
Adjuvant Immune Enhancement of Subunit Vaccine Encoding pSCPI of Streptococcus iniae in Channel Catfish (Ictalurus punctatus)

Abstract
:1. Introduction
2. Results
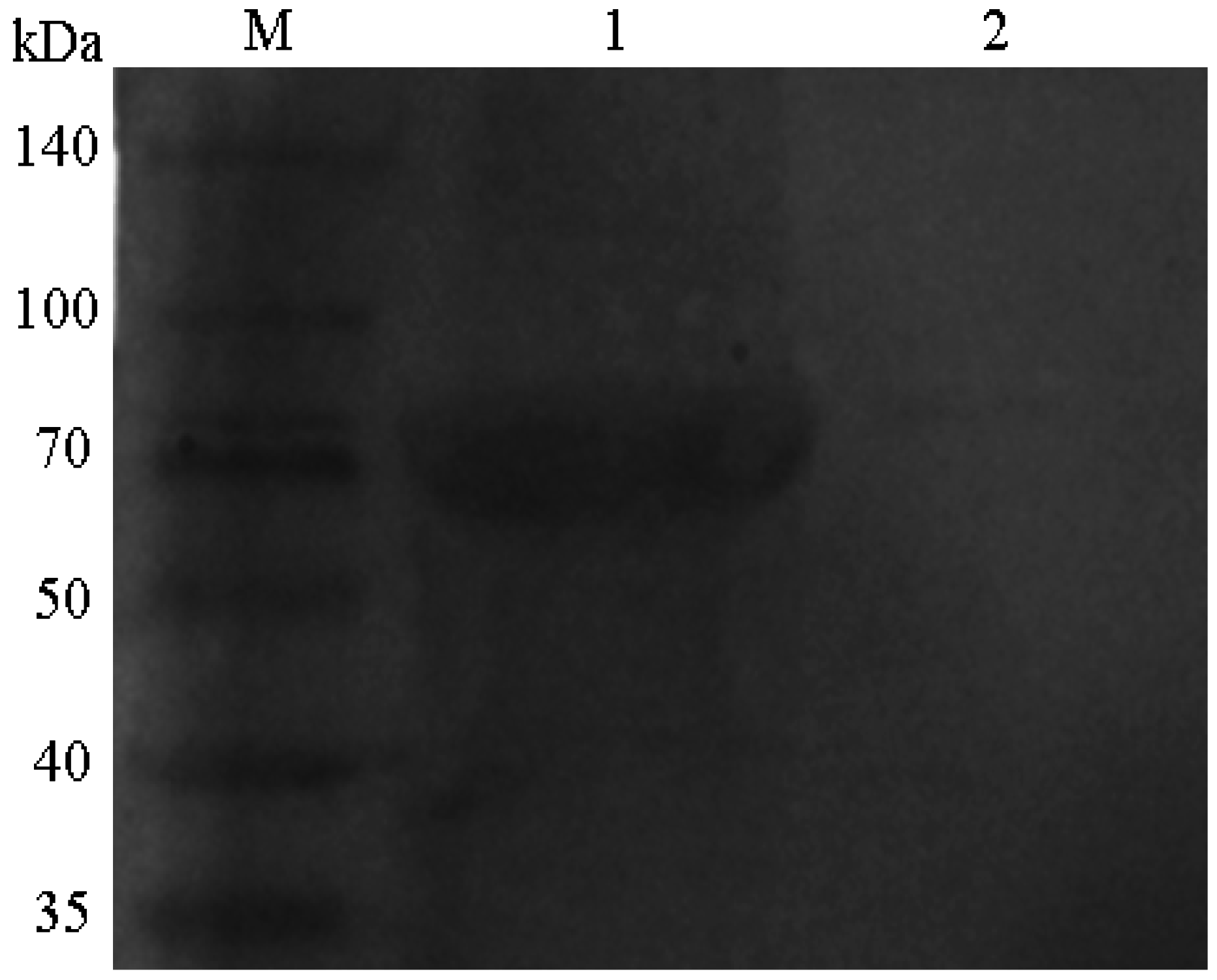
2.1. Western Blotting Analysis of the pSCPI Protein

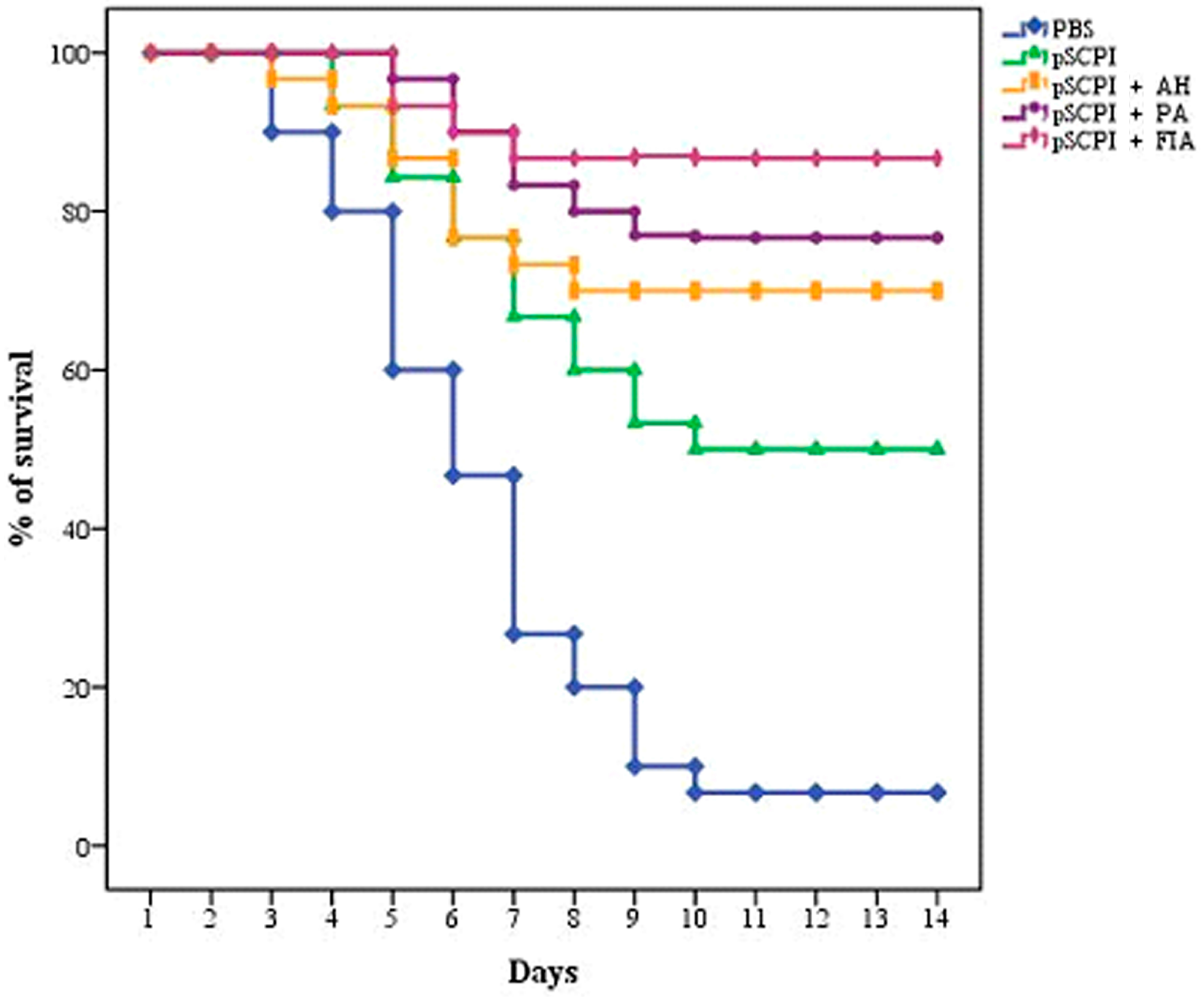
2.2. Efficacy Against the S. iniae Strain DGX07

2.3. Lysozyme Activity

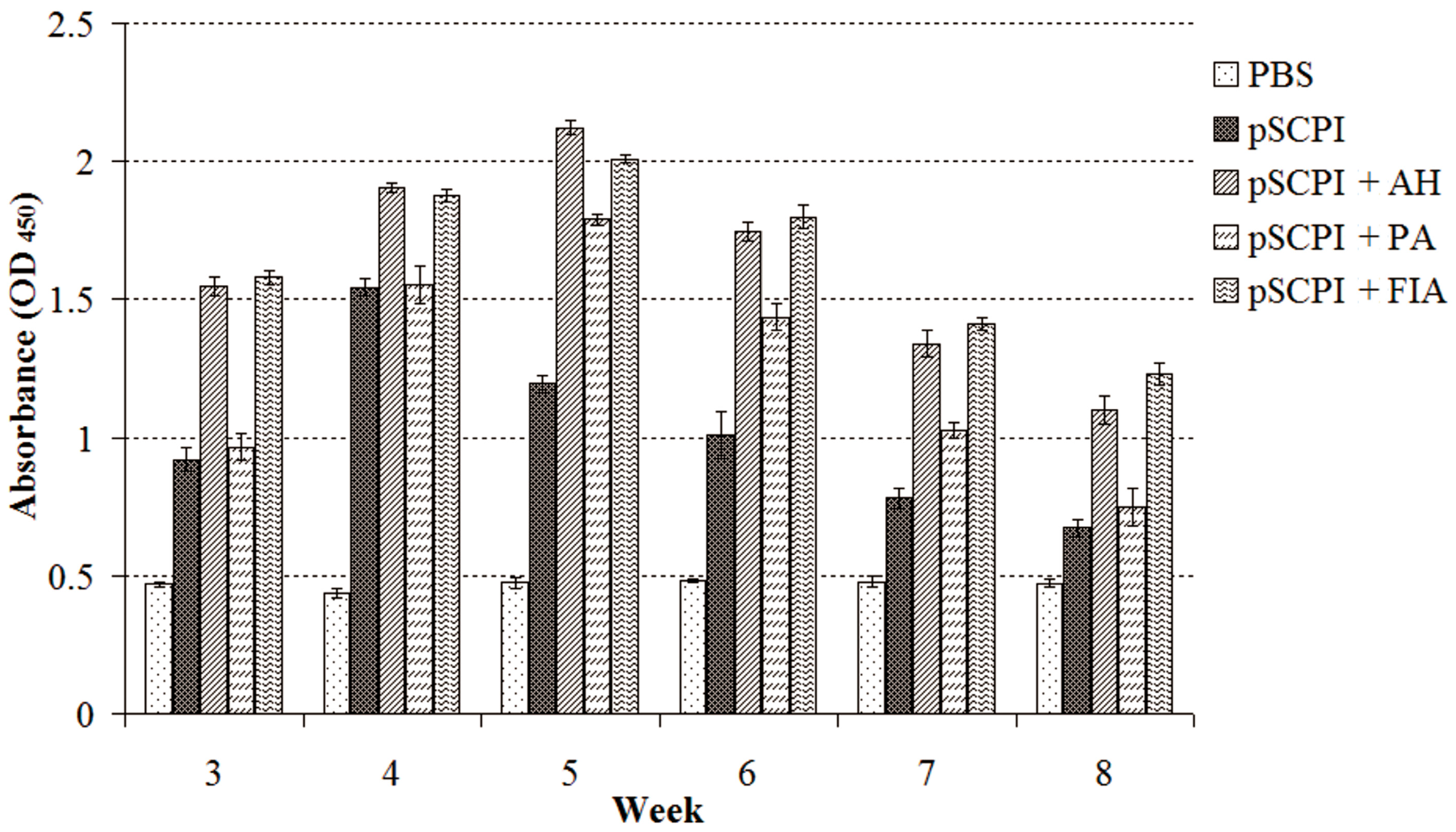
2.4. Specific Antibody Titer Detection

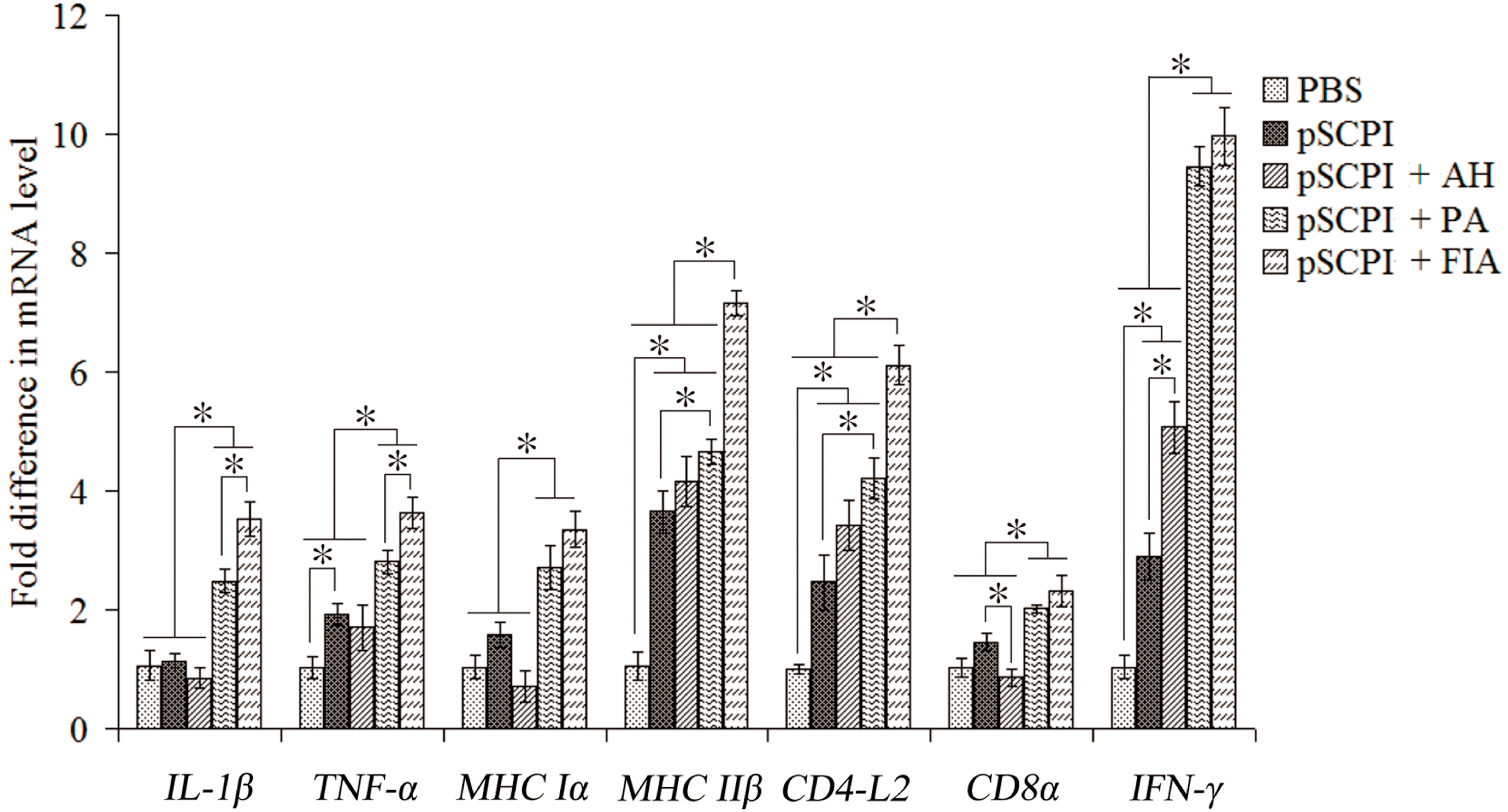
2.5. Expression of Immune-Related Genes

3. Discussion
4. Materials and Methods
4.1. Bacterial Strain, Plasmid and Growth Conditions
4.2. Animals
4.3. Expression and Purification of the pSCPI Protein
4.4. Preparation of Specific Antisera
4.5. Western Blotting
4.6. Preparation of the Adjuvant and Vaccine
4.7. Fish immunization and Challenge
4.8. Detection of Lysozyme Activity
4.9. ELISA
4.10. Quantitative Real-Time Reverse Transcription-PCR (qRT-PCR) Analysis of the Expression of Immune-Related Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences (5′→3′) | Target Gene | Product Size (bp) | GenBank |
|---|---|---|---|---|
| 18S F | GGACACGGAAAGGATTGACAGA | 18S rRNA | 121 | AF021880.1 |
| 18S R | GAGGAGTCTCGTTCGTTATCGG | 174 | DQ160229.1 | |
| IL-1β1 F | GCCATGTTGCTAATGTTGTAATCG | IL-1 β | 146 | U77598.1 |
| IL-1β1 R | TGTCTTGCAGGCTGTAACTCTTG | 129 | AJ417565.2 | |
| MHC II F | CGGGAAGGAGATTAAAGGAGGT | MHC II β | 100 | AF053547.1 |
| MHC II R | GTTTGGTGAAGCTGGCGTGT | 122 | NM_001200227.1 | |
| TNF α F | CGCACAACAAACCAGACGAGAC | TNF- α | 122 | GQ179649.1 |
| TNF α R | ACCACTGCATAGATACGCTCGAA | 100 | NM_001200217.1 | |
| MHC I F | GGTATCATCGTTGGTGTAGCCG | MHC I α | 121 | AF021880.1 |
| MHC I R | GGACAGGTTTGAAGCCAGAGTT | 174 | DQ160229.1 | |
| CD4 F | GCAGGGCACGGATAGATGGA | CD4-L2 | 146 | U77598.1 |
| CD4 R | TGGGTTCGCAGAGGCTGATAC | 129 | AJ417565.2 | |
| CD8 F | CCGACAGTGCCTACGACTAAAGC | CD8 α | 100 | AF053547.1 |
| CD8 R | CCAGCAGCCAAAGGAATGAAG | 122 | NM_001200227.1 | |
| IFN F | TGCACGAAGTGAAAGACCAAA | IFN-γ | 122 | GQ179649.1 |
| IFN R | TTAAGGTCCAGCAGCTCAGTGA | 100 | NM_001200217.1 |
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zou, J.; Carrington, A.; Collet, B.; Dijkstra, J.M.; Yoshiura, Y.; Bols, N.; Secombes, C. Identification and bioactivities of IFN-γ in rainbow trout Oncorhynchus mykiss: The first Th1-type cytokine characterized functionally in fish. J. Immunol. 2005, 175, 2484–2494. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Wang, K.; Chen, D.; Huang, X.; He, M.; Yin, Z. Stenotrophomonas maltophilia, an emerging opportunist pathogen for cultured Channel catfish, Ictalurus punctatus, in China. Aquaculture 2010, 308, 132–135. [Google Scholar] [CrossRef]
- Baiano, J.; Barnes, A.C. Towards control of Streptococcus iniae. Emerg. Infect. Dis. 2009, 15, 1891–1896. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-F.; Wang, K.-Y.; Geng, Y.; Wang, J.; Huang, L.-Y.; Li, J.-M. Streptococcus iniae isolated from Channel catfish (Ictalurus punctatus) in China. Isr. J. Aquac. Bamidgeh 2011, 63–70. [Google Scholar]
- Perera, R.P.; Johnson, S.K.; Collins, M.D.; Lewis, D.H. Streptococcus iniae associated with mortality of Tilapia nilotica × T. aurea hybrids. J. Aquat. Anim. Health 1994, 6, 335–340. [Google Scholar] [CrossRef]
- Agnew, W.; Barnes, A.C. Streptococcus iniae: An aquatic pathogen of global veterinary significance and a challenging candidate for reliable vaccination. Vet. Microbiol. 2007, 122, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.B.; Aziz, R.K.; Vicknair, M.R.; Nizet, V.; Buchanan, J.T. Streptococcus iniae M-like protein contributes to virulence in fish and is a target for live attenuated vaccine development. PLoS ONE 2008, 3, e2824. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-F.; Zhang, B.-C.; Li, J.; Sun, L. Sil: A Streptococcus iniae bacteriocin with dual role as an antimicrobial and an immunomodulator that inhibits innate immune response and promotes S. iniae infection. PLoS ONE 2014, 9, e96222. [Google Scholar] [CrossRef] [PubMed]
- Low, D.; Liu, E.; Fuller, J.; McGeer, A. Streptococcus iniae: An emerging pathogen in the aquaculture industry. Emerg. Infect. 1999, 53–66. [Google Scholar]
- He, Y.; Wang, K.-Y.; Xiao, D.; Chen, D.-F.; Huang, L.; Liu, T.; Wang, J.; Geng, Y.; Wang, E.-L.; Yang, Q. A recombinant truncated surface immunogenic protein (tSip) plus adjuvant FIA confers active protection against Group B streptococcus infection in tilapia. Vaccine 2014, 32, 7025–7032. [Google Scholar] [CrossRef] [PubMed]
- Guy, B. The perfect mix: Recent progress in adjuvant research. Nat. Rev. Microbiol. 2007, 5, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.-D.; Cheng, S.; Hu, Y.-H.; Sun, L. Comparative study of the effects of aluminum adjuvants and Freund's incomplete adjuvant on the immune response to an Edwardsiella tarda major antigen. Vaccine 2010, 28, 1832–1837. [Google Scholar] [CrossRef] [PubMed]
- Tafalla, C.; Bøgwald, J.; Dalmo, R.A. Adjuvants and immunostimulants in fish vaccines: Current knowledge and future perspectives. Fish Shellfish Immunol. 2013, 35, 1740–1750. [Google Scholar] [CrossRef] [PubMed]
- Vinay, T.-N.; Park, C.-S.; Kim, H.-Y.; Jung, S.-J. Toxicity and dose determination of quillaja saponin, aluminum hydroxide and squalene in olive flounder (Paralichthys olivaceus). Vet. Immunol. Immunopathol. 2014, 158, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Gjessing, M.C.; Falk, K.; Weli, S.C.; Koppang, E.O.; Kvellestad, A. A sequential study of incomplete Freund’s adjuvant-induced peritonitis in Atlantic cod. Fish Shellfish Immun. 2012, 32, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Yingeng, W.; Qingyin, W.; Nannan, D.; Meijie, L.; Jiangbo, Q.; Bin, L.; Lan, W. Study on the immune enhancement of different immunoadjuvants used in the pentavalent vaccine for turbots. Fish Shellfish Immunol. 2012, 32, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, A.; Rodríguez, A.; Esteban, M.A.; Meseguer, J. In vivo effects of propolis, a honeybee product, on gilthead seabream innate immune responses. Fish Shellfish Immunol. 2005, 18, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Gong, S.; Yu, D.; Yuan, H. Propolis and Herba Epimedii extracts enhance the non-specific immune response and disease resistance of Chinese sucker, Myxocyprinus asiaticus. Fish Shellfish Immunol. 2009, 26, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Yonar, M.E.; Yonar, S.M.; Silici, S. Protective effect of propolis against oxidative stress and immunosuppression induced by oxytetracycline in rainbow trout (Oncorhynchus mykiss, W.). Fish Shellfish Immunol. 2011, 31, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Bomford, R. The comparative selectivity of adjuvants for humoral and cell-mediated immunity. II. Effect on delayed-type hypersensitivity in the mouse and guinea pig, and cell-mediated immunity to tumour antigens in the mouse of Freund’s incomplete and complete adjuvants, alhydrogel, Corynebacterium parvum, Bordetella pertussis, muramyl dipeptide and saponin. Clin. Exp. Immunol. 1980, 39, 435–441. [Google Scholar] [PubMed]
- Bomford, R.; Stapleton, M.; Winsor, S.; McKnight, A.; Andronova, T. The control of the antibody isotype response to recombinant human immunodeficiency virus gp120 antigen by adjuvants. AIDS Res. Hum. Retrovir. 1992, 8, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- El Sayed, H.; Ahmad, T.A. The use of propolis as vaccine’s adjuvant. Vaccine 2012, 31, 31–39. [Google Scholar]
- Shet, A.; Kaplan, E.L.; Johnson, D.R.; Cleary, P.P. Immune response to group A streptococcal C5a peptidase in children: Implications for vaccine development. J. Infect. Dis. 2003, 188, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Cleary, P.P.; Matsuka, Y.V.; Huynh, T.; Lam, H.; Olmsted, S.B. Immunization with C5a peptidase from either group A or B streptococci enhances clearance of group A streptococci from intranasally infected mice. Vaccine 2004, 22, 4332–4341. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Debol, S.; Lam, H.; Eby, R.; Edwards, L.; Matsuka, Y.; Olmsted, S.B.; Cleary, P.P. Immunization with C5a peptidase or peptidase-type III polysaccharide conjugate vaccines enhances clearance of group B streptococci from lungs of infected mice. Infect. Immun. 2002, 70, 6409–6415. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.-L.; Wang, K.-Y.; Xiao, M.-W.; Wang, J.; Li, L.-M.; He, Y.; Chen, D.-F.; Huang, L.-Y. Fractional expression of functional domain of C5a peptidase of Strepticiccus iniae and evaluation of protective efficacy in Channel catfish (Ictalurus punctatus). Oceanol. Et Limnol. Sin. 2013, 44, 1229–1234. [Google Scholar]
- Shet, A.; Kaplan, E.; Johnson, D.; Cleary, P.P. Human immunogenicity studies on group A streptococcal C5a peptidase (SCPA) as a potential vaccine against group A streptococcal infections. Indian J. Med. Res. 2004, 119, 95–98. [Google Scholar]
- Lindblad, E.B. Aluminium adjuvants—In retrospect and prospect. Vaccine 2004, 22, 3658–3668. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-X.; Xie, Y.; Ye, Y.-P. Advances in saponin-based adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Arvouet-Grand, A.; Lejeune, B.; Bastide, P.; Pourrat, A.; Privat, A.; Legret, P. Propolis extract. I. Acute toxicity and determination of acute primary cutaneous irritation index. J. Pharm. Belg. 1992, 48, 165–170. [Google Scholar]
- Orsi, R.; Funari, S.; Soares, A.; Calvi, S.; Oliveira, S.; Sforcin, J.; Bankova, V. Immunomodulatory action of propolis on macrophage activation. J. Venom. Anim. Toxins 2000, 6, 205–219. [Google Scholar] [CrossRef]
- Sforcin, J. Propolis and the immune system: A review. J. Ethnopharmacol. 2007, 113, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vinay, T.-N.; Kim, Y.-J.; Jung, M.-H.; Kim, W.-S.; Kim, D.-H.; Jung, S.-J. Inactivated vaccine against viral hemorrhagic septicemia (VHS) emulsified with squalene and aluminum hydroxide adjuvant provides long term protection in olive flounder (Paralichthys olivaceus). Vaccine 2013, 31, 4603–4610. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.; Fletcher, T. The immunohistochemical localization of lysozyme in plaice (Pleuronectes platessa L.) tissues. J. Fish Biol. 1976, 9, 329–334. [Google Scholar] [CrossRef]
- Yousif, A.; Albright, L.; Evelyn, T. Occurrence of lysozyme in the eggs of coho salmon Oncorhynchus kisutch. Dis. Aquat. Org. 1991, 10, 45–49. [Google Scholar] [CrossRef]
- Swain, P.; Dash, S.; Sahoo, P.; Routray, P.; Sahoo, S.; Gupta, S.; Meher, P.; Sarangi, N. Non-specific immune parameters of brood Indian major carp Labeo rohita and their seasonal variations. Fish Shellfish Immunol. 2007, 22, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Gómez, G.D.; Balcázar, J.L. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol. 2008, 52, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Acosta, J.; Carpio, Y.; Valdés, I.; Velázquez, J.; Zamora, Y.; Morales, R.; Morales, A.; Rodríguez, E.; Estrada, M.P. Co-administration of tilapia α-helical antimicrobial peptides with subunit antigens boost immunogenicity in mice and tilapia (Oreochromis niloticus). Vaccine 2014, 32, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Barbu, V.; Dautry, F. Northern blot normalization with a 28S rRNA olinucleotide probe. Nucleic Acids Res. 1989, 17, 7115–7115. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, W.; Slagboom, P.; Vijg, J. Quantitative comparison of mRNA levels in mammalian tissues: 28S ribosomal RNA level as an accurate internal control. Nucleic Acids Res. 1989, 17, 10137–10138. [Google Scholar] [CrossRef] [PubMed]
- Mansur, N.R.; Meyer-Siegler, K.; Wurzer, J.C.; Sirover, M.A. Cell cycle regulation of the glyceraldehyde3phosphate dehydrogenaseluracil DNA glycosylase gene in normal human cells. Nucleic Acids Res. 1993, 21, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, P.; Taylor, W.R.; Greenberg, A.H.; Wright, J.A. Comparison of glyceraldehyde-3-phosphate dehydrogenase and 28S-ribosomal RNA gene expression as RNA loading controls for northern blot analysis of cell lines of varying malignant potential. Anal. Biochem. 1994, 216, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Small, B.C.; Murdock, C.A.; Bilodeau-Bourgeois, A.L.; Peterson, B.C.; Waldbieser, G.C. Stability of reference genes for real-time PCR analyses in Channel catfish (Ictalurus punctatus) tissues under varying physiological conditions. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Guo, Z.; Shen, Z.; Wang, J.; Hu, Y.; Wang, D. The immune enhancement of propolis adjuvant on inactivated porcine parvovirus vaccine in guinea pig. Cell Immunol. 2011, 270, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Levine, T.P.; Chain, B.M.; Brodsky, F. The cell biology of antigen processing. Crit. Rev. Biochem. Mol. 1991, 26, 439–473. [Google Scholar] [CrossRef] [PubMed]
- Boehm, U.; Klamp, T.; Groot, M.; Howard, J. Cellular responses to interferon-γ. Annu. Rev. Immunol. 1997, 15, 749–795. [Google Scholar] [CrossRef] [PubMed]
- Page, M.; Thorpe, R. Purification of IgG by precipitation with sodium sulfate or ammonium sulfate. Protein Protoc. Handb. 2002, 983–984. [Google Scholar]
- Madoff, L.C.; Michel, J.; Kasper, D. A monoclonal antibody identifies a protective C-protein α-antigen epitope in group B streptococci. Infect. Immun. 1991, 59, 204–210. [Google Scholar] [PubMed]
- Paulino, N.; Scremin, F.M.; Raichaski, L.B.; Marcucci, M.C.; Scremin, A.; Calixto, J.B. Mechanisms involved in the relaxant action of the ethanolic extract of propolis in the guinea-pig trachea in-vitro. J. Pharm. Pharmacol. 2002, 54, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Amend, D.F. Potency testing of fish vaccines. In Fish Biol. Serodiagn. Vaccines; International Association of Biological Standardization: West Virginia, WV, USA, 1981; pp. 447–454. [Google Scholar]
- Sankaran, K.; Gurnani, S. On the variation in the catalytic activity of lysozyme in fishes. Indian J. Biochem. Biol. 1972, 9, 162–165. [Google Scholar]
- Studnicka, M.; Siwicki, A.; Ryka, B. Lysozyme level in Carp (Cyrpinus-Carpio L). Bamidgeh 1986, 38, 22–25. [Google Scholar]
- Huang, L.; Wang, K.; Xiao, D.; Chen, D.; Geng, Y.; Wang, J.; He, Y.; Wang, E.; Huang, J.; Xiao, G. Safety and immunogenicity of an oral DNA vaccine encoding Sip of Streptococcus agalactiae from Nile tilapia Oreochromis niloticus delivered by live attenuated Salmonella typhimurium. Fish Shellfish Immunol. 2014, 38, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-W.; Sun, K.; Cheng, S.; Sun, L. Characterization of DegQVh, a serine protease and a protective immunogen from a pathogenic Vibrio harveyi strain. Appl. Environ. Microbiol. 2008, 74, 6254–6262. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Zheng, Z.; Wang, K.; Wang, J.; He, Y.; Wang, E.; Chen, D.; Ouyang, P.; Geng, Y.; Huang, X. Adjuvant Immune Enhancement of Subunit Vaccine Encoding pSCPI of Streptococcus iniae in Channel Catfish (Ictalurus punctatus). Int. J. Mol. Sci. 2015, 16, 28001-28013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226082
Jiang J, Zheng Z, Wang K, Wang J, He Y, Wang E, Chen D, Ouyang P, Geng Y, Huang X. Adjuvant Immune Enhancement of Subunit Vaccine Encoding pSCPI of Streptococcus iniae in Channel Catfish (Ictalurus punctatus). International Journal of Molecular Sciences. 2015; 16(12):28001-28013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226082
Chicago/Turabian StyleJiang, Jie, Zonglin Zheng, Kaiyu Wang, Jun Wang, Yang He, Erlong Wang, Defang Chen, Ping Ouyang, Yi Geng, and Xiaoli Huang. 2015. "Adjuvant Immune Enhancement of Subunit Vaccine Encoding pSCPI of Streptococcus iniae in Channel Catfish (Ictalurus punctatus)" International Journal of Molecular Sciences 16, no. 12: 28001-28013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226082