentiation 2013, 85, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Morita, M.; Yokoyama, K.; Suzuki, T.; Yamamoto, T. Tob2 inhibits peroxisome proliferator-activated receptor γ2 expression by sequestering Smads and C/EBPα during adipocyte differentiation. Mol. Cell. Biol. 2012, 32, 5067–5077. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Chen, X.Y.; Zhang, Z.J.; Kang, Y.; Liao, W.M.; Yu, W.H.; Xiang, A.P. Expression patterns of transcription factor PPARγ and C/EBP family members during in vitro adipogenesis of human bone marrow mesenchymal stem cells. Cell Biol. Int. 2015, 39, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J., Jr.; Liu, X.S.; et al. PPARγ and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.M.; Namkung, J.; Go, Y.; Shong, K.E.; Kim, K.; Kim, H.; Park, B.Y.; Lee, H.W.; Jeon, Y.H.; Song, J.; et al. Regulation of systemic energy homeostasis by serotonin in adipose tissues. Nat. Commun. 2015, 13, 6794. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.M.; Tseng, Y.H.; Schulz, T.J. Adipogenic fate commitment of muscle-derived progenitor cells: Isolation, culture, and differentiation. Methods Mol. Biol. 2014, 1213, 229–243. [Google Scholar] [PubMed]
- Hung, C.M.; Calejman, C.M.; Sanchez-Gurmaches, J.; Li, H.; Clish, C.B.; Hettmer, S.; Wagers, A.J.; Guertin, D.A. Rictor/mTORC2 loss in the Myf5 lineage reprograms brown fat metabolism and protects mice against obesity and metabolic disease. Cell Rep. 2014, 8, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Li, G.; Yang, X.; Ba, K.; Fu, Y.; Fu, N.; Cai, X.; Li, G.; Chen, Q.; Wang, M.; et al. Effects of bone morphogenetic protein-4 (BMP-4) on adipocyte differentiation from mouse adipose-derived stem cells. Cell Prolif. 2013, 46, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Takagi, T.; Kanesashi, S.N.; Kurahashi, T.; Nomura, T.; Harada, J.; Ishii, S. Schnurri-2 controls BMP-dependent adipogenesis via interaction with Smad proteins. Dev. Cell 2006, 10, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Okla, M.; Ha, J.H.; Temel, R.E.; Chung, S. BMP7 drives human adipogenic stem cells into metabolically active beige adipocytes. Lipids 2015, 50, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Bjork, B.; Yang, W.; Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scimè, A.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.H.; Kokkotou, E.; Schulz, T.J.; Huang, T.L.; Winnay, J.N.; Taniguchi, C.M.; Tran, T.T.; Suzuki, R.; Espinoza, D.O.; Yamamoto, Y.; et al. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure. Nature 2008, 454, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Resnick, R.J.; Rakowski, C.; Shalloway, D.; Taylor, S.J.; Blobel, G.A. Physical and functional interaction between the transcriptional cofactor CBP and the KH domain protein Sam68. Mol. Cancer Res. 2002, 1, 48–55. [Google Scholar] [PubMed]
- Itoh, M.; Haga, I.; Li, Q.H.; Fujisawa, J. Identification of cellular mRNA targets for RNA-binding protein Sam68. Nucleic Acids Res. 2002, 30, 5452–5464. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Cheng, M.; Boriboun, C.; Ardehali, M.M.; Jiang, C.; Liu, Q.; Han, S.; Goukassian, D.A.; Tang, Y.L.; Zhao, T.C. Inhibition of Sam68 triggers adipose tissue browning. J. Endocrinol. 2015, 225, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Huot, M.É.; Vogel, G.; Zabarauskas, A.; Ngo, C.T.; Coulombe-Huntington, J.; Majewski, J.; Richard, S. The Sam68 STAR RNA-binding protein regulates mTOR alternative splicing during adipogenesis. Mol. Cell 2012, 46, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Pratt, G.A.; Martinez, F.J.; Yeo, G.W.; Lykke-Andersen, J. Target Discrimination in Nonsense-Mediated mRNA Decay Requires Upf1 ATPase Activity. Mol. Cell 2015, 59, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Richard, S. Sam68 Regulates S6K1 Alternative Splicing during Adipogenesis. Mol. Cell. Biol. 2015, 35, 1926–1939. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wu, W.; Li, H.; Cheng, Y.; Wei, N.; Zong, J.; Feng, X.; Xie, Z.; Chen, D.; Manley, J.L.; et al. Transcriptome analysis of alternative splicing events regulated by SRSF10 reveals position-dependent splicing modulation. Nucleic Acids Res. 2014, 42, 4019–4030. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.; Manley, J.L. The SR protein SRp38 represses splicing in M phase cells. Cell 2002, 111, 407–417. [Google Scholar] [CrossRef]
- Feng, Y.; Valley, M.T.; Lazar, J.; Yang, A.L.; Bronson, R.T.; Firestein, S.; Coetzee, W.A.; Manley, J.L. SRp38 regulates alternative splicing and is required for Ca2+ handling in the embryonic heart. Dev. Cell 2009, 16, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cheng, Y.; Wu, W.; Liu, Y.; Wei, N.; Feng, X.; Xie, Z.; Feng, Y. SRSF10 regulates alternative splicing and is required for adipocyte differentiation. Mol. Cell. Biol. 2014, 34, 2198–2207. [Google Scholar] [CrossRef] [PubMed]
- Gauger, K.J.; Bassa, L.M.; Henchey, E.M.; Wyman, J.; Bentley, B.; Brown, M.; Shimono, A.; Schneider, S.S. Mice deficient in Sfrp1 exhibit increased adiposity, dysregulated glucose metabolism, and enhanced macrophage infiltration. PLoS ONE 2013, 8, e78320. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of adipogenesis by Wnt signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.K.; Lee, M.Y.; Kim, J.W.; Kim, M.; Moon, J.S.; Lee, Y.J.; Ahn, Y.H.; Kim, K.S. Lipin1 is a key factor for the maturation and maintenance of adipocytes in the regulatory network with CCAAT/enhancer-binding protein α and peroxisome proliferator-activated receptor γ 2. J. Biol. Chem. 2008, 283, 34896–34906. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ji, C.; Huang, J.; Yang, F.; Zhang, H.; Liu, L.; Yin, J. The mRNA of lipin1 and its isoforms are differently expressed in the longissimus dorsi muscle of obese and lean pigs. Mol. Biol. Rep. 2011, 38, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Ronkainen, J.; Huusko, T.J.; Soininen, R.; Mondini, E.; Cinti, F.; Mäkelä, K.A.; Kovalainen, M.; Herzig, K.H.; Järvelin, M.R.; Sebert, S.; et al. Fat mass- and obesity-associated gene Fto affects the dietary response in mouse white adipose tissue. Sci. Rep. 2015, 5, 9233. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, Y.; Sun, B.F.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.J.; Ping, X.L.; Chen, Y.S.; Wang, W.J.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, L.; Chen, J.; Wang, Y. mRNA m6A methylation downregulates adipogenesis in porcine adipocytes. Biochem. Biophys. Res. Commun. 2015, 459, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Razin, S.V.; Borunova, V.V.; Maksimenko, O.G.; Kantidze, O.L. Cys2His2 zinc finger protein family: Classification, functions, and major members. Biochemistry 2012, 77, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Meruvu, S.; Hugendubler, L.; Mueller, E. Regulation of adipocyte differentiation by the zinc finger protein ZNF638. J. Biol. Chem. 2011, 286, 26516–26523. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Ma, X.; Meruvu, S.; Hugendubler, L.; Mueller, E. The adipogenic transcriptional cofactor ZNF638 interacts with splicing regulators and influences alternative splicing. J. Lipid Res. 2014, 55, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Tarn, W.Y. RBM4 down-regulates PTB and antagonizes its activity in muscle cell-specific alternative splicing. J. Cell Biol. 2011, 193, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Tarn, W.Y. RNA-binding motif protein 4 translocates to cytoplasmic granules and suppresses translation via argonaute2 during muscle cell differentiation. J. Biol. Chem. 2009, 284, 34658–34665. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Yan, Y.T.; Hsieh, W.K.; Peng, P.J.; Su, C.H.; Tarn, W.Y. RBM4 promotes pancreas cell differentiation and insulin expression. Mol. Cell. Biol. 2013, 33, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Tarn, W.Y.; Hsieh, W.K. Emerging role for RNA binding motif protein 4 in the development of brown adipocytes. Biochim. Biophys. Acta 2014, 1843, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C. RBM4-MEF2C network constitutes a feed-forward circuit that facilitates the differentiation of brown adipocytes. RNA Biol. 2015, 12, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.R.; Carter, G.; Li, P.; Patel, R.; Watson, J.E.; Patel, N.A. Long Non-Coding RNA NEAT1 Associates with SRp40 to Temporally Regulate PPARγ2 Splicing during Adipogenesis in 3T3-L1 Cells. Genes 2014, 5, 1050–1063. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Carter, G.; Romero, J.; Gower, K.M.; Watson, J.; Patel, N.A.; Cooper, D.R. Clk/STY (cdc2-like kinase 1) and Akt regulate alternative splicing and adipogenesis in 3T3-L1 pre-adipocytes. PLoS ONE 2013, 8, e53268. [Google Scholar] [CrossRef] [PubMed]
- Goodson, M.L.; Young, B.M.; Snyder, C.A.; Schroeder, A.C.; Privalsky, M.L. Alteration of NCoR corepressor splicing in mice causes increased body weight and hepatosteatosis without glucose intolerance. Mol. Cell. Biol. 2014, 34, 4104–4114. [Google Scholar] [CrossRef] [PubMed]
- Faist, F.; Short, S.; Kneale, G.G.; Sharpe, C.R. Alternative splicing determines the interaction of SMRT isoforms with nuclear receptor-DNA complexes. Biosci. Rep. 2009, 29, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.M.; Guo, C.; Zhang, J. Nuclear receptor corepressor complexes in cancer: Mechanism, function and regulation. Am. J. Clin. Exp. Urol. 2014, 2, 169–187. [Google Scholar] [PubMed]
- Vella, K.R.; Ramadoss, P.; Costa-E-Sousa, R.H.; Astapova, I.; Ye, F.D.; Holtz, K.A.; Harris, J.C.; Hollenberg, A.N. Thyroid hormone signaling in vivo requires a balance between coactivators and corepressors. Mol. Cell. Biol. 2014, 34, 1564–1575. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Li, Y.; Gow, C.H.; Wong, M.; Zha, J.; Yan, C.; Liu, H.; Wang, Y.; Burris, T.P.; Zhang, J. The optimal corepressor function of nuclear receptor corepressor (NCoR) for peroxisome proliferator-activated receptor γ requires G protein pathway suppressor 2. J. Biol. Chem. 2015, 290, 3666–3679. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Spann, N.J.; Kaikkonen, M.U.; Lu, M.; Oh da, Y.; Fox, J.N.; Bandyopadhyay, G.; Talukdar, S.; Xu, J.; Lagakos, W.S.; et al. NCoR repression of LXRs restricts macrophage biosynthesis of insulin-sensitizing omega 3 fatty acids. Cell 2013, 155, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Bartella, V.; Rizza, P.; Barone, I.; Zito, D.; Giordano, F.; Giordano, C.; Catalano, S.; Mauro, L.; Sisci, D.; Panno, M.L.; et al. Estrogen receptor β binds Sp1 and recruits a corepressor complex to the estrogen receptor α gene promoter. Breast Cancer Res. Treat. 2012, 134, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Feng, D.; Fang, B.; Mullican, S.E.; You, S.H.; Lim, H.W.; Everett, L.J.; Nabel, C.S.; Li, Y.; Selvakumaran, V.; et al. Deacetylase-independent function of HDAC3 in transcription and metabolism requires nuclear receptor corepressor. Mol. Cell 2013, 52, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Emont, M.P.; Mantis, S.; Kahn, J.H.; Landeche, M.; Han, X.; Sargis, R.M.; Cohen, R.N. Silencing Mediator of Retinoid and Thyroid Hormone Receptors (SMRT) regulates glucocorticoid action in adipocytes. Mol. Cell. Endocrinol. 2015, 407, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Dib, L.; Bugge, A.; Collins, S. LXRα fuels fatty acid-stimulated oxygen consumption in white adipocytes. J. Lipid Res. 2014, 55, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Malartre, M.; Short, S.; Sharpe, C. Alternative splicing generates multiple SMRT transcripts encoding conserved repressor domains linked to variable transcription factor interaction domains. Nucleic Acids Res. 2004, 32, 4676–4686. [Google Scholar] [CrossRef] [PubMed]
- Snyder, C.A.; Goodson, M.L.; Schroeder, A.C.; Privalsky, M.L. Regulation of corepressor alternative mRNA splicing by hormonal and metabolic signaling. Mol. Cell. Endocrinol. 2015, 413, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Fang, N.; Lou, J.; Zhang, W.; Xu, S.; Liu, H.; Fang, Q.; Wang, Z.; Liu, J.; Men, X.; et al. TRB3 is involved in free fatty acid-induced INS-1-derived cell apoptosis via the protein kinase C δ pathway. PLoS ONE 2014, 9, e96089. [Google Scholar] [CrossRef] [PubMed]
- Gautrey, H.L.; Tyson-Capper, A.J. Regulation of Mcl-1 by SRSF1 and SRSF5 in cancer cells. PLoS ONE 2012, 7, e51497. [Google Scholar] [CrossRef] [PubMed]
- Apostolatos, A.; Song, S.; Acosta, S.; Peart, M.; Watson, J.E.; Bickford, P.; Cooper, D.R.; Patel, N.A. Insulin promotes neuronal survival via the alternatively spliced protein kinase CδII isoform. J. Biol. Chem. 2012, 287, 9299–9310. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Apostolatos, A.; Carter, G.; Ajmo, J.; Gali, M.; Cooper, D.R.; You, M.; Bisht, K.S.; Patel, N.A. Protein kinase C δ (PKCδ) splice variants modulate apoptosis pathway in 3T3L1 cells during adipogenesis: Identification of PKCδII inhibitor. J. Biol. Chem. 2013, 288, 26834–26846. [Google Scholar] [CrossRef] [PubMed]
- Funcke, J.B.; Zoller, V.; El Hay, M.A.; Debatin, K.M.; Wabitsch, M.; Fischer-Posovszky, P. TNF-related apoptosis-inducing ligand promotes human preadipocyte proliferation via ERK1/2 activation. FASEB J. 2015, 29, 3065–3075. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.; Apostolatos, A.; Patel, R.; Mathur, A.; Cooper, D.; Murr, M.; Patel, N.A. Dysregulated Alternative Splicing Pattern of PKCδ during Differentiation of Human Preadipocytes Represents Distinct Differences between Lean and Obese Adipocytes. ISRN Obes. 2013, 2013, 161345. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, D.; Li, F.; Shi, J.; Song, J. Different roles of protein kinase C-βI and -Δ in the regulation of adipocytedifferentiation. Int. J. Biochem. Cell Biol. 2006, 38, 2151–2163. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.S.; Carter, G.; Cooper, D.R.; Apostolatos, H.; Patel, N.A. Transformer 2β homolog (Drosophila) (TRA2B) regulates protein kinase C δI (PKCδI) splice variant expression during 3T3L1 preadipocyte cell cycle. J. Biol. Chem. 2014, 289, 31662–31672. [Google Scholar] [CrossRef] [PubMed]
- Rohatgi, A. High-density lipoprotein function measurement in human studies: Focus on cholesterol efflux capacity. Prog. Cardiovasc. Dis. 2015, 58, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Suhy, A.; Hartmann, K.; Papp, A.C.; Wang, D.; Sadee, W. Regulation of cholesteryl ester transfer protein expression by upstream polymorphisms: Reduced expression associated with rs247616. Pharmacogenet. Genom. 2015, 25, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Santamarina-Fojo, S.; Yamashita, S.; Matsuzawa, Y.; Brewer, H.B., Jr. Exon 10 skipping caused by intron 10 splice donor site mutation in cholesteryl ester transfer protein gene results in abnormal downstream splice site selection. J. Lipid Res. 1996, 37, 2065–2073. [Google Scholar] [PubMed]
- Lira, M.E.; Loomis, A.K.; Paciga, S.A.; Lloyd, D.B.; Thompson, J.F. Expression of CETP and of splice variants induces the same level of ER stress despite secretion efficiency differences. J. Lipid Res. 2008, 49, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Izem, L.; Greene, D.J.; Bialkowska, K.; Morton, R.E. Overexpression of full-length cholesteryl ester transfer protein in SW872 cells reduces lipid accumulation. J. Lipid Res. 2015, 56, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Izem, L.; Morton, R.E. Possible role for intracellular cholesteryl ester transfer protein in adipocyte lipid metabolism and storage. J. Biol. Chem. 2007, 282, 21856–21865. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Ayers, S.; Milanesi, A.; Teng, X.; Rabi, S.; Akiba, Y.; Brent, G.A. Thyroid hormone receptor sumoylation is required for preadipocyte differentiation and proliferation. J. Biol. Chem. 2015, 290, 7402–7415. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Feng, T.; Zhu, N.; Liu, P.; Han, X.; Chen, M.; Wang, X.; Li, N.; Li, Y.; Xu, Y.; et al. Identification of a novel selective agonist of PPARγ with no promotion of adipogenesis and less inhibition of osteoblastogenesis. Sci. Rep. 2015, 5, 9530. [Google Scholar] [CrossRef] [PubMed]
- Zadegan, F.G.; Ghaedi, K.; Kalantar, S.M.; Peymani, M.; Hashemi, M.S.; Baharvand, H.; Nasr-Esfahani, M.H. Cardiac differentiation of mouse embryonic stem cells is influenced by a PPAR γ/PGC-1α-FNDC5 pathway during the stage of cardiac precursor cell formation. Eur. J. Cell Biol. 2015, 94, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Yates, T.; Davies, M.J.; Henson, J.; Edwardson, C.; Webb, D.; Bodicoat, D.H.; Webb, M.; Howard, P.; Cooper, J.A.; Humphries, S.E.; et al. Effect of the PPARG2 Pro12Ala Polymorphism on Associations of Physical Activity and Sedentary Time with Markers of Insulin Sensitivity in Those with an Elevated Risk of Type 2 Diabetes. PLoS ONE 2015, 10, e0124062. [Google Scholar] [CrossRef] [PubMed]
- De Silva, T.M.; Modrick, M.L.; Ketsawatsomkron, P.; Lynch, C.; Chu, Y.; Pelham, C.J.; Sigmund, C.D.; Faraci, F.M. Role of peroxisome proliferator-activated receptor-γ in vascular muscle in the cerebral circulation. Hypertension 2014, 64, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- McClelland, S.; Shrivastava, R.; Medh, J.D. Regulation of Translational Efficiency by Disparate 5′ UTRs of PPARγ Splice Variants. PPAR Res. 2009, 2009, 193413. [Google Scholar] [CrossRef] [PubMed]
- Norazmi, M.N.; Mohamed, R.; Nurul, A.A.; Yaacob, N.S. The modulation of PPARγ1 and PPARγ2 mRNA expression by ciglitazone in CD3/CD28-activated naïve and memory CD4+ T cells. Clin. Dev. Immunol. 2012, 2012, 849195. [Google Scholar] [CrossRef] [PubMed]
- Servera, M.; López, N.; Serra, F.; Palou, A. Expression of “brown-in-white” adipocyte biomarkers shows gender differences and the influence of early dietary exposure. Genes Nutr. 2014, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Elsen, M.; Raschke, S.; Tennagels, N.; Schwahn, U.; Jelenik, T.; Roden, M.; Romacho, T.; Eckel, J. BMP4 and BMP7 induce the white-to-brown transition of primary human adipose stem cells. Am. J. Physiol. Cell Physiol. 2014, 306, C431–C440. [Google Scholar] [CrossRef] [PubMed]
- Arany, Z.; He, H.; Lin, J.; Hoyer, K.; Handschin, C.; Toka, O.; Ahmad, F.; Matsui, T.; Chin, S.; Wu, P.H.; et al. Transcriptional coactivator PGC-1 α controls the energy state and contractile function of cardiac muscle. Cell Metab. 2005, 1, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Lustig, Y.; Ruas, J.L.; Estall, J.L.; Lo, J.C.; Devarakonda, S.; Laznik, D.; Choi, J.H.; Ono, H.; Olsen, J.V.; Spiegelman, B.M. Separation of the gluconeogenic and mitochondrial functions of PGC-1α through S6 kinase. Genes Dev. 2011, 25, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.S.; Fernand, V.; Zhang, Y.; Shin, J.; Jun, H.J.; Joshi, Y.; Gettys, T.W. NT-PGC-1α protein is sufficient to link β3-adrenergic receptor activation to transcriptional and physiological components of adaptive thermogenesis. J. Biol. Chem. 2012, 287, 9100–9111. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Wu, J.; Chang, J.S.; Zhang, P.; Wang, J.; Zhang, Y.; Gettys, T.W.; Zhang, Y. Effect of exercise intensity on isoform-specific expressions of NT-PGC-1 α mRNA in mouse skeletal muscle. Biomed. Res. Int. 2014, 2014, 402175. [Google Scholar] [CrossRef] [PubMed]
- Fiermonte, G.; Dolce, V.; Palmieri, L.; Ventura, M.; Runswick, M.J.; Palmieri, F.; Walker, J.E. Identification of the human mitochondrial oxodicarboxylate carrier. Bacterial expression, reconstitution, functional characterization, tissue distribution, and chromosomal location. J. Biol. Chem. 2001, 276, 8225–8230. [Google Scholar] [CrossRef] [PubMed]
- Niimi, M.; Tao, L.; Lin, S.H.; Yin, J.; Wu, X.; Fukui, H.; Kambayashi, J.; Ye, J.; Sun, B. Involvement of an alternatively spliced mitochondrial oxodicarboxylate carrier in adipogenesis in 3T3-L1 cells. J. Biomed. Sci. 2009, 16, 92. [Google Scholar] [CrossRef] [PubMed]
- Pyronnet, S.; Pradayrol, L.; Sonenberg, N. Alternative splicing facilitates internal ribosome entry on the ornithine decarboxylase mRNA. Cell Mol. Life Sci. 2005, 62, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Saltzman, A.L.; Kim, Y.K.; Misquitta, C.; Shai, O.; Maquat, L.E.; Frey, B.J.; Blencowe, B.J. Quantitative microarray profiling provides evidence against widespread coupling of alternative splicing with nonsense-mediated mRNA decay to control gene expression. Genes Dev. 2006, 20, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.; Shiue, L.; Hayes, J.D.; Centers, R.; Fitzwater, S.; Loewen, R.; Edmondson, L.R.; Bryant, J.; Smith, M.; Rommelfanger, C.; et al. Detection and measurement of alternative splicing using splicing-sensitive microarrays. Methods 2005, 37, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Tsafou, K.; Stolte, C.; Pletscher-Frankild, S.; O’Donoghue, S.I.; Jensen, L.J. Comprehensive comparison of large-scale tissue expression datasets. Peer J. 2015, 3, e1054. [Google Scholar] [CrossRef] [PubMed]
- Brosseau, J.P.; Lucier, J.F.; Lapointe, E.; Durand, M.; Gendron, D.; Gervais-Bird, J.; Tremblay, K.; Perreault, J.P.; Elela, S.A. High-throughput quantification of splicing isoforms. RNA 2010, 16, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Madgwick, A.; Fort, P.; Hanson, P.S.; Thibault, P.; Gaudreau, M.C.; Lutfalla, G.; Möröy, T.; Abou Elela, S.; Chaudhry, B.; Elliott, D.J.; et al. Neural differentiation modulates the vertebrate brain specific splicing program. PLoS ONE 2015, 10, e0125998. [Google Scholar] [CrossRef] [PubMed]
- Tokheim, C.; Park, J.W.; Xing, Y. PrimerSeq: Design and visualization of RT-PCR primers for alternative splicing using RNA-seq data. Genom. Proteom. Bioinform. 2014, 12, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Lambert, N.; Robertson, A.; Jangi, M.; McGeary, S.; Sharp, P.A.; Burge, C.B. RNA Bind-n-Seq: Quantitative assessment of the sequence and structural binding specificity of RNA binding proteins. Mol. Cell 2014, 54, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Hafner, M.; Landthaler, M.; Burger, L.; Khorshid, M.; Hausser, J.; Berninger, P.; Rothballer, A.; Ascano, M., Jr.; Jungkamp, A.C.; Munschauer, M.; et al. Transcriptome-wide identification of RNA-binding protein and microRNA target sites by PAR-CLIP. Cell 2010, 141, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Darnell, R.B. Mapping in vivo protein-RNA interactions at single-nucleotide resolution from HITS-CLIP data. Nat. Biotechnol. 2011, 29, 607–614. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, J.-C. Impacts of Alternative Splicing Events on the Differentiation of Adipocytes. Int. J. Mol. Sci. 2015, 16, 22169-22189. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160922169
Lin J-C. Impacts of Alternative Splicing Events on the Differentiation of Adipocytes. International Journal of Molecular Sciences. 2015; 16(9):22169-22189. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160922169
Chicago/Turabian StyleLin, Jung-Chun. 2015. "Impacts of Alternative Splicing Events on the Differentiation of Adipocytes" International Journal of Molecular Sciences 16, no. 9: 22169-22189. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms160922169