
Carbon Ion Irradiated Neural Injury Induced the Peripheral Immune Effects in Vitro or in Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
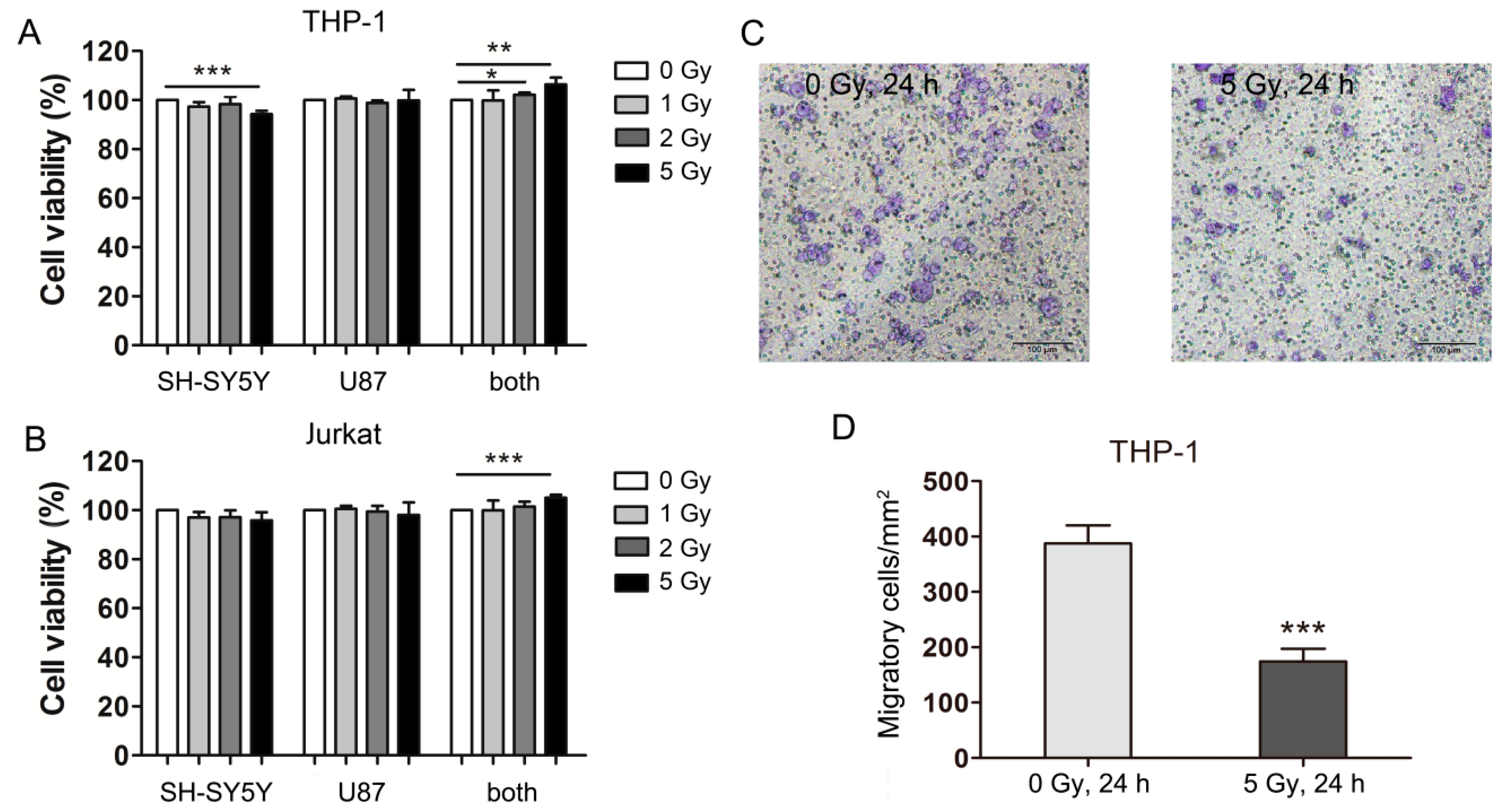
2.1. Carbon Ion-Irradiated Neural Cells Mediate Immune Effects in Vitro
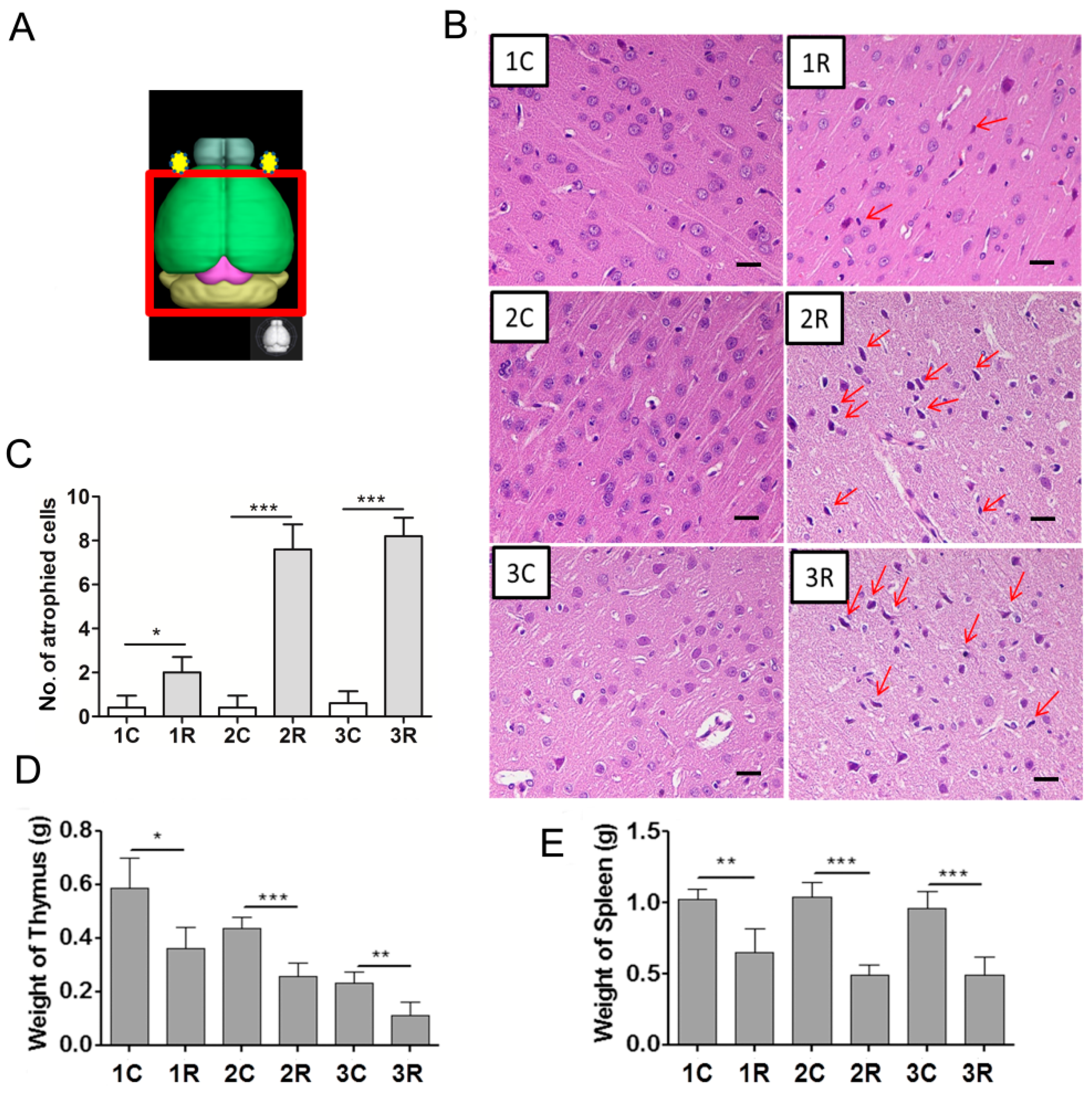
2.2. Carbon Ion-Irradiated Neural Cells Mediate Immune Effects in Vivo

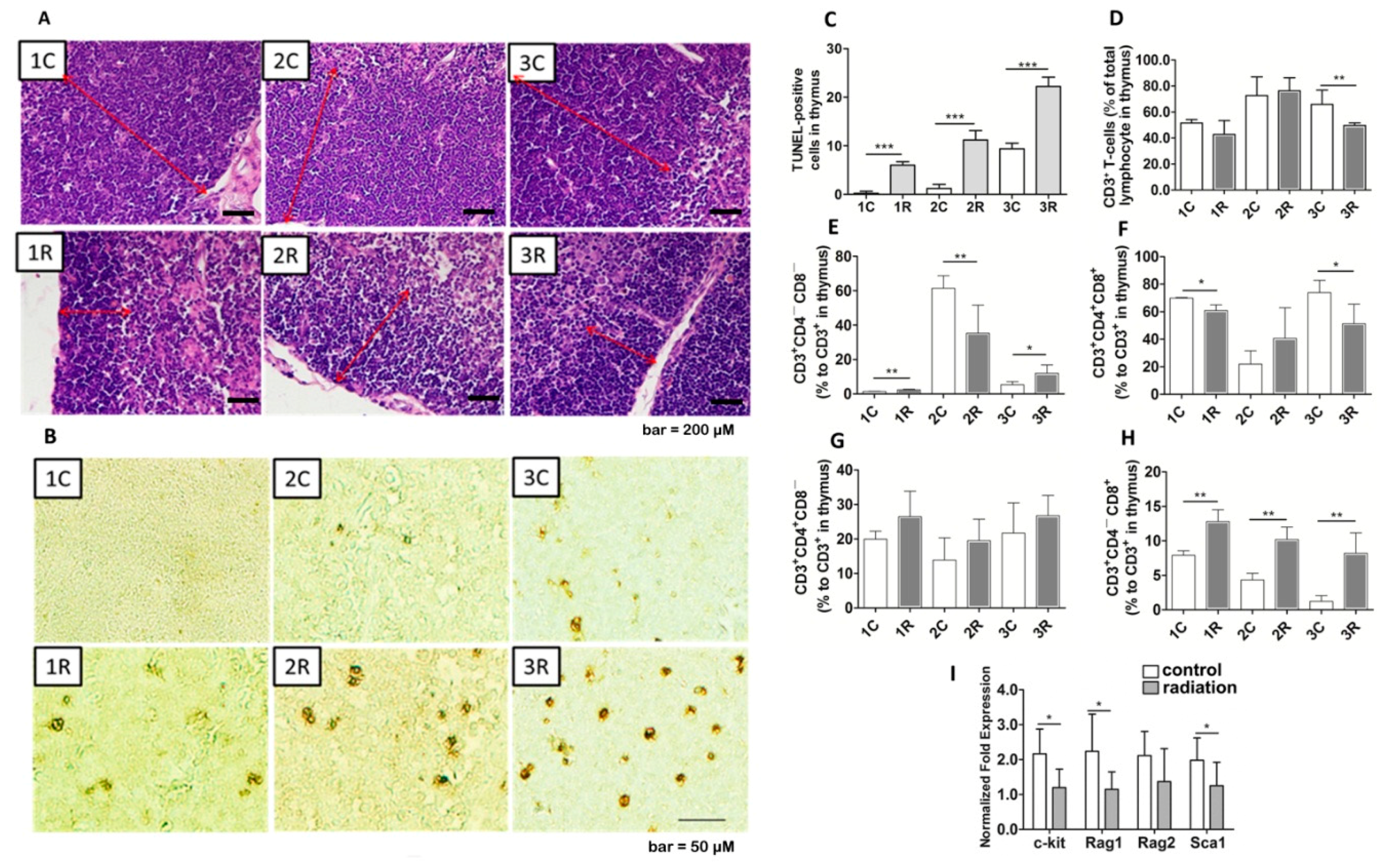
2.3. BLCIR-Induced Apoptosis, Abnormality of T-Cell Development in Thymus

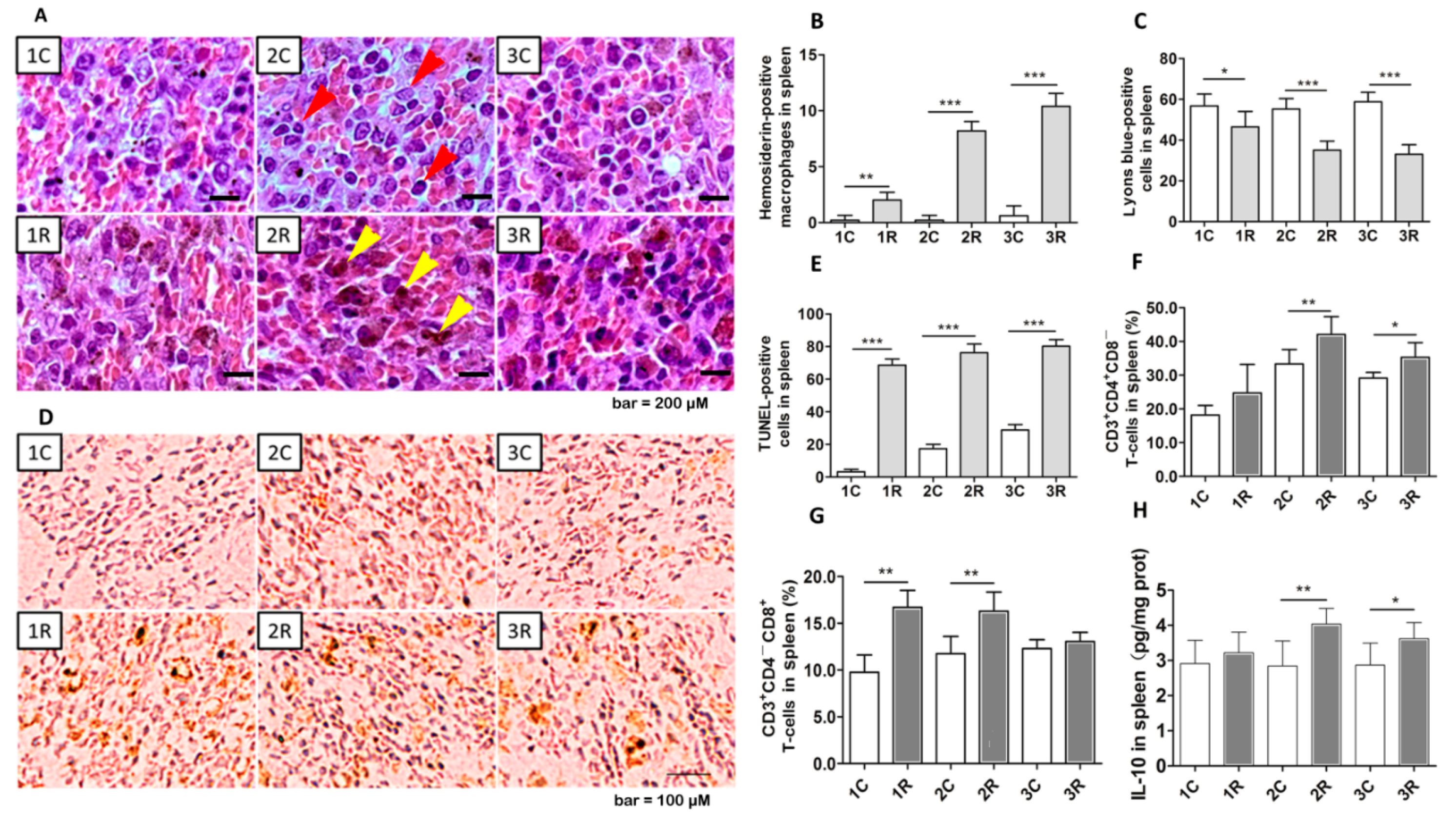
2.4. BLCIR Induced Apoptosis, T-Cell Distribution in Spleen

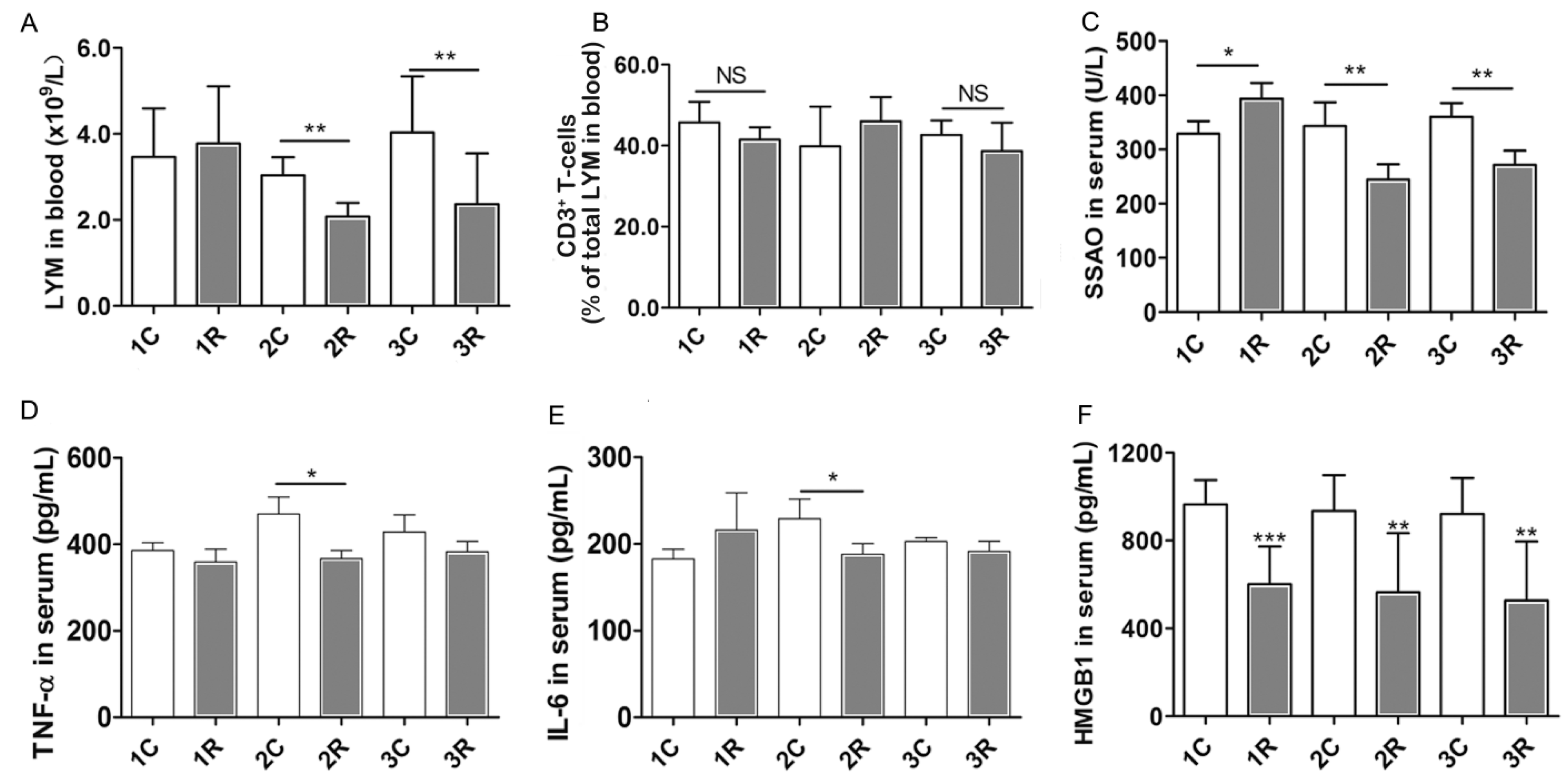
2.5. BLCIR-Induced Immunosuppression Features in Peripheral Blood


3. Discussion
4. Experimental Section
4.1. Cells and Animals
4.2. Carbon Ion Radiation Procedure and Experimental Design
4.3. Histopathology and Apoptosis
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Real-Time Quantitative PCR
4.6. Flow Cytometry Analysis of T Lymphocyte Subsets
4.7. Complete Blood Count of Peripheral Blood
4.8. Cell Viability Assay
4.9. Cell Migration Assays
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BLCIR | brain-localized carbon ion radiation |
| SSAO | semicarbazide-sensitive amino oxidase |
| TNF-α | tumor necrosis factor-α |
| IL-6 | interleukin 6 |
| HMGB1 | high mobility group box-1 |
| NS | not significant |
| LYM | lymphocyte |
| TUNEL | terminal deoxynucleotidyltransferase dUTP nick end labeling |
| PCR | polymerase chain reaction |
| IL-10 | interleukin 10 |
| HPA | hypothalamic-pituitary-adrenal |
| GR | glucocorticoid receptor |
| GC | glucocorticoid |
| FBS | fetal bovine serum |
| HIRFL | Heavy Ion Research Facility in Lanzhou |
| LET | liner energy transfer |
| ELISA | enzyme-linked immunosorbent assay |
References
- Miyawaki, D.; Murakami, M.; Demizu, Y.; Sasaki, R.; Niwa, Y.; Terashima, K.; Nishimura, H.; Hishikawa, Y.; Sugimura, K. Brain injury after proton therapy or carbon ion therapy for head-and-neck cancer and skull base tumors. Int. J. Radiat. Oncol. Biol. Phys. 2009, 75, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Combs, S.E.; Kessel, K.; Habermehl, D.; Haberer, T.; Jakel, O.; Debus, J. Proton and carbon ion radiotherapy for primary brain tumors and tumors of the skull base. Acta Oncol. 2013, 52, 1504–1509. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T. Particle radiotherapy with carbon ion beams. EPMA J. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Suit, H.; DeLaney, T.; Goldberg, S.; Paganetti, H.; Clasie, B.; Gerweck, L.; Niemierko, A.; Hall, E.; Flanz, J.; Hallman, J.; et al. Proton vs. carbon ion beams in the definitive radiation treatment of cancer patients. Radiother. Oncol. 2010, 95, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Padovani, L.; Andre, N.; Constine, L.S.; Muracciole, X. Neurocognitive function after radiotherapy for paediatric brain tumours. Nat. Rev. Neurol. 2012, 8, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Reppingen, N.; Held, K.D. Immunologically augmented cancer treatment using modern radiotherapy. Trends Mol. Med. 2013, 19, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, R.A.; Murillo-Rojas, J. Alzheimer’s disease and HLA-A2: Linking neurodegenerative to immune processes through an in silico approach. Biomed. Res. Int. 2014, 2014, 791238. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Kummer, M.P.; Latz, E. Innate immune activation in neurodegenerative disease. Nat. Rev. Immunol. 2014, 14, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Maguire-Zeiss, K.A.; Federoff, H.J. Future directions for immune modulation in neurodegenerative disorders: Focus on Parkinson’s disease. J. Neural Transm. 2010, 117, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, A.A.; Ingrassia, A.; de Menezes, R.X.; van Kesteren, R.E.; Rozemuller, A.J.; Heutink, P.; van de Berg, W.D. Evidence for Immune Response, Axonal Dysfunction and Reduced Endocytosis in the Substantia Nigra in Early Stage Parkinson’s Disease. PLoS ONE 2015, 10, e0128651. [Google Scholar] [CrossRef] [PubMed]
- Trager, U.; Andre, R.; Lahiri, N.; Magnusson-Lind, A.; Weiss, A.; Grueninger, S.; McKinnon, C.; Sirinathsinghji, E.; Kahlon, S.; Pfister, E.L.; et al. HTT-lowering reverses Huntington’s disease immune dysfunction caused by NFkappaB pathway dysregulation. Brain 2014, 137, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Zhou, H.; Chai, Y.; Ponnaiya, B.; Ivanov, V.N. Radiation induced non-targeted response: Mechanism and potential clinical implications. Curr. Mol. Pharmacol. 2011, 4, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Siva, S.; MacManus, M.P.; Martin, R.F.; Martin, O.A. Abscopal effects of radiation therapy: A clinical review for the radiobiologist. Cancer Lett. 2015, 356, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Formenti, S.C.; Demaria, S. Systemic effects of local radiotherapy. Lancet Oncol. 2009, 10, 718–726. [Google Scholar] [CrossRef]
- Mella, C.; Suarez-Arrabal, M.C.; Lopez, S.; Stephens, J.; Fernandez, S.; Hall, M.W.; Ramilo, O.; Mejias, A. Innate immune dysfunction is associated with enhanced disease severity in infants with severe respiratory syncytial virus bronchiolitis. J. Infect. Dis. 2013, 207, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Gersemann, M.; Wehkamp, J.; Stange, E.F. Innate immune dysfunction in inflammatory bowel disease. J. Intern. Med. 2012, 271, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Apetoh, L.; Ghiringhelli, F.; Tesniere, A.; Obeid, M.; Ortiz, C.; Criollo, A.; Mignot, G.; Maiuri, M.C.; Ullrich, E.; Saulnier, P.; et al. Toll-like receptor 4-dependent contribution of the immune system to anticancer chemotherapy and radiotherapy. Nat. Med. 2007, 13, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Burnette, B.C.; Liang, H.; Lee, Y.; Chlewicki, L.; Khodarev, N.N.; Weichselbaum, R.R.; Fu, Y.X.; Auh, S.L. The efficacy of radiotherapy relies upon induction of type i interferon-dependent innate and adaptive immunity. Cancer Res. 2011, 71, 2488–2496. [Google Scholar] [CrossRef] [PubMed]
- Pecaut, M.J.; Gridley, D.S. Impact of head-only iron ion radiation on the peripheral LPS response. Vivo 2011, 25, 903–916. [Google Scholar]
- Bentzen, S.M. Preventing or reducing late side effects of radiation therapy: Radiobiology meets molecular pathology. Nat. Rev. Cancer 2006, 6, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Harrabi, S.B.; Adeberg, S.; Welzel, T.; Rieken, S.; Habermehl, D.; Debus, J.; Combs, S.E. Long term results after fractionated stereotactic radiotherapy (FSRT) in patients with craniopharyngioma: Maximal tumor control with minimal side effects. Radiat. Oncol. 2014, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Meral, R.; Tambas, M.; Guveli, M.E.; Kemikler, G.; Altun, M.; Cakir, A.; Okutan, M.; Ahmed, S.M.; Donmez, N.; Disci, R. Side Effects of Stereotactic Fractionated Radiation Therapy for Malignant and Benign Lesions. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, S831. [Google Scholar] [CrossRef]
- Moretones, C.; Villa, S.; Navarro, V.; Villabona, C.; Majos, C.; Acebes, J.; Soler, J. Pituitary adenomas: Two different methods of fractionated stereotactic radiotherapy. Outcome and side effects. Radiother. Oncol. 2004, 73, S431. [Google Scholar]
- Wang, J.; Boerma, M.; Kulkarni, A.; Hollenberg, M.D.; Hauer-Jensen, M. Activation of protease activated receptor 2 by exogenous agonist exacerbates early radiation injury in rat intestine. Int. J. Radiat. Oncol. Biol. Phys. 2010, 77, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Geraci, J.P.; Mariano, M.S.; Jackson, K.L. Radiation hepatology of the rat: Time-dependent recovery. Radiat. Res. 1993, 136, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Bala, M. Concerted action of Nrf2-ARE pathway, MRN complex, HMGB1 and inflammatory cytokines—Implication in modification of radiation damage. Redox Biol. 2014, 2, 832–846. [Google Scholar]
- Carver, J.R.; Shapiro, C.L.; Ng, A.; Jacobs, L.; Schwartz, C.; Virgo, K.S.; Hagerty, K.L.; Somerfield, M.R.; Vaughn, D.J. American Society of Clinical Oncology clinical evidence review on the ongoing care of adult cancer survivors: Cardiac and pulmonary late effects. J. Clin. Oncol. 2007, 25, 3991–4008. [Google Scholar] [CrossRef] [PubMed]
- Stone, H.B.; Coleman, C.N.; Anscher, M.S.; McBride, W.H. Effects of radiation on normal tissue: Consequences and mechanisms. Lancet Oncol. 2003, 4, 529–536. [Google Scholar] [CrossRef]
- Wang, F.; Awan, U.F.; Wang, Y.; Wang, L.; Qing, H.; Ma, H.; Deng, Y. Damage of neuroblastoma cell SH-SY5Y mediated by MPP+ inhibits proliferation of T-cell leukemia Jurkat by co-culture system. Int. J. Mol. Sci. 2014, 15, 10738–10750. [Google Scholar] [CrossRef] [PubMed]
- Dooley, J.; Liston, A. Molecular control over thymic involution: From cytokines and microRNA to aging and adipose tissue. Eur. J. Immunol. 2012, 42, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Massa, S.; Balciunaite, G.; Ceredig, R.; Rolink, A.G. Critical role for c-kit (CD117) in T cell lineage commitment and early thymocyte development in vitro. Eur. J. Immunol. 2006, 36, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Bassing, C.H.; Swat, W.; Alt, F.W. The mechanism and regulation of chromosomal V(D)J recombination. Cell 2002, 109, S45–S55. [Google Scholar] [CrossRef]
- Spangrude, G.J.; Aihara, Y.; Weissman, I.L.; Klein, J. The Stem-Cell Antigens Sca-1 and Sca-2 Subdivide Thymic and Peripheral Lymphocyte-T into Unique Subsets. J. Immunol. 1988, 141, 3697–3707. [Google Scholar] [PubMed]
- Sologuren, I.; Rodríguez-Gallego, C.; Lara, P.C. Immune effects of high dose radiation treatment: Implications of ionizing radiation on the development of bystander and abscopal effects. Transl. Cancer Res. 2014, 3, 18–31. [Google Scholar]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Maxeiner, H.G.; Marion Schneider, E.; Kurfiss, S.T.; Brettschneider, J.; Tumani, H.; Bechter, K. Cerebrospinal fluid and serum cytokine profiling to detect immune control of infectious and inflammatory neurological and psychiatric diseases. Cytokine 2014, 69, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Barney, M.; Call, G.B.; McIlmoil, C.J.; Husein, O.F.; Adams, A.; Balls, A.G.; Oliveira, G.K.; Miner, E.C.; Richards, R.A.; Crawford, B.K.; et al. Stimulation by interleukin-6 and inhibition by tumor necrosis factor of cortisol release from bovine adrenal zona fasciculata cells through their receptors. Endocrine 2000, 13, 369–377. [Google Scholar] [CrossRef]
- Talaber, G.; Tuckermann, J.P.; Okret, S. ACTH controls thymocyte homeostasis independent of glucocorticoids. Faseb J. 2015, 29, 2526–2534. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Lublin, F.D.; Knobler, R.L. Modulation of T cell-endothelial adhesion by astrocyte conditioned medium. Glia 1997, 21, 408–412. [Google Scholar] [CrossRef]
- Zhang, H.X.; Shao, B.; Zhuge, Q.C.; Wang, P.; Zheng, C.C.; Huang, W.L.; Yang, C.Q.; Wang, B.; Su, D.M.; Jin, K.L. Cross-Talk between Human Neural Stem/Progenitor Cells and Peripheral Blood Mononuclear Cells in an Allogeneic Co-Culture Model. PLoS ONE 2015, 10, e0117432. [Google Scholar] [CrossRef] [PubMed]
- Derventzi, A.; Nikolopoulou, M.; Apostolou, A.; Kataki, A.; Bakopoulos, K.; Androulis, A.; Kilidireas, C.; Zografos, G.; Konstadoulakis, M.M. An in vitro model for investigating human autologous neuronal-astrocyte and immune cell interactions underlying neurodegenerative and immunosuppressive processes in neuropathy. Brain Res. 2014, 1587, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.E.; Li, Z.X.; Liu, Y.; Moulder, J.E.; Zhao, M. Whole-Body Imaging of High-Dose Ionizing Irradiation-Induced Tissue Injuries Using Tc-99m-Duramycin. J. Nucl. Med. 2013, 54, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.B.; Chen, F.; Yao, X.C.; Zhu, J.B.; Wang, C.; Zhang, J.L.; Li, X.Y. Protective Effect of Lycium ruthenicum Murr. Against Radiation Injury in Mice. Int. J. Environ. Res. Public Health 2015, 12, 8332–8347. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Zhen-Yu, W.; Xin, Y.; Lin, Y.; Li-Li, Z.; Hai-Na, B.; Xiao-Yu, L. Protective effects of sulfated derivatives of polysaccharides extracted from Auricularia auricular on hematologic injury induced by radiation. Int. J. Radiat. Res. 2014, 12, 99–111. [Google Scholar]
- Park, H.S.; Kim, S.; Lee, Y.; Choi, M.S.; Choi, M.U. Alteration of lipid composition of rat thymus during thymic atrophy by whole-body X-irradiation. Int. J. Radiat. Biol. 2006, 82, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Qiao, S.J.; Tuckermann, J.; Okret, S.; Jondal, M. Thymus-derived glucocorticoids mediate androgen effects on thymocyte homeostasis. Faseb J. 2010, 24, 5043–5051. [Google Scholar] [CrossRef] [PubMed]
- Purton, J.F.; Zhan, Y.F.; Liddicoat, D.R.; Hardy, C.L.; Lew, A.M.; Cole, T.J.; Godfrey, D.I. Glucocorticoid receptor deficient thymic and peripheral T cells develop normally in adult mice. Eur. J. Immunol. 2002, 32, 3546–3555. [Google Scholar] [CrossRef]
- Radu, C.G.; Cheng, D.H.; Nijagal, A.; Riedinger, M.; McLaughlin, J.; Yang, L.V.; Johnson, J.; Witte, O.N. Normal immune development and glucocorticoid-induced thymocyte apoptosis in mice deficient for the T-Cell death-associated gene 8 receptor. Mol. Cell. Biol. 2006, 26, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Zilberman, Y.; Zafrir, E.; Ovadia, H.; Yefenof, E.; Guy, R.; Sionov, R.V. The glucocorticoid receptor mediates the thymic epithelial cell-induced apoptosis of CD4+8+ thymic lymphoma cells. Cell. Immunol. 2004, 227, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Pozzesi, N.; Fierabracci, A.; Liberati, A.M.; Martelli, M.P.; Ayroldi, E.; Riccardi, C.; Delfino, D.V. Role of caspase-8 in thymus function. Cell Death Differ. 2014, 21, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P. The Laboratory Rat: Relating Its Age With Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Petrovic, J.S.; Markovic, R.Z. Changes in Cortisol Binding to Soluble Receptor Proteins in Rat-Liver and Thymus during Development and Aging. Dev. Biol. 1975, 45, 176–182. [Google Scholar] [CrossRef]
- Straub, R.H.; Rauch, L.; Fassold, A.; Lowin, T.; Pongratz, G. Neuronally Released Sympathetic Neurotransmitters Stimulate Splenic Interferon-gamma Secretion From T Cells in Early Type II Collagen-Induced Arthritis. Arthritis Rheum. 2008, 58, 3450–3460. [Google Scholar] [CrossRef] [PubMed]
- Meinel, T.; Pongratz, G.; Rauch, L.; Straub, R.H. Neuronal alpha 1/2-adrenergic stimulation of IFN-gamma, IL-6, and CXCL-1 in murine spleen in late experimental arthritis. Brain Behav. Immun. 2013, 33, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H. Complexity of the bi-directional neuroimmune junction in the spleen. Trends Pharmacol. Sci. 2004, 25, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Harris, T.J.; Lim, M.; Drake, C.G.; Tran, P.T. Immune modulation and stereotactic radiation: Improving local and abscopal responses. Biomed. Res. Int. 2013, 2013, 658126. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Rao, L.; Wang, H.L.; Mao, Z.W.; Lei, R.H.; Yang, Z.Y.; Qing, H.; Deng, Y.L. Transcriptome analysis of glioma cells for the dynamic response to gamma-irradiation and dual regulation of apoptosis genes: A new insight into radiotherapy for glioblastomas. Cell Death Dis. 2013, 4, e895. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, R.; Zhao, T.; Li, Q.; Wang, X.; Ma, H.; Deng, Y. Carbon Ion Irradiated Neural Injury Induced the Peripheral Immune Effects in Vitro or in Vivo. Int. J. Mol. Sci. 2015, 16, 28334-28346. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226109
Lei R, Zhao T, Li Q, Wang X, Ma H, Deng Y. Carbon Ion Irradiated Neural Injury Induced the Peripheral Immune Effects in Vitro or in Vivo. International Journal of Molecular Sciences. 2015; 16(12):28334-28346. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226109
Chicago/Turabian StyleLei, Runhong, Tuo Zhao, Qiang Li, Xiao Wang, Hong Ma, and Yulin Deng. 2015. "Carbon Ion Irradiated Neural Injury Induced the Peripheral Immune Effects in Vitro or in Vivo" International Journal of Molecular Sciences 16, no. 12: 28334-28346. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226109