
The Role of MicroRNAs as Predictors of Response to Tamoxifen Treatment in Breast Cancer Patients

Abstract
:
1. Introduction
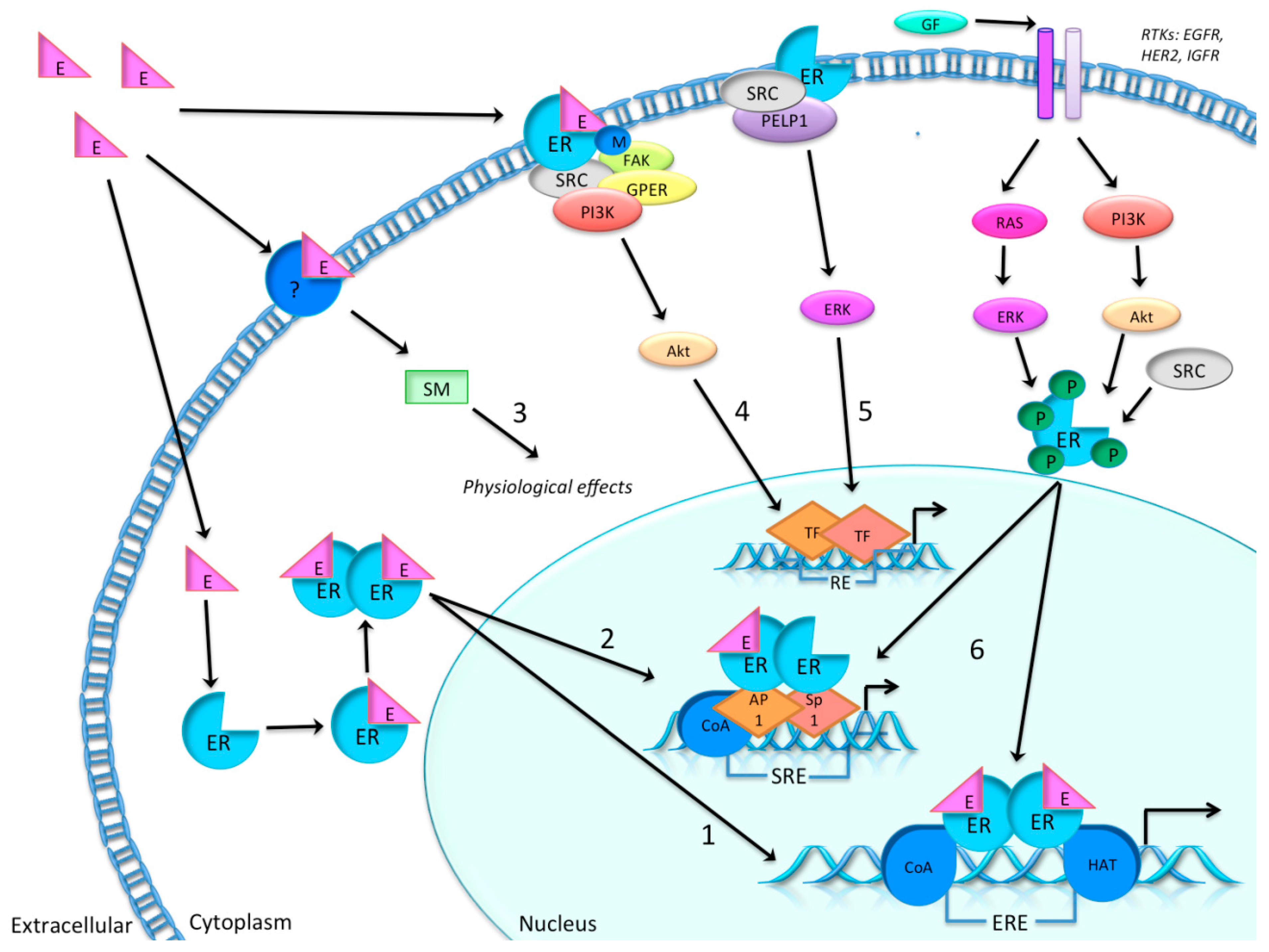
1.1. Estradiol and the Estrogen Receptor
1.2. Endocrine Treatment Regimens


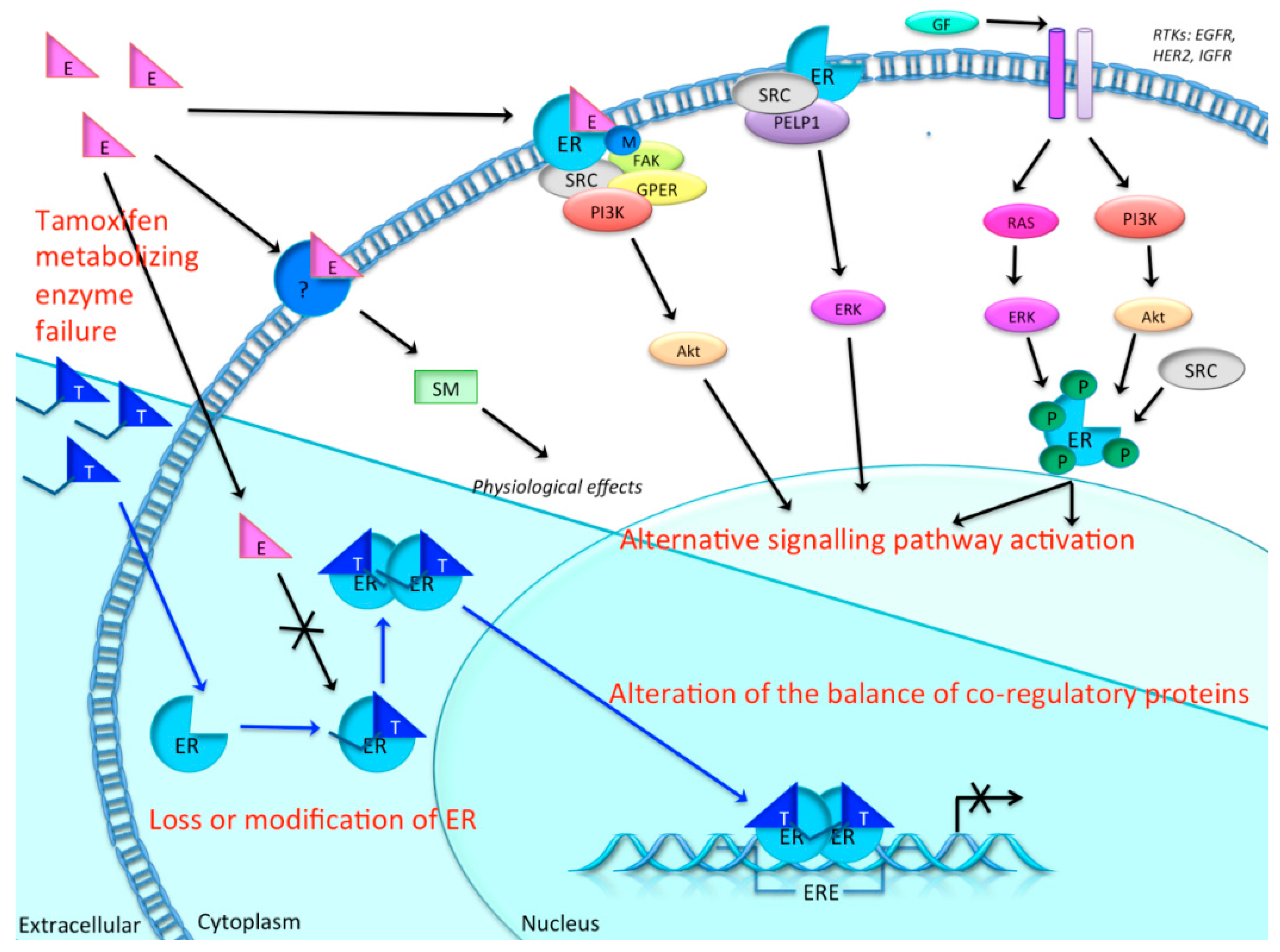
1.3. Resistance to Tamoxifen
1.4. MicroRNAs

2. Methods
2.1. Literature Search

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MicroRNA | Material and Patients | Clinical Outcome | Reference | Predicted Targets or Pathways |
|---|---|---|---|---|
| miR-342-5p | FFPE of tamoxifen-treated primary tumor, n = 16 | Ten patients responded to tamoxifen and had non-recurrent disease: two-fold the level of miR-342 expression; Six patients developed recurrences and metastasis during tamoxifen treatment and had low levels of miR-342 | [46] | Target genes: GEMIN4 and BMP7 Predicted pathways: cell death, apoptosis of breast cancer cells, mitotic roles of polo-like kinase |
| miR-30c-5p | Primary tumors, tamoxifen-treated following advanced disease, n = 246 | Increasing levels of miR-30c was associated with clinical benefit of tamoxifen treatment, as measured by longer PFS | [49] | Target genes: PPARGC1B, Makorin-3, UBAC1, PTPDC1 Predicted pathways: HER2, signal ransduction, and oncology pathway, RAC1 cell motility signaling pathway |
| miR-210-3p | Fresh-frozen tamoxifen-treated primary tumors, n = 89 | High level of miR-210 expression was associated with a higher risk of recurrence than a lower level of miR-210 | [50] | Target genes:ACVR1B, CBFA2T1 Predicted pathways: cell cycle, cell adhesion and immune response |
| miR-26a | Frozen tamoxifen-treated tumors of metastatic patients, n = 235 | Higher levels of miR-26a were significantly associated with clinical benefit (i.e., complete or partial response, or stable disease), and with favorable TTP (i.e., first detection of disease progression) | [51] | Target genes: CDC2, CCNE1 Predicted pathways: cell cycle regulation pathway |
| miR-126-5p and miR-10a-5p | FFPE from postmenopausal tamoxifen-treated patients, Validation set: n = 34 with recurrence; n = 47 without recurrence | Low expression of miR-10a and miR-126 correlated significantly with reduced relapse-free time | [47] | Target genes: n/a Predicted pathways: n/a |
| miR-519a-3p | GEO datasets of breast cancer patients, Discovery set: n = 632, Validation set: n = 586 | High expression of miRNA-519a correlated significantly with poorer disease-free survival in ER+ breast cancer patients | [48] | Target genes: PTEN, RB1, CDKN1A/p21 Predicted pathways: PI3K/Akt pathway |
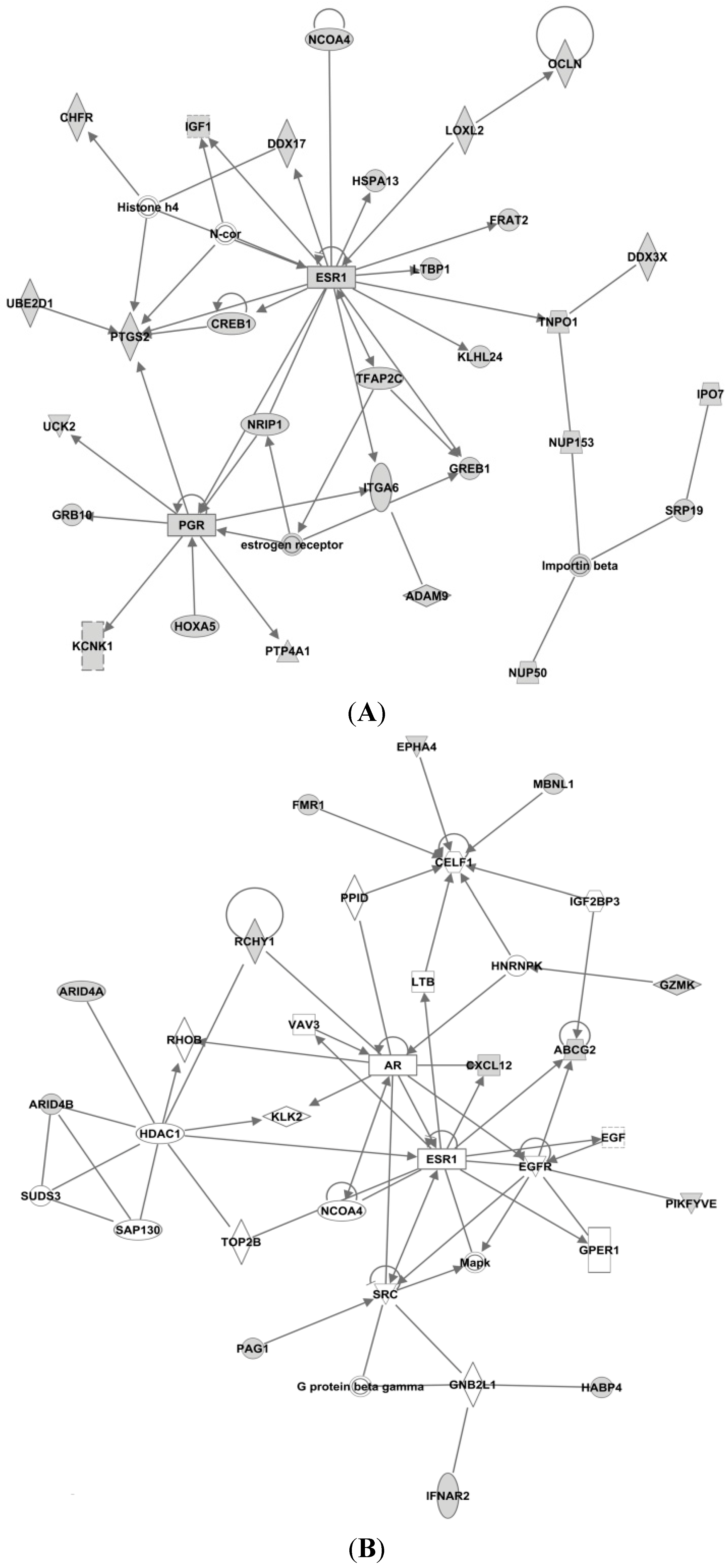
2.2. Ingenuity Pathway Analysis
| MicroRNA | No. of Target Genes |
|---|---|
| miR-342-5p | 337 |
| miR-30c-5p | 1420 |
| miR-210-3p | 78 |
| miR-26a | 892 |
| miR-126a-5p | 37 |
| miR-10a-5p | 338 |
| miR-519a-3p | 86 |
3. MicroRNAs in Breast Cancer
3.1. MicroRNAs in Breast Cancer Tumor Tissue
3.2. Circulating MicroRNAs in Breast Cancer
3.3. Tamoxifen-Related MicroRNAs Found in Breast Cancer Tissue
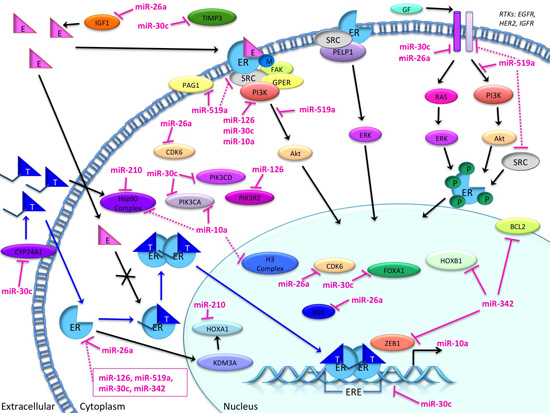
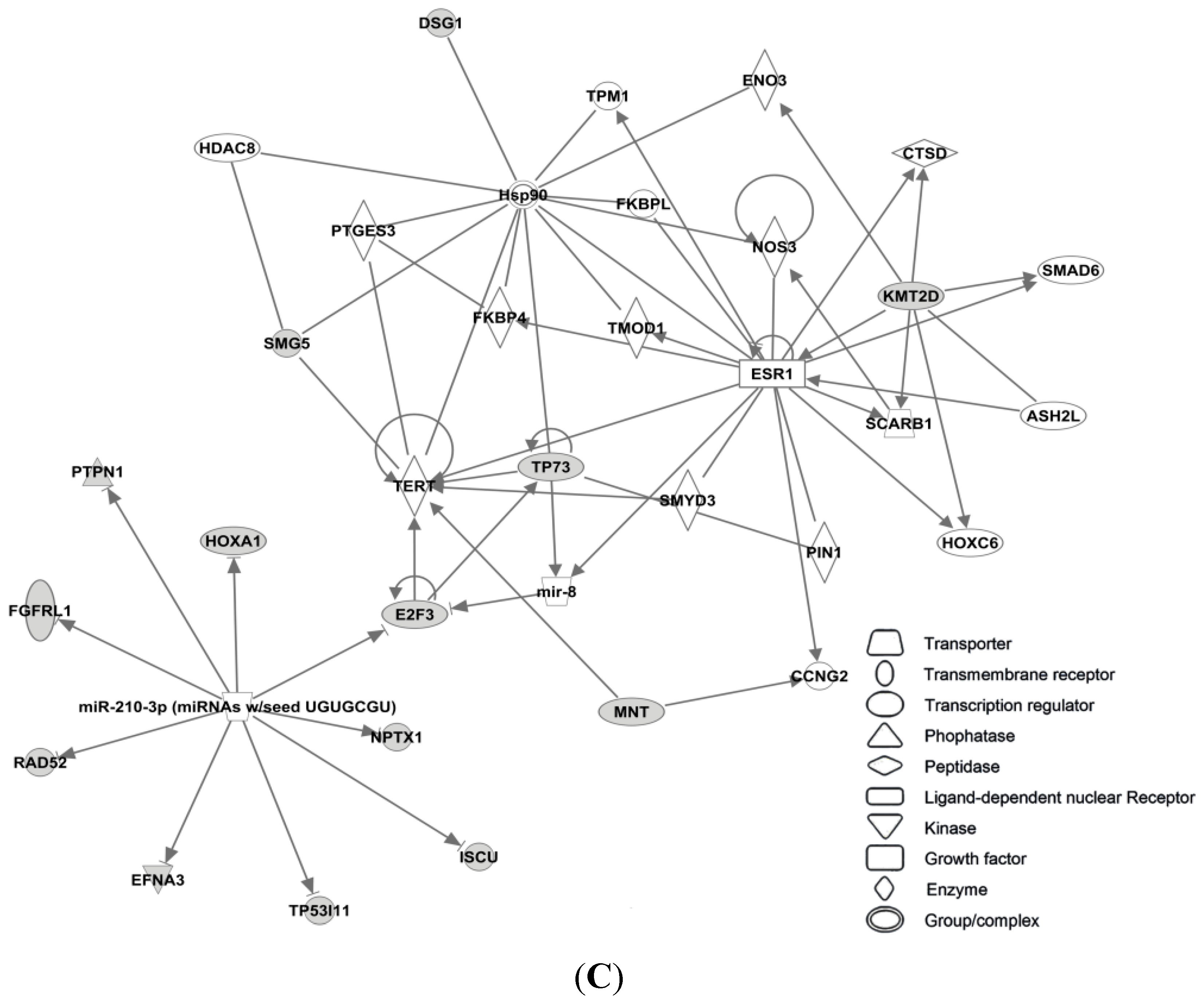
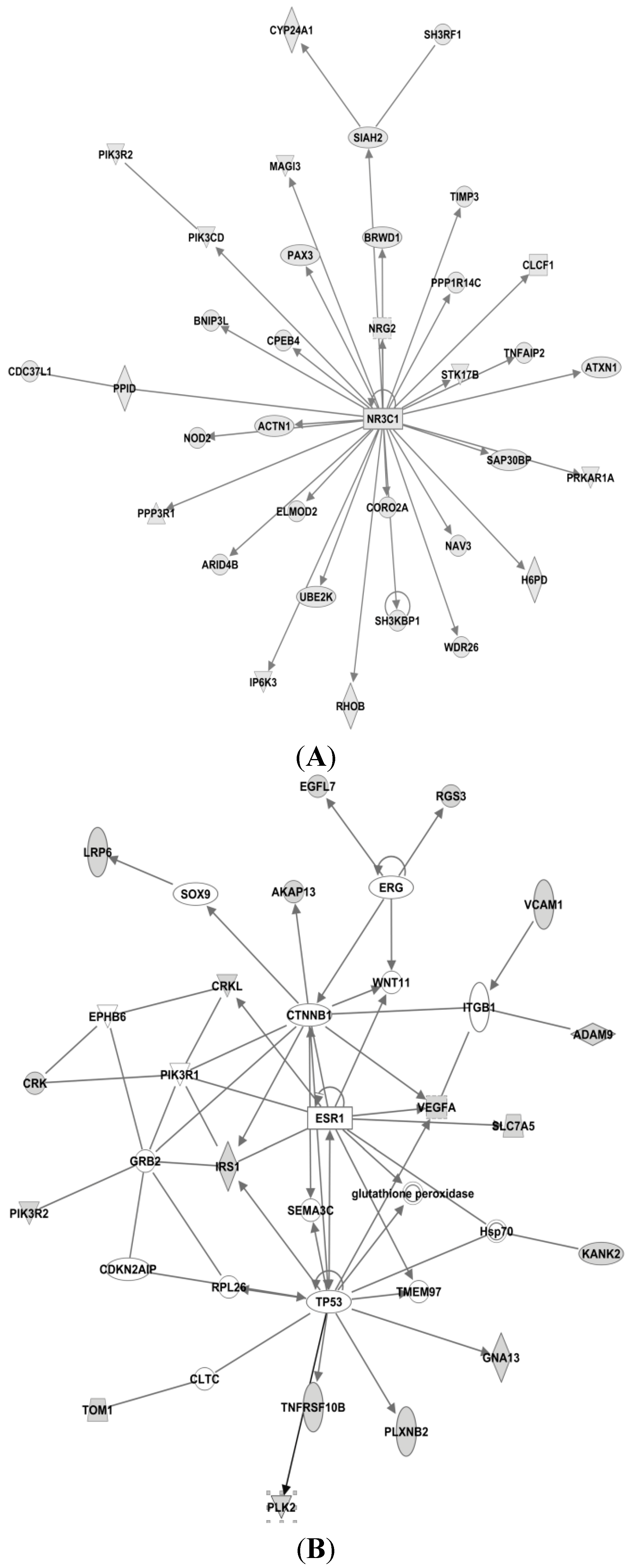
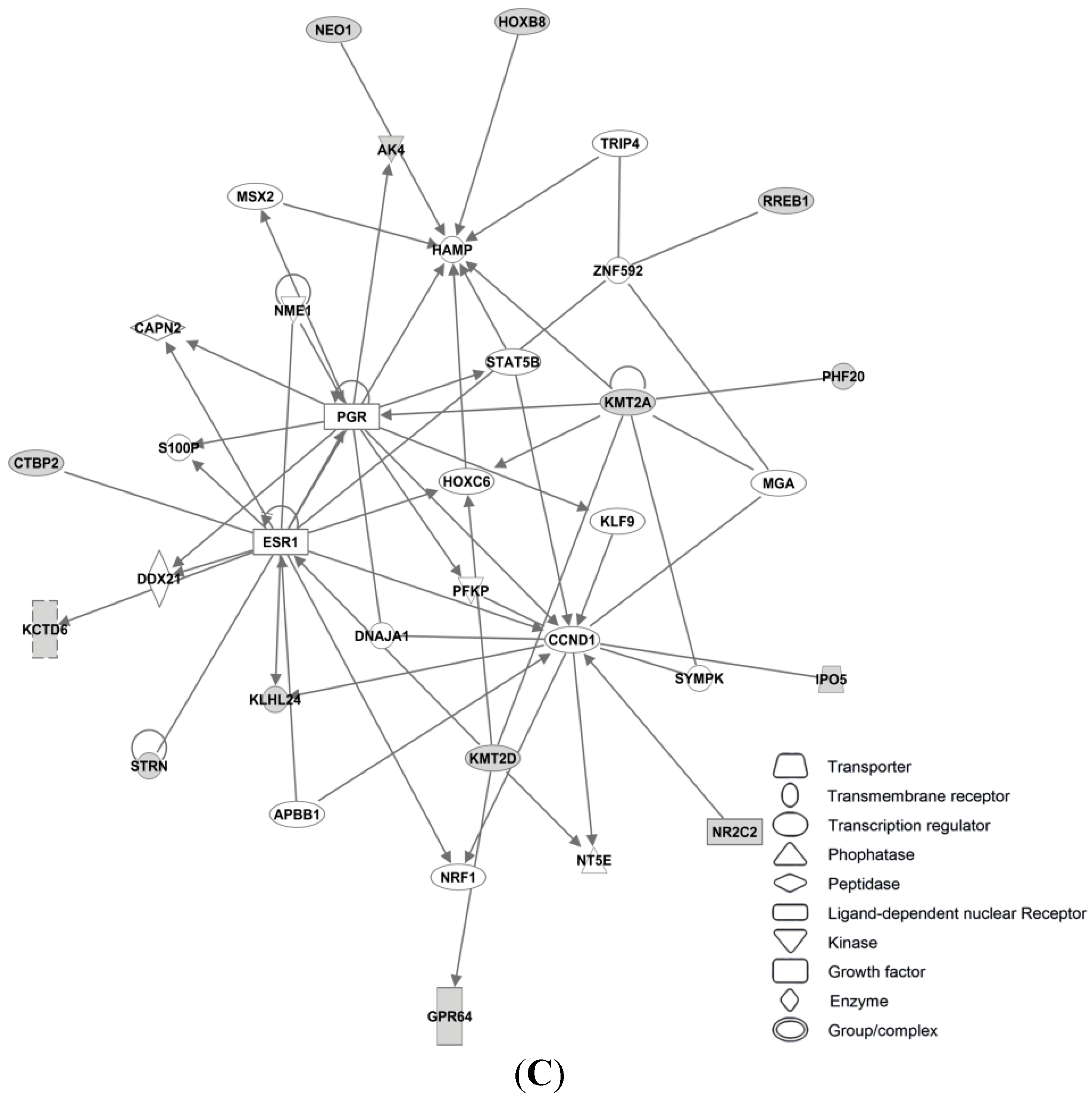
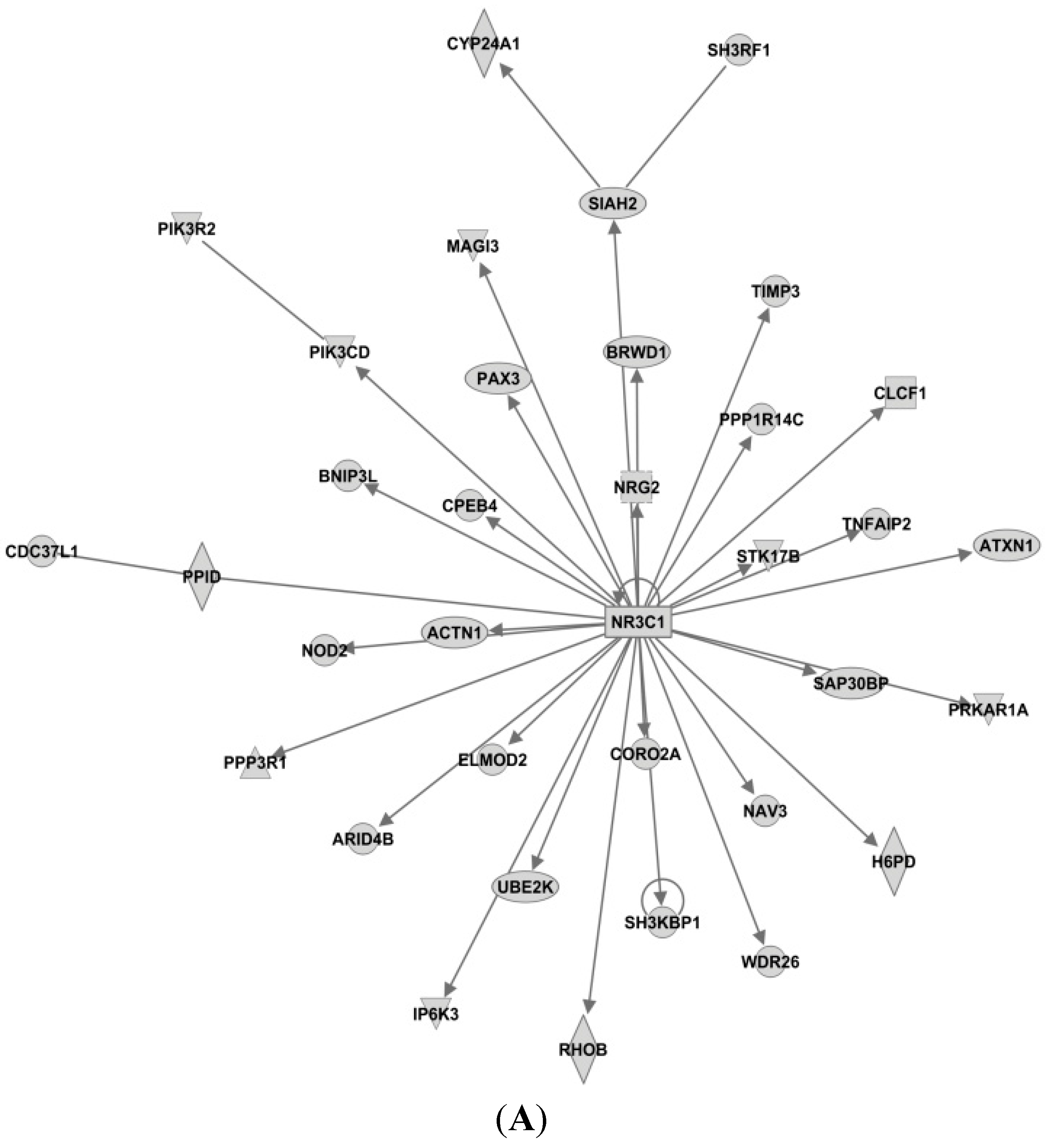
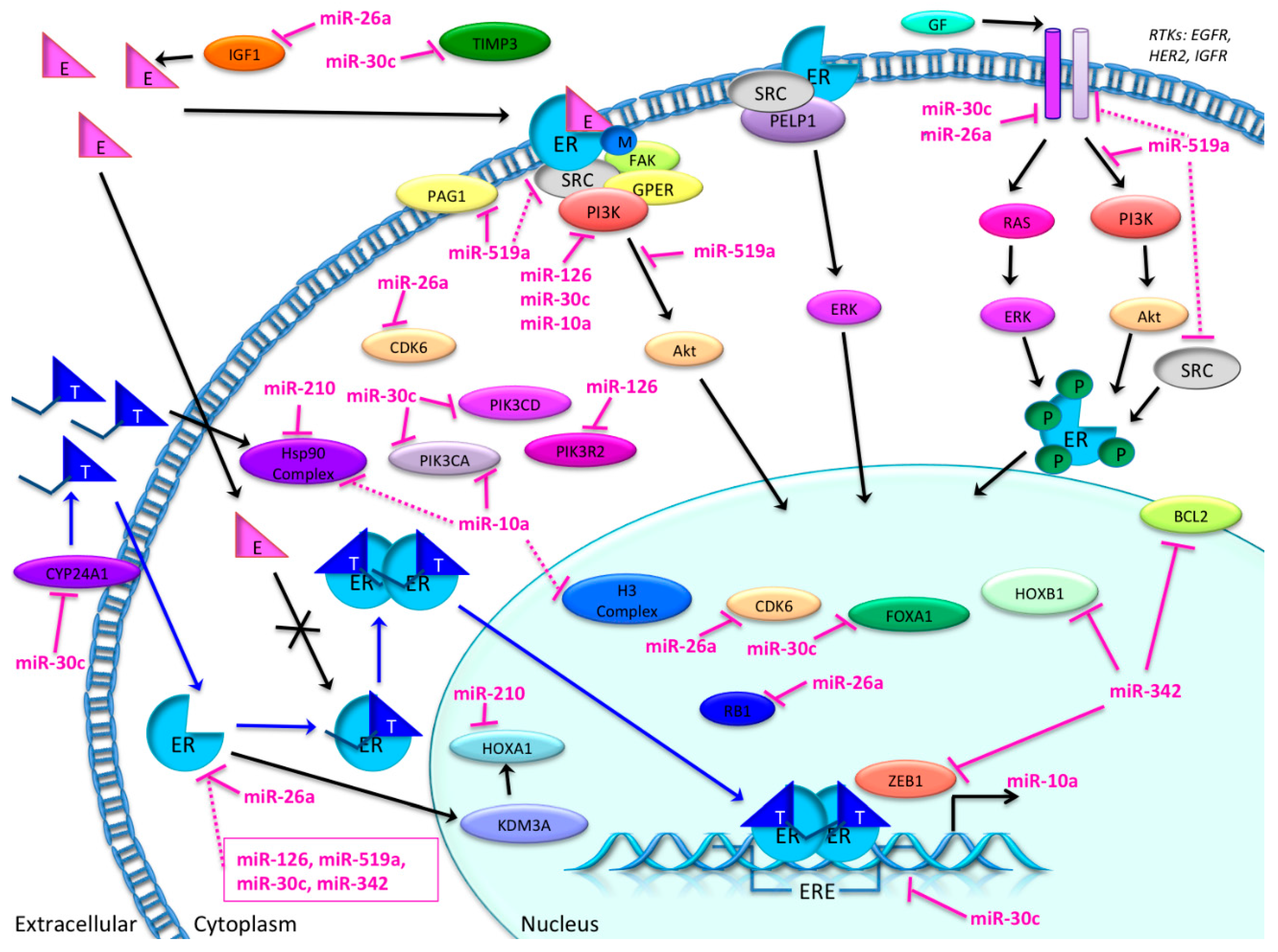
3.4. Candidate MicroRNAs in Signaling Pathways and Their Relevant Target Genes in Tamoxifen Resistance
3.4.1. Targets of miR-26a
3.4.2. Targets of miR-519
3.4.3. Targets of miR-210


3.4.4. Targets of miR-30c


3.4.5. Targets of miR-126
3.4.6. Targets of miR-342



3.4.7. Targets of miR-10a
4. Discussion

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Haynes, B.P.; Viale, G.; Galimberti, V.; Rotmensz, N.; Gibelli, B.; A'Hern, R.; Smith, I.E.; Dowsett, M. Expression of key estrogen-regulated genes differs substantially across the menstrual cycle in estrogen receptor-positive primary breast cancer. Breast Cancer Res. Treat. 2013, 138, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Baak, J.P.A.; van Diest, P.J.; Voorhorst, F.J.; van der Wall, E.; Beex, L.V.A.M.; Vermorken, J.B.; Janssen, E.A.M.; Gudlaugsson, E.; other collaborators of the Multicenter Morphometric Mammary Carcinoma Project (MMMCP). The prognostic value of proliferation in lymph-node-negative breast cancer patients is age dependent. Eur. J. Cancer 2007, 43, 527–535. [Google Scholar] [PubMed]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thurlimann, B.; Senn, H.J. Personalizing the treatment of women with early breast cancer: Highlights of the St gallen international expert consensus on the primary therapy of early breast cancer 2013. Ann. Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef] [PubMed]
- Cuzick, J.; Dowsett, M.; Pineda, S.; Wale, C.; Salter, J.; Quinn, E.; Zabaglo, L.; Mallon, E.; Green, A.R.; Ellis, I.O.; et al. Prognostic value of a combined estrogen receptor, progesterone receptor, ki-67, and human epidermal growth factor receptor 2 immunohistochemical score and comparison with the genomic health recurrence score in early breast cancer. J. Clin. Oncol. 2011, 29, 4273–4278. [Google Scholar] [CrossRef] [PubMed]
- National Comprehensive Cancer Network (NCCN). Available online: http://www.nccn.org/ (accessed on 4 October 2015).
- Early Breast Cancer Trialists’ Collaborative Group. Tamoxifen for early breast cancer: An overview of the randomised trials. Early breast cancer trialists’ collaborative group. Lancet 1998, 351, 1451–1467. [Google Scholar]
- Early Breast Cancer Trialists’ Collaborative Group. Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: An overview of the randomised trials. Lancet 2005, 365, 1687–1717. [Google Scholar]
- Carlson, R.W.; Allred, D.C.; Anderson, B.O.; Burstein, H.J.; Carter, W.B.; Edge, S.B.; Erban, J.K.; Farrar, W.B.; Forero, A.; Giordano, S.H.; et al. Invasive breast cancer, clinical practice guidelines, NCCN. J. Natl. Compr. Cancer Netw. 2011, 9, 136–222. [Google Scholar]
- Burstein, H.J.; Prestrud, A.A.; Seidenfeld, J.; Anderson, H.; Buchholz, T.A.; Davidson, N.E.; Gelmon, K.E.; Giordano, S.H.; Hudis, C.A.; Malin, J.; et al. American society of clinical oncology clinical practice guideline: Update on adjuvant endocrine therapy for women with hormone receptor-positive breast cancer. J. Clin. Oncol. 2010, 28, 3784–3796. [Google Scholar] [CrossRef] [PubMed]
- Gnant, M.; Harbeck, N.; Thomssen, C. St. Gallen 2011: Summary of the consensus discussion. Breast Care 2011, 6, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Musgrove, E.A.; Sutherland, R.L. Biological determinants of endocrine resistance in breast cancer. Nat. Rev. Cancer 2009, 9, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Dos Anjos Pultz, B.; da Luz, F.A.; de Faria, P.R.; Oliveira, A.P.; de Araujo, R.A.; Silva, M.J. Far beyond the usual biomarkers in breast cancer: A review. J. Cancer 2014, 5, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian micrornas predominantly act to decrease target mrna levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Haggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [PubMed]
- Paech, K.; Webb, P.; Kuiper, G.G.; Nilsson, S.; Gustafsson, J.; Kushner, P.J.; Scanlan, T.S. Differential ligand activation of estrogen receptors eralpha and erbeta at ap1 sites. Science 1997, 277, 1508–1510. [Google Scholar] [CrossRef] [PubMed]
- Russo, J.; Russo, I.H. Genotoxicity of steroidal estrogens. Trends Endocrinol. Metab. 2004, 15, 211–214. [Google Scholar] [CrossRef]
- Tsai, M.J.; O’Malley, B.W. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu. Rev. Biochem. 1994, 63, 451–486. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.; Pan, H.; Godwin, J.; Gray, R.; Arriagada, R.; Raina, V.; Abraham, M.; Alencar, V.H.M.; Badran, A.; Bonfill, X.; et al. Long-term effects of continuing adjuvant tamoxifen to 10 years versus stopping at 5 years after diagnosis of estrogen receptor-positive breast cancer: Atlas, a randomised trial. Lancet 2013, 381, 805–816. [Google Scholar] [CrossRef]
- Hayes, E.L.; Lewis-Wambi, J.S. Mechanisms of endocrine resistance in breast cancer: An overview of the proposed roles of noncoding RNA. Breast Cancer Res. 2015, 17, 40. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.; Costantino, J.P.; Wickerham, D.L.; Cecchini, R.S.; Cronin, W.M.; Robidoux, A.; Bevers, T.B.; Kavanah, M.T.; Atkins, J.N.; Margolese, R.G.; et al. Tamoxifen for the prevention of breast cancer: Current status of the national surgical adjuvant breast and bowel project p-1 study. J. Natl. Cancer Inst. 2005, 97, 1652–1662. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.A.; Saccani-Jotti, G.; Clarke, R.; Johnston, S.R.; Anderson, E.; Howell, A.; A’Hern, R.; Salter, J.; Detre, S.; Nicholson, R.; et al. Induction of apoptosis by tamoxifen and ici 182780 in primary breast cancer. Int. J. Cancer 1997, 72, 608–613. [Google Scholar] [CrossRef]
- Deligdisch, L.; Kalir, T.; Cohen, C.J.; de Latour, M.; Le Bouedec, G.; Penault-Llorca, F. Endometrial histopathology in 700 patients treated with tamoxifen for breast cancer. Gynecol. Oncol. 2000, 78, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Strom, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [PubMed]
- Norwegian Breast Cancer Group (NBCG). Available online: http://nbcg.no/ (accessed on 4 October 2015).
- Nass, N.; Kalinski, T. Tamoxifen resistance: From cell culture experiments towards novel biomarkers. Pathology 2015, 211, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, J.M.; Carey, L.A.; McLeod, H.L. Cyp2d6 and tamoxifen: DNA matters in breast cancer. Nat. Rev. Cancer 2009, 9, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, M.; Sedmak, D.; Jewell, S. Effect of fixatives and tissue processing on the content and integrity of nucleic acids. Am. J. Pathol. 2002, 161, 1961–1971. [Google Scholar] [CrossRef]
- Gjerde, J.; Geisler, J.; Lundgren, S.; Ekse, D.; Varhaug, J.E.; Mellgren, G.; Steen, V.M.; Lien, E.A. Associations between tamoxifen, estrogens, and fsh serum levels during steady state tamoxifen treatment of postmenopausal women with breast cancer. BMC Cancer 2010, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Cronin-Fenton, D.P.; Damkier, P.; Lash, T.L. Metabolism and transport of tamoxifen in relation to its effectiveness: New perspectives on an ongoing controversy. Future Oncol. 2014, 10, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Lien, E.A.; Soiland, H.; Lundgren, S.; Aas, T.; Steen, V.M.; Mellgren, G.; Gjerde, J. Serum concentrations of tamoxifen and its metabolites increase with age during steady-state treatment. Breast Cancer Res. Treat. 2013, 141, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Geisler, J.; Lonning, P.E. Aromatase inhibition: Translation into a successful therapeutic approach. Clin. Cancer Res. 2005, 11, 2809–2821. [Google Scholar] [CrossRef] [PubMed]
- Dowsett, M.; Jones, A.; Johnston, S.R.; Jacobs, S.; Trunet, P.; Smith, I.E. In vivo measurement of aromatase inhibition by letrozole (cgs 20267) in postmenopausal patients with breast cancer. Clin. Cancer Res. 1995, 1, 1511–1515. [Google Scholar] [PubMed]
- Geisler, J.; Haynes, B.; Anker, G.; Dowsett, M.; Lonning, P.E. Influence of letrozole and anastrozole on total body aromatization and plasma estrogen levels in postmenopausal breast cancer patients evaluated in a randomized, cross-over study. J. Clin. Oncol. 2002, 20, 751–757. [Google Scholar] [CrossRef]
- Forbes, J.F.; Cuzick, J.; Buzdar, A.; Howell, A.; Tobias, J.S.; Baum, M. Effect of anastrozole and tamoxifen as adjuvant treatment for early-stage breast cancer: 100-month analysis of the atac trial. Lancet Oncol. 2008, 9, 45–53. [Google Scholar] [PubMed]
- Mouridsen, H.; Giobbie-Hurder, A.; Goldhirsch, A.; Thurlimann, B.; Paridaens, R.; Smith, I.; Mauriac, L.; Forbes, J.; Price, K.N.; Regan, M.M.; et al. Letrozole therapy alone or in sequence with tamoxifen in women with breast cancer. N. Engl. J. Med. 2009, 361, 766–776. [Google Scholar] [PubMed]
- Osborne, C.K.; Schiff, R. Mechanisms of endocrine resistance in breast cancer. Annu. Rev. Med. 2011, 62, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Viedma-Rodriguez, R.; Baiza-Gutman, L.; Salamanca-Gomez, F.; Diaz-Zaragoza, M.; Martinez-Hernandez, G.; Ruiz Esparza-Garrido, R.; Velazquez-Flores, M.A.; Arenas-Aranda, D. Mechanisms associated with resistance to tamoxifen in estrogen receptor-positive breast cancer (review). Oncol. Rep. 2014, 32, 3–15. [Google Scholar] [PubMed]
- Garcia-Becerra, R.; Santos, N.; Diaz, L.; Camacho, J. Mechanisms of resistance to endocrine therapy in breast cancer: Focus on signaling pathways, mirnas and genetically based resistance. Int. J. Mol. Sci. 2012, 14, 108–145. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.R. Biological rationale for endocrine therapy in breast cancer. Best Pract. Res. Clin. Endocrinol. Metab. 2004, 18, 1–32. [Google Scholar] [CrossRef]
- Mirbase: The Microrna Database. Available online: http://www.mirbase.org/index.shtml (accessed on 4 October 2015).
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. Mirbase: Microrna sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. Mirbase: Annotating high confidence micrornas using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- rna genes mir15 and mir16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microrna genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. Micrornas as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cittelly, D.M.; Das, P.M.; Spoelstra, N.S.; Edgerton, S.M.; Richer, J.K.; Thor, A.D.; Jones, F.E. Downregulation of mir-342 is associated with tamoxifen resistant breast tumors. Mol. Cancer 2010, 9, 317. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, R.; Achinger-Kawecka, J.; Winter, S.; Fritz, P.; Lo, W.Y.; Schroth, W.; Brauch, H. Increased expression of mir-126 and mir-10a predict prolonged relapse-free time of primary estrogen receptor-positive breast cancer following tamoxifen treatment. Eur. J. Cancer 2013, 49, 3598–3608. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.; Shukla, K.; Balwierz, A.; Soons, Z.; Konig, R.; Sahin, O.; Wiemann, S. Microrna-519a is a novel oncomir conferring tamoxifen resistance by targeting a network of tumour-suppressor genes in ER+ breast cancer. J. Pathol. 2014, 233, 368–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Gonzalez, F.G.; Sieuwerts, A.M.; Smid, M.; Look, M.P.; Meijer-van Gelder, M.E.; de Weerd, V.; Sleijfer, S.; Martens, J.W.; Foekens, J.A. Microrna-30c expression level is an independent predictor of clinical benefit of endocrine therapy in advanced estrogen receptor positive breast cancer. Breast Cancer Res. Treat. 2011, 127, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Rothe, F.; Ignatiadis, M.; Chaboteaux, C.; Haibe-Kains, B.; Kheddoumi, N.; Majjaj, S.; Badran, B.; Fayyad-Kazan, H.; Desmedt, C.; Harris, A.L.; et al. Global microrna expression profiling identifies mir-210 associated with tumor proliferation, invasion and poor clinical outcome in breast cancer. PLoS ONE 2011, 6, e20980. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.P.; Reijm, E.A.; Sieuwerts, A.M.; Ruigrok-Ritstier, K.; Look, M.P.; Rodriguez-Gonzalez, F.G.; Heine, A.A.; Martens, J.W.; Sleijfer, S.; Foekens, J.A.; et al. High mir-26a and low cdc2 levels associate with decreased ezh2 expression and with favorable outcome on tamoxifen in metastatic breast cancer. Breast Cancer Res. Treat. 2012, 133, 937–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Schooneveld, E.; Wildiers, H.; Vergote, I.; Vermeulen, P.B.; Dirix, L.Y.; van Laere, S.J. Dysregulation of micrornas in breast cancer and their potential role as prognostic and predictive biomarkers in patient management. Breast Cancer Res. 2015, 17, 21. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. Microrna gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [PubMed]
- Sempere, L.F.; Christensen, M.; Silahtaroglu, A.; Bak, M.; Heath, C.V.; Schwartz, G.; Wells, W.; Kauppinen, S.; Cole, C.N. Altered microrna expression confined to specific epithelial cell subpopulations in breast cancer. Cancer Res. 2007, 67, 11612–11620. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.X.; Huang, X.F.; Shao, Q.; Huang, M.Y.; Deng, L.; Wu, Q.L.; Zeng, Y.X.; Shao, J.Y. Microrna mir-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosis. RNA 2008, 14, 2348–2360. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.X.; Wu, Q.N.; Zhang, Y.; Li, Y.Y.; Liao, D.Z.; Hou, J.H.; Fu, J.; Zeng, M.S.; Yun, J.P.; Wu, Q.L.; et al. Knockdown of mir-21 in human breast cancer cell lines inhibits proliferation, in vitro migration and in vivo tumor growth. Breast Cancer Res. 2011, 13, R2. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bian, C.; Liao, L.; Li, J.; Zhao, R.C. Mir-17–5p promotes human breast cancer cell migration and invasion through suppression of hbp1. Breast Cancer Res. Treat. 2011, 126, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Leivonen, S.K.; Makela, R.; Ostling, P.; Kohonen, P.; Haapa-Paananen, S.; Kleivi, K.; Enerly, E.; Aakula, A.; Hellstrom, K.; Sahlberg, N.; et al. Protein lysate microarray analysis to identify micrornas regulating estrogen receptor signaling in breast cancer cell lines. Oncogene 2009, 28, 3926–3936. [Google Scholar] [CrossRef] [PubMed]
- Jonsdottir, K.; Janssen, S.R.; Da Rosa, F.C.; Gudlaugsson, E.; Skaland, I.; Baak, J.P.; Janssen, E.A. Validation of expression patterns for nine mirnas in 204 lymph-node negative breast cancers. PLoS ONE 2012, 7, e48692. [Google Scholar] [CrossRef] [PubMed]
- Calvano Filho, C.M.; Calvano-Mendes, D.C.; Carvalho, K.C.; Maciel, G.A.; Ricci, M.D.; Torres, A.P.; Filassi, J.R.; Baracat, E.C. Triple-negative and luminal a breast tumors: Differential expression of mir-18a-5p, mir-17–5p, and mir-20a-5p. Tumour Biol. 2014, 35, 7733–7741. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.; Moller, C.; Jirstrom, K.; Lee, A.; Busch, S.; Lamb, R.; Landberg, G. Downregulation of mir-92a is associated with aggressive breast cancer features and increased tumour macrophage infiltration. PLoS ONE 2012, 7, e36051. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.L.; Schmittgen, T.D.; et al. Detection of microrna expression in human peripheral blood microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [PubMed]
- Stuckrath, I.; Rack, B.; Janni, W.; Jager, B.; Pantel, K.; Schwarzenbach, H. Aberrant plasma levels of circulating mir-16, mir-107, mir-130a and mir-146a are associated with lymph node metastasis and receptor status of breast cancer patients. Oncotarget 2015, 6, 13387. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.; Rack, B.; Muller, V.; Janni, W.; Pantel, K.; Schwarzenbach, H. Circulating microRNAs as blood-based markers for patients with primary and metastatic breast cancer. Breast Cancer Res. 2010, 12, R90. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, D.; Zucknick, M.; Wallwiener, M.; Cuk, K.; Modugno, C.; Scharpff, M.; Schott, S.; Heil, J.; Turchinovich, A.; Yang, R.; et al. Circulating mirnas as surrogate markers for circulating tumor cells and prognostic markers in metastatic breast cancer. Clin. Cancer Res. 2012, 18, 5972–5982. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Sharples, R.A.; Scicluna, B.J.; Hill, A.F. Exosomes provide a protective and enriched source of mirna for biomarker profiling compared to intracellular and cell-free blood. J. Extracell. Vesicles 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Yoshioka, Y.; Hagiwara, K.; Tominaga, N.; Ochiya, T. Functional analysis of exosomal microrna in cell-cell communication research. Methods Mol. Biol. 2013, 1024, 1–10. [Google Scholar] [PubMed]
- Taylor, D.D.; Black, P.H. Shedding of plasma membrane fragments. Neoplastic and developmental importance. Dev. Biol. 1986, 3, 33–57. [Google Scholar]
- Zhao, L.; Liu, W.; Xiao, J.; Cao, B. The role of exosomes and “exosomal shuttle microrna” in tumorigenesis and drug resistance. Cancer Lett. 2015, 356, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.X.; Liu, X.M.; Lv, M.M.; Chen, L.; Zhao, J.H.; Zhong, S.L.; Ji, M.H.; Hu, Q.; Luo, Z.; Wu, J.Z.; et al. Exosomes from drug-resistant breast cancer cells transmit chemoresistance by a horizontal transfer of micrornas. PLoS ONE 2014, 9, e95240. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lai, X.; Yu, S.; Chen, S.; Ma, Y.; Zhang, Y.; Li, H.; Zhu, X.; Yao, L.; Zhang, J. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res. Treat. 2014, 147, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Reijm, E.A.; Jansen, M.P.; Ruigrok-Ritstier, K.; van Staveren, I.L.; Look, M.P.; van Gelder, M.E.; Sieuwerts, A.M.; Sleijfer, S.; Foekens, J.A.; Berns, E.M. Decreased expression of ezh2 is associated with upregulation of ER and favorable outcome to tamoxifen in advanced breast cancer. Breast Cancer Res. Treat. 2011, 125, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.; Bentley, J.; Wang, L.Z.; Newell, D.R.; Robson, C.N.; Shapiro, G.I.; Curtin, N.J. Pre-clinical evaluation of cyclin-dependent kinase 2 and 1 inhibition in anti-estrogen-sensitive and resistant breast cancer cells. Br. J. Cancer 2010, 102, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Bosco, E.E.; Wang, Y.; Xu, H.; Zilfou, J.T.; Knudsen, K.E.; Aronow, B.J.; Lowe, S.W.; Knudsen, E.S. The retinoblastoma tumor suppressor modifies the therapeutic response of breast cancer. J. Clin. Investig. 2007, 117, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; Le Roith, D. Insulin-like growth factor 1 and oestradiol promote cell proliferation of mcf-7 breast cancer cells: New insights into their synergistic effects. Mol. Pathol. 2001, 54, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Riedemann, J.; Macaulay, V.M. Igf1r signalling and its inhibition. Endocr. Relat. Cancer 2006, 13, S33–S43. [Google Scholar] [CrossRef] [PubMed]
- Fagan, D.H.; Yee, D. Crosstalk between igf1r and estrogen receptor signaling in breast cancer. J. Mammary Gland Biol. Neoplasia 2008, 13, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Sharon, E. Igf-1r as an anti-cancer target—Trials and tribulations. Chin. J. Cancer 2013, 32, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Planas-Silva, M.D.; Hamilton, K.N. Targeting c-Src kinase enhances tamoxifen’s inhibitory effect on cell growth by modulating expression of cell cycle and survival proteins. Cancer Chemother. Pharmacol. 2007, 60, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Morgan, L.; Gee, J.; Pumford, S.; Farrow, L.; Finlay, P.; Robertson, J.; Ellis, I.; Kawakatsu, H.; Nicholson, R.; Hiscox, S. Elevated src kinase activity attenuates tamoxifen response in vitro and is associated with poor prognosis clinically. Cancer Biol. Ther. 2009, 8, 1550–1558. [Google Scholar] [CrossRef] [PubMed]
- Larsen, S.L.; Laenkholm, A.V.; Duun-Henriksen, A.K.; Bak, M.; Lykkesfeldt, A.E.; Kirkegaard, T. Src drives growth of antiestrogen resistant breast cancer cell lines and is a marker for reduced benefit of tamoxifen treatment. PLoS ONE 2015, 10, e0118346. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, A.; Ignatov, T.; Weissenborn, C.; Eggemann, H.; Bischoff, J.; Semczuk, A.; Roessner, A.; Costa, S.D.; Kalinski, T. G-protein-coupled estrogen receptor gpr30 and tamoxifen resistance in breast cancer. Breast Cancer Res. Treat. 2011, 128, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Liu, M.; Yang, L.; Tu, G.; Zhu, Q.; Chen, M.; Cheng, H.; Luo, H.; Fu, W.; Li, Z.; et al. Acquisition of epithelial-mesenchymal transition phenotype in the tamoxifen-resistant breast cancer cell: A new role for g protein-coupled estrogen receptor in mediating tamoxifen resistance through cancer-associated fibroblast-derived fibronectin and beta1-integrin signaling pathway in tumor cells. Breast Cancer Res. 2015, 17, 69. [Google Scholar] [PubMed]
- Mahajan, K.; Lawrence, H.R.; Lawrence, N.J.; Mahajan, N.P. Ack1 tyrosine kinase interacts with histone demethylase kdm3a to regulate the mammary tumor oncogene hoxa1. J. Biol. Chem. 2014, 289, 28179–28191. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, K.; Mahajan, N.P. Ack1/tnk2 tyrosine kinase: Molecular signaling and evolving role in cancers. Oncogene 2014, 34, 4162–4167. [Google Scholar] [CrossRef] [PubMed]
- Lianos, G.D.; Alexiou, G.A.; Mangano, A.; Mangano, A.; Rausei, S.; Boni, L.; Dionigi, G.; Roukos, D.H. The role of heat shock proteins in cancer. Cancer Lett. 2015, 360, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Leung, E.; Gruner, S.; Schapira, M.; Houry, W.A. Tamoxifen enhances the hsp90 molecular chaperone atpase activity. PLoS ONE 2010, 5, e9934. [Google Scholar] [CrossRef] [PubMed]
- Whitesell, L.; Santagata, S.; Mendillo, M.L.; Lin, N.U.; Proia, D.A.; Lindquist, S. Hsp90 empowers evolution of resistance to hormonal therapy in human breast cancer models. Proc. Natl. Acad. Sci. USA 2014, 111, 18297–18302. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.; Holmes, K.A.; Ross-Innes, C.S.; Schmidt, D.; Carroll, J.S. Foxa1 is a critical determinant of estrogen receptor function and endocrine response. Nat. Genet. 2011, 43, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ross-Innes, C.S.; Stark, R.; Teschendorff, A.E.; Holmes, K.A.; Ali, H.R.; Dunning, M.J.; Brown, G.D.; Gojis, O.; Ellis, I.O.; Green, A.R.; et al. Differential oestrogen receptor binding is associated with clinical outcome in breast cancer. Nature 2012, 481, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, K.D.; Brown, G.R.; Hiatt, S.M.; Thibaud-Nissen, F.; Astashyn, A.; Ermolaeva, O.; Farrell, C.M.; Hart, J.; Landrum, M.J.; McGarvey, K.M.; et al. Refseq: An update on mammalian reference sequences. Nucleic Acids Res. 2014, 42, D756–D763. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, J.; Yde, C.W.; Lykkesfeldt, A.E. 1alpha,25-dihydroxyvitamin d3 inhibits cell growth and nfkappab signaling in tamoxifen-resistant breast cancer cells. Steroids 2014, 85, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Gan, R.; Yang, Y.; Yang, X.; Zhao, L.; Lu, J.; Meng, Q.H. Downregulation of mir-221/222 enhances sensitivity of breast cancer cells to tamoxifen through upregulation of timp3. Cancer Gene Ther. 2014, 21, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Vogt, P.K. Class i pi3k in oncogenic cellular transformation. Oncogene 2008, 27, 5486–5496. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase (pi3k) pathway in cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Sabine, V.S.; Crozier, C.; Brookes, C.L.; Drake, C.; Piper, T.; van de Velde, C.J.; Hasenburg, A.; Kieback, D.G.; Markopoulos, C.; Dirix, L.; et al. Mutational analysis of pi3k/akt signaling pathway in tamoxifen exemestane adjuvant multinational pathology study. J. Clin. Oncol. 2014, 32, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.W.; Rexer, B.N.; Garrett, J.T.; Arteaga, C.L. Mutations in the phosphatidylinositol 3-kinase pathway: Role in tumor progression and therapeutic implications in breast cancer. Breast Cancer Res. 2011, 13, 224. [Google Scholar] [CrossRef] [PubMed]
- Juric, D.; Castel, P.; Griffith, M.; Griffith, O.L.; Won, H.H.; Ellis, H.; Ebbesen, S.H.; Ainscough, B.J.; Ramu, A.; Iyer, G.; et al. Convergent loss of pten leads to clinical resistance to a pi(3)kalpha inhibitor. Nature 2015, 518, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Ujihira, T.; Ikeda, K.; Suzuki, T.; Yamaga, R.; Sato, W.; Horie-Inoue, K.; Shigekawa, T.; Osaki, A.; Saeki, T.; Okamoto, K.; et al. Microrna-574–3p, identified by microrna library-based functional screening, modulates tamoxifen response in breast cancer. Sci. Rep. 2015, 5, 7641. [Google Scholar] [CrossRef] [PubMed]
- Manavalan, T.T.; Teng, Y.; Litchfield, L.M.; Muluhngwi, P.; Al-Rayyan, N.; Klinge, C.M. Reduced expression of mir-200 family members contributes to antiestrogen resistance in ly2 human breast cancer cells. PLoS ONE 2013, 8, e62334. [Google Scholar] [CrossRef] [PubMed]
- Cittelly, D.M.; Das, P.M.; Salvo, V.A.; Fonseca, J.P.; Burow, M.E.; Jones, F.E. Oncogenic her2 delta 16 suppresses mir-15a/16 and deregulates bcl-2 to promote endocrine resistance of breast tumors. Carcinogenesis 2010, 31, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- Lash, T.L.; Lien, E.A.; Sorensen, H.T.; Hamilton-Dutoit, S. Genotype-guided tamoxifen therapy: Time to pause for reflection? Lancet Oncol. 2009, 10, 825–833. [Google Scholar] [CrossRef]
- Lonning, P.E. Molecular basis for therapy resistance. Mol. Oncol. 2010, 4, 284–300. [Google Scholar] [CrossRef] [PubMed]
- Russnes, H.G.; Navin, N.; Hicks, J.; Borresen-Dale, A.L. Insight into the heterogeneity of breast cancer through next-generation sequencing. J. Clin. Investig. 2011, 121, 3810–3818. [Google Scholar] [CrossRef] [PubMed]
- Flageng, M.H.; Moi, L.L.; Dixon, J.M.; Geisler, J.; Lien, E.A.; Miller, W.R.; Lonning, P.E.; Mellgren, G. Nuclear receptor co-activators and her-2/neu are upregulated in breast cancer patients during neo-adjuvant treatment with aromatase inhibitors. Br. J. Cancer 2009, 101, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Moi, L.L.; Flageng, M.H.; Gjerde, J.; Madsen, A.; Rost, T.H.; Gudbrandsen, O.A.; Lien, E.A.; Mellgren, G. Steroid receptor coactivators, her-2 and her-3 expression is stimulated by tamoxifen treatment in dmba-induced breast cancer. BMC Cancer 2012, 12, 247. [Google Scholar] [CrossRef] [PubMed]
- Creighton, C.J.; Fu, X.; Hennessy, B.T.; Casa, A.J.; Zhang, Y.; Gonzalez-Angulo, A.M.; Lluch, A.; Gray, J.W.; Brown, P.H.; Hilsenbeck, S.G.; et al. Proteomic and transcriptomic profiling reveals a link between the pi3k pathway and lower estrogen-receptor (ER) levels and activity in ER+ breast cancer. Breast Cancer Res. 2010, 12, R40. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Creighton, C.J.; Biswal, N.C.; Kumar, V.; Shea, M.; Herrera, S.; Contreras, A.; Gutierrez, C.; Wang, T.; Nanda, S.; et al. Overcoming endocrine resistance due to reduced pten levels in estrogen receptor-positive breast cancer by co-targeting mammalian target of rapamycin, protein kinase b, or mitogen-activated protein kinase kinase. Breast Cancer Res. 2014, 16, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, J.N.; Loo, S.Y.; Datta, A.; Siveen, K.S.; Yap, W.N.; Cai, W.; Shin, E.M.; Wang, C.; Kim, J.E.; Chan, M.; et al. Micrornas in breast cancer: Regulatory roles governing the hallmarks of cancer. Biol. Rev. Camb. Philos. Soc. 2015. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Galasso, M.; Sana, M.E.; Wise, T.F.; Palatini, J.; Huebner, K.; Croce, C.M. Breast cancer signatures for invasiveness and prognosis defined by deep sequencing of microRNA. Proc. Natl. Acad. Sci. USA 2012, 109, 3024–3029. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Ding, K.; Li, R.; Zhang, W.; Li, G.; Kong, X.; Qian, P.; Lobie, P.E.; Zhu, T. Identification of mir-26 as a key mediator of estrogen stimulated cell proliferation by targeting chd1, greb1 and kpna2. Breast Cancer Res. 2014, 16, R40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Du, Y.Y.; Lin, Y.F.; Chen, Y.T.; Yang, L.; Wang, H.J.; Ma, D. The cell growth suppressor, mir-126, targets irs-1. Biochem. Biophys. Res. Commun. 2008, 377, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Paik, S.; Shak, S.; Tang, G.; Kim, C.; Baker, J.; Cronin, M.; Baehner, F.L.; Walker, M.G.; Watson, D.; Park, T.; et al. A multigene assay to predict recurrence of tamoxifen-treated, node-negative breast cancer. N. Engl. J. Med. 2004, 351, 2817–2826. [Google Scholar] [CrossRef] [PubMed]
- Albain, K.S.; Paik, S.; van’t Veer, L. Prediction of adjuvant chemotherapy benefit in endocrine responsive, early breast cancer using multigene assays. Breast 2009, 18, S141–S145. [Google Scholar] [CrossRef]
- Engstrom, M.J.; Opdahl, S.; Hagen, A.I.; Romundstad, P.R.; Akslen, L.A.; Haugen, O.A.; Vatten, L.J.; Bofin, A.M. Molecular subtypes, histopathological grade and survival in a historic cohort of breast cancer patients. Breast Cancer Res. Treat. 2013, 140, 463–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Chang, J.T.; Gwin, W.R.; Zhu, J.; Ambs, S.; Geradts, J.; Lyerly, H.K. A signature of epithelial-mesenchymal plasticity and stromal activation in primary tumor modulates late recurrence in breast cancer independent of disease subtype. Breast Cancer Res. 2014, 16, 407. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.C.; Truong, P.T.; Wai, E.S.; Nichol, A.; Weir, L.; Speers, C.; Hayes, M.M.; Baliski, C.; Tyldesley, S. Population-based analysis of the impact and generalizability of the nsabp-b24 study on endocrine therapy for patients with ductal carcinoma in situ of the breastdagger. Ann. Oncol. 2015. [Google Scholar] [CrossRef]
- Bernhard, J.; Luo, W.; Ribi, K.; Colleoni, M.; Burstein, H.J.; Tondini, C.; Pinotti, G.; Spazzapan, S.; Ruhstaller, T.; Puglisi, F.; et al. Patient-reported outcomes with adjuvant exemestane versus tamoxifen in premenopausal women with early breast cancer undergoing ovarian suppression (text and soft): A combined analysis of two phase 3 randomised trials. Lancet Oncol. 2015, 16, 848–858. [Google Scholar] [CrossRef]
- Soiland, H.; Hagen, K.B.; Gjerde, J.; Lende, T.H.; Lien, E.A. Breaking away: High fracture rates may merit a new trial of adjuvant endocrine therapy in scandinavian breast cancer patients. Acta Oncol. 2013, 52, 861–862. [Google Scholar] [CrossRef] [PubMed]
- Lintermans, A.; Neven, P. Safety of aromatase inhibitor therapy in breast cancer. Expert Opin. Drug Saf. 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egeland, N.G.; Lunde, S.; Jonsdottir, K.; Lende, T.H.; Cronin-Fenton, D.; Gilje, B.; Janssen, E.A.M.; Søiland, H. The Role of MicroRNAs as Predictors of Response to Tamoxifen Treatment in Breast Cancer Patients. Int. J. Mol. Sci. 2015, 16, 24243-24275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161024243
Egeland NG, Lunde S, Jonsdottir K, Lende TH, Cronin-Fenton D, Gilje B, Janssen EAM, Søiland H. The Role of MicroRNAs as Predictors of Response to Tamoxifen Treatment in Breast Cancer Patients. International Journal of Molecular Sciences. 2015; 16(10):24243-24275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161024243
Chicago/Turabian StyleEgeland, Nina G., Siri Lunde, Kristin Jonsdottir, Tone H. Lende, Deirdre Cronin-Fenton, Bjørnar Gilje, Emiel A. M. Janssen, and Håvard Søiland. 2015. "The Role of MicroRNAs as Predictors of Response to Tamoxifen Treatment in Breast Cancer Patients" International Journal of Molecular Sciences 16, no. 10: 24243-24275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161024243