
Vaccination of Silver Sea Bream (Sparus sarba) against Vibrio alginolyticus: Protective Evaluation of Different Vaccinating Modalities

Abstract
:1. Introduction
2. Results
2.1. Safety Assessment
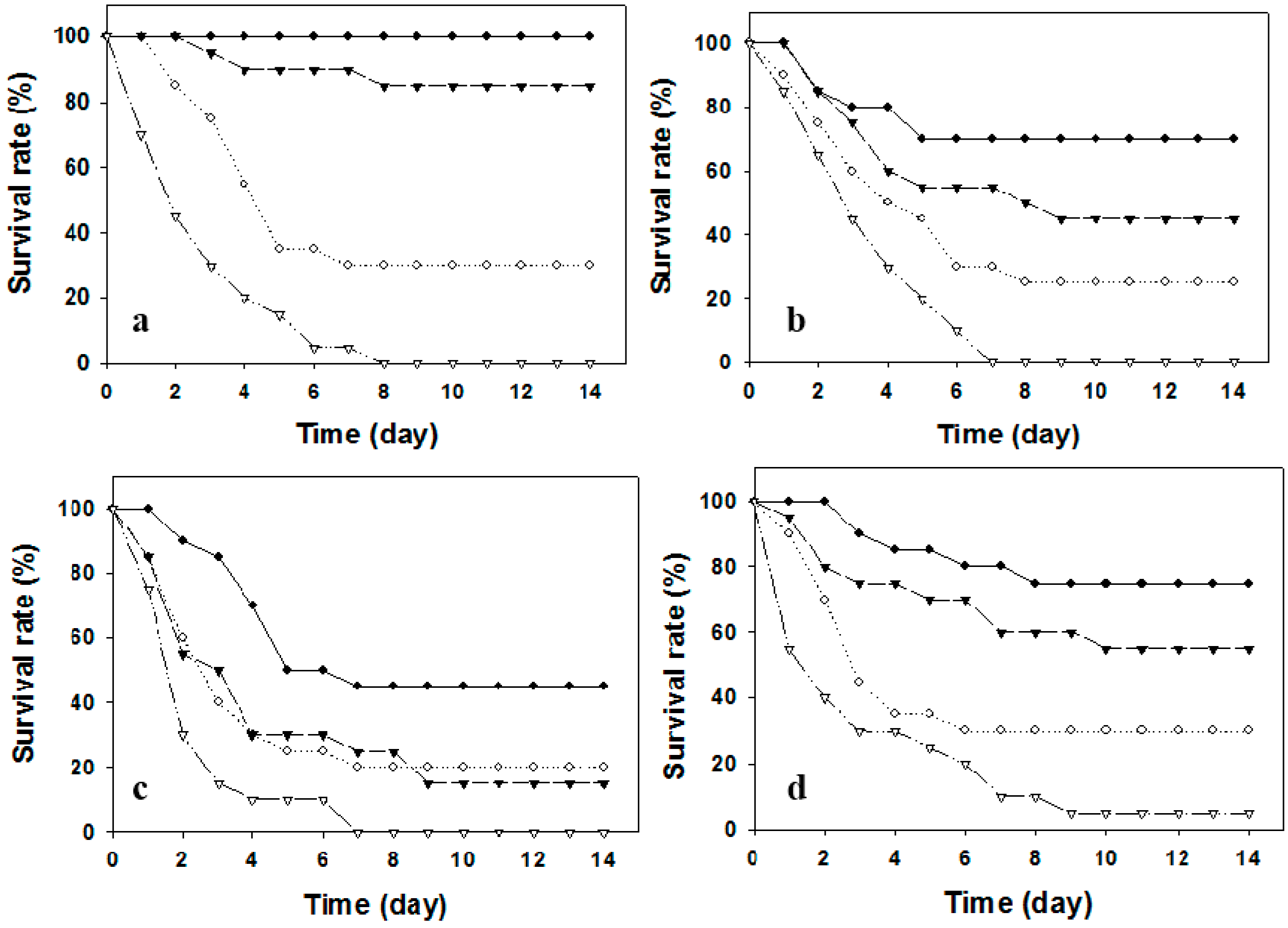
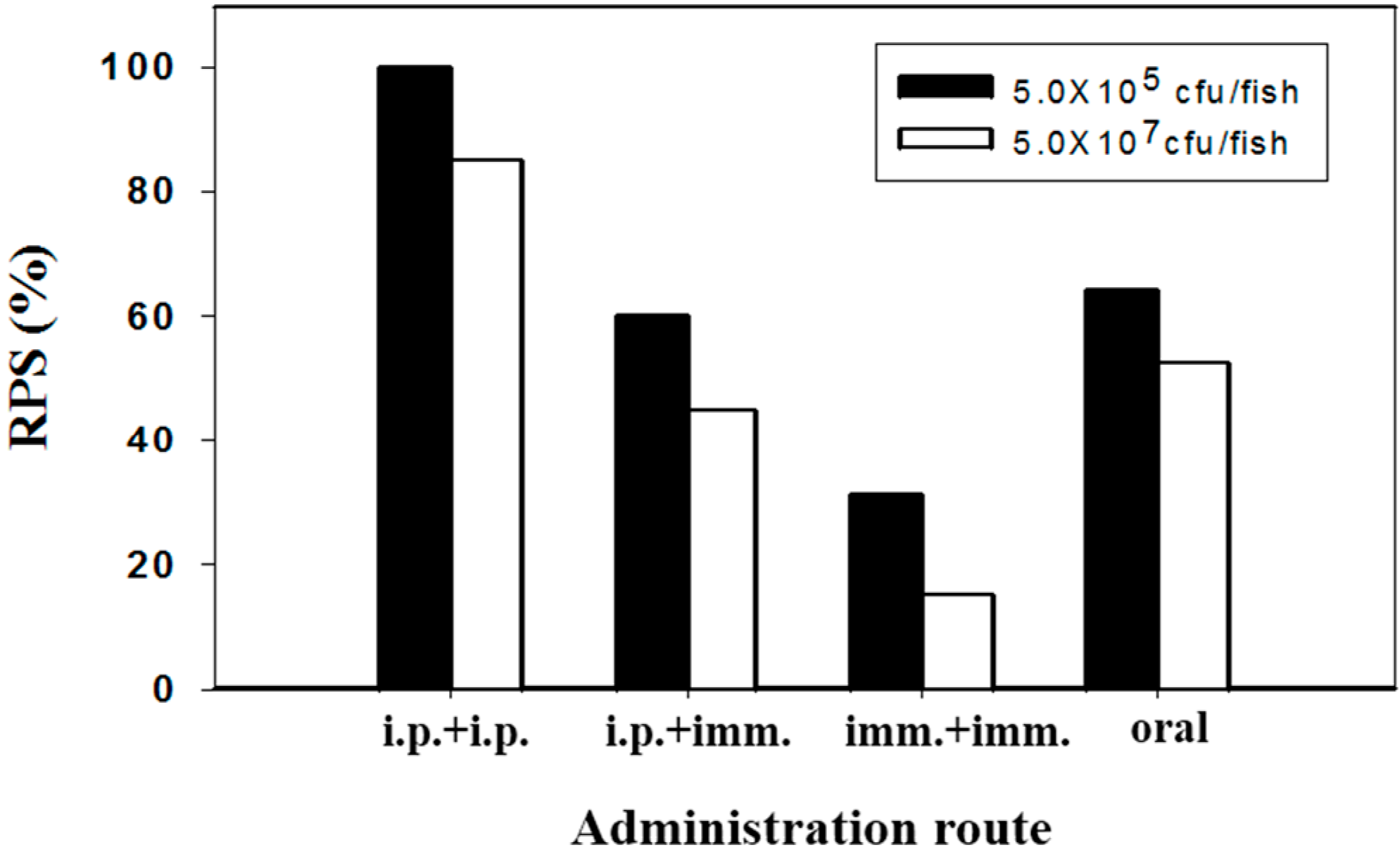
2.2. Protection against Pathogenic Challenge

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccines | Number of Vaccinated Fish | Number of Challenged Fish | Number of Dead Fish | Mortality (%) | RPS |
|---|---|---|---|---|---|
| Whole Cell Bacterins | |||||
| Formalin-killed | 36 | 35 | 7 | 20 | 80 |
| Phenol-killed | 36 | 34 | 8 | 23.5 | 76.5 |
| Chloroform-killed | 36 | 35 | 25 | 71.4 | 28.6 |
| Heat-killed | 36 | 36 | 18 | 50 | 50 |
| Subcellular Bacterins | |||||
| LPS | 36 | 36 | 0 | 0 | 100 |
| Control | 36 | 36 | 36 | 100 | 0 |


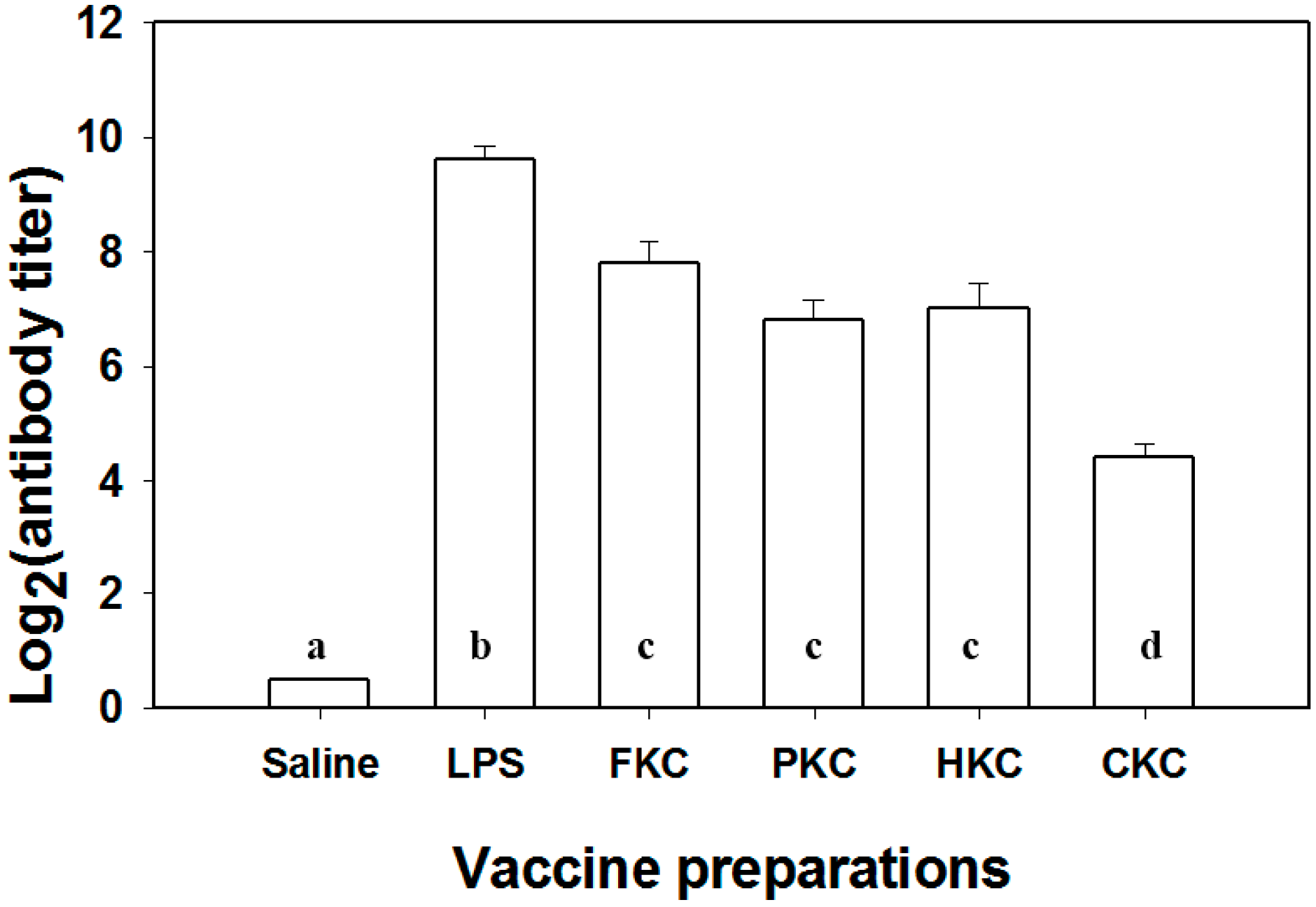
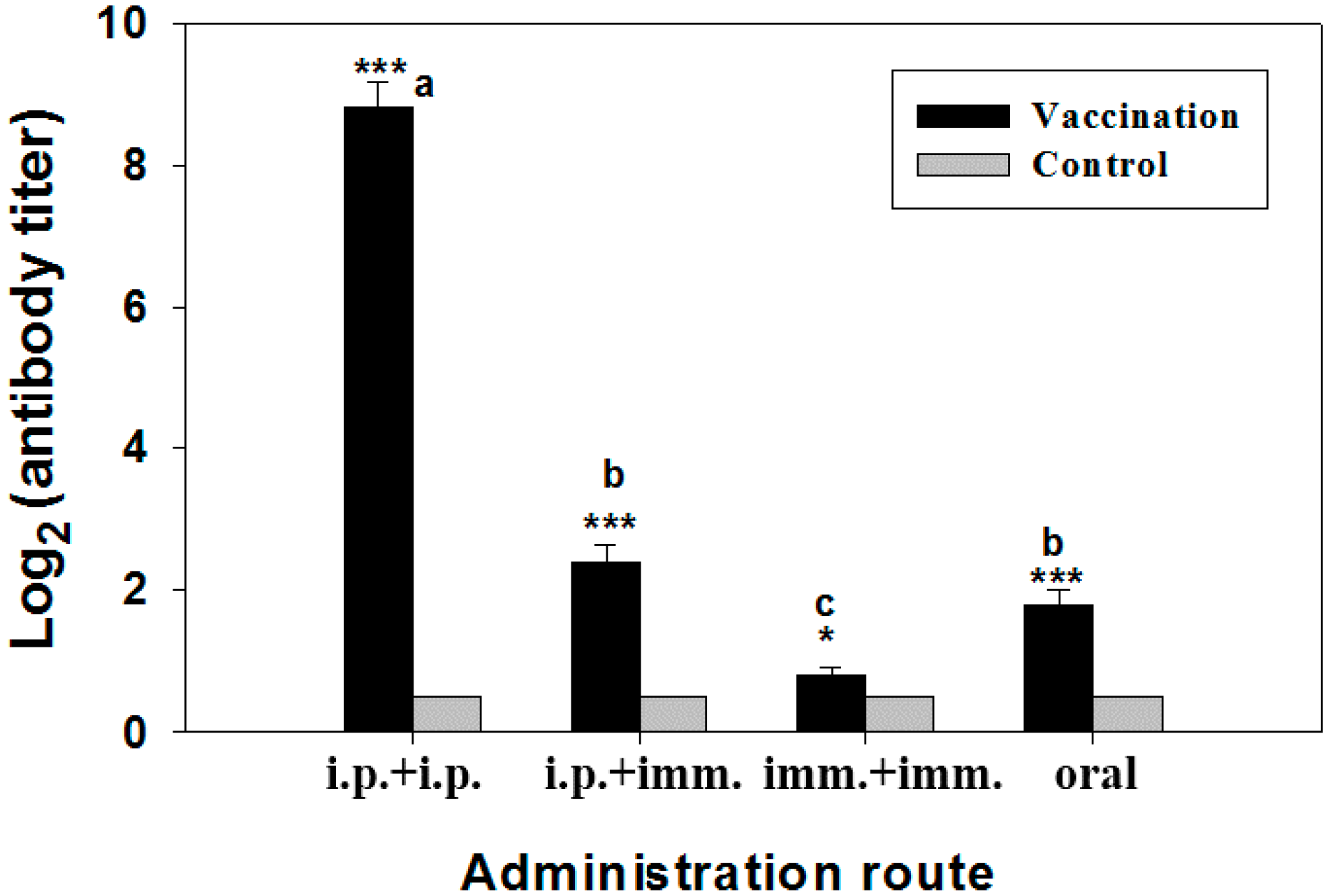
2.3. Specific Antibody Titer


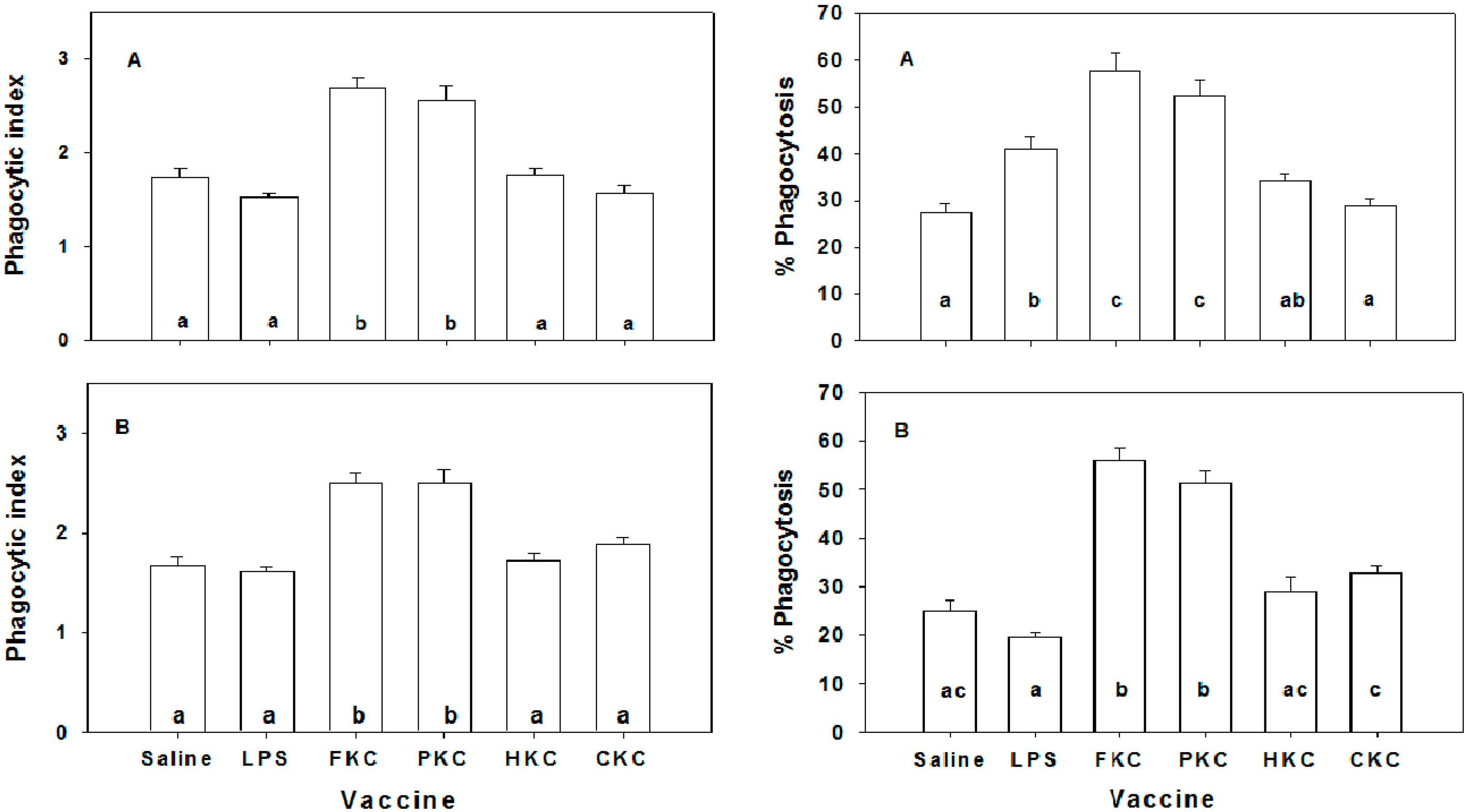
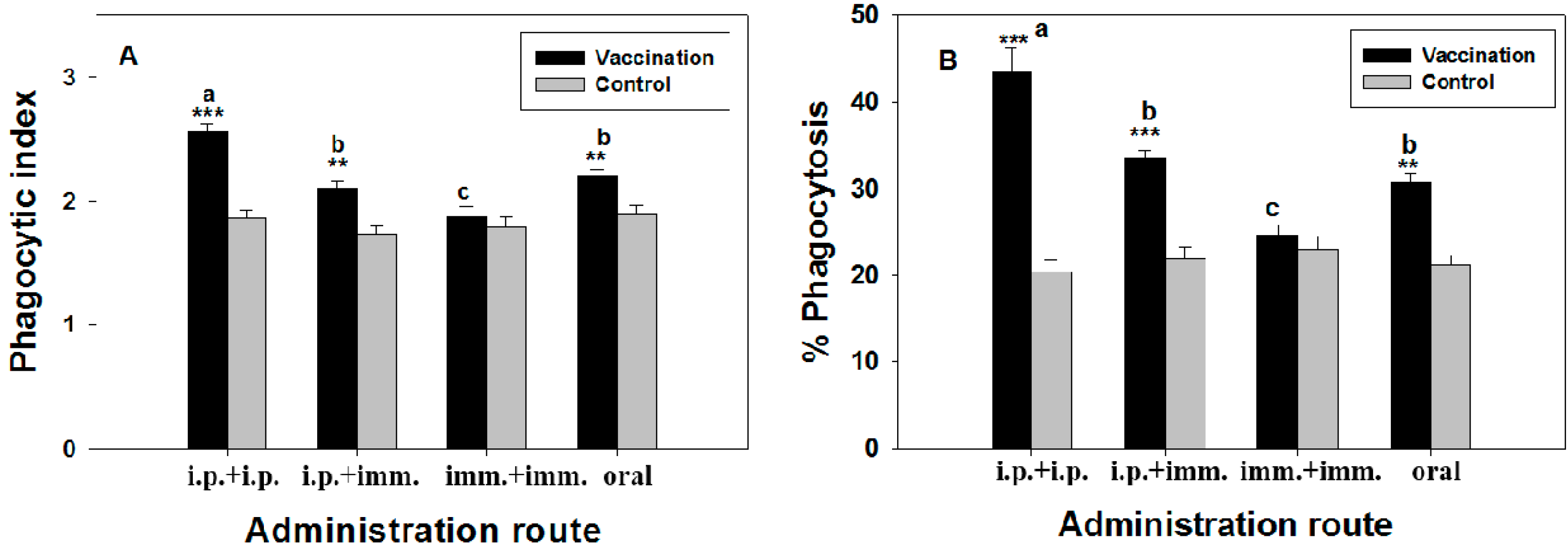
2.4. Macrophage Phagoctic Activity


2.5. Hematology and Organ Weight

3. Discussion
4. Materials and Methods
4.1. Experimental Fish
4.2. Bacterial Cultures
4.3. Vaccine Preparation
4.4. Immunization Strategies
4.5. Pathogenic Challenge
4.6. Hematological Parameters and Organ Weights
4.7. Serum Cortisol
4.8. Agglutinating Antibody Titer
4.9. Macrophage Phagocytosis
4.10. Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens, 2nd ed.; Austin, B., Austin, D.A., Eds.; Ellis Horwood, Ltd.: Chichester, UK, 1993; pp. 265–307. [Google Scholar]
- Toranzo, A.E.; Magarinos, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture system. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Colquhoun, A.J.; Lillehaug, A. Vaccination against Vibriosis. In Fish Vaccination, 1st ed.; Gudding, R., Lillehaug, A., Evensen, O., Eds.; John Wiley & Sons, Ltd.: San Francisco, CA, USA, 2014; pp. 172–184. [Google Scholar]
- Lillehaug, A. Vaccination Strategies and Procedures. In Fish Vaccination, 1st ed.; Gudding, R., Lillehaug, A., Evensen, O., Eds.; John Wiley & Sons, Ltd.: San Francisco, CA, USA, 2014; pp. 140–152. [Google Scholar]
- Dec, C.; Angelidis, P.; Laurencin, F.B. Effects of oral vaccination against vibriosis in turbot, Scophthalmus maximus (L.), and sea bass, Dicentrarchus labrax (L.). J. Fish Dis. 1990, 13, 369–376. [Google Scholar] [CrossRef]
- Ainsworth, A.J.; Rice, C.D.; Xue, L. Immune responses of channel catfish, Ictalurus punctatus (Rafinsque) after oral or intraperitoneal vaccination with particulate or soluble Edwardsiella ictaluri antigen. J. Fish Dis. 1995, 18, 397–409. [Google Scholar] [CrossRef]
- Ji, R.; Zou, W.; Hu, S.; Yan, Q. Vaccination in three different ways against vibriosis of Seriola dumerili caused by Vibrio hollisae. Chin. J. Oceanol. Limnol. 2008, 26, 233–237. [Google Scholar] [CrossRef]
- Hu, Y.H.; Deng, T.; Sun, B.G.; Sun, L. Development and efficacy of an attenuated Vibrio harveyi vaccine candidate with cross protectivity against Vibrio alginolyticus. Fish Shellfish Immunol. 2012, 32, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Galeotti, M.; Romano, N.; Volpatti, D.; Bulfon, C.; Brunetti, A.; Tiscar, P.G.; Mosca, F.; Bertonii, F.; Marchetti, M.G.; Abelli, L. Innovative vaccination protocol against vibriosis in Dicentrarchus labrax (L.) junveniles: Improvement of immune parameters and protection to challenge. Vaccine 2013, 31, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Santarem, M.M.; Figueras, A. Kinetics of phagocytic activity, plaque-forming cells and specific agglutinins of turbot (Scophthalmus maximus L.) immunized with O-antigen of Vibrio damsela and Pasteurella piscicida. Fish Shellfish Immunol. 1994, 4, 527–537. [Google Scholar] [CrossRef]
- Schroder, M.B.; Mikkelsen, H.; Bordal, S.; Gravningen, K.; Lund, V. Early vaccination and protection of Atlantic cod (Gadus morhua L.) juveniles against classical vibriosis. Aquaculture 2006, 254, 46–53. [Google Scholar] [CrossRef]
- Mikkelsen, H.; Lund, V.; Larsen, R.; Seppola, M. Vibriosis vaccines based on various sero-subgroups of Vibio anguillarum O2 induce specific protection in Atlantic cod (Gadus morhua L.). Fish Shellfish Immunol. 2011, 3, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wu, H.; Xiao, J.; Wang, Q.; Liu, Q.; Zhang, Y. Immune response evoked by infection with Vibrio anguillarum in zebrafish bath–vaccinated with a live attenuated strain. Veterin. Immunol. Immunopathol. 2013, 154, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, B.; Wu, H.; Gao, L.; Liu, Q.; Wang, Q.; Xiao, J.; Zhang, Y. Th17-like immune response in fish mucosal tissues after administration of live attenuated Vibrio anguillarum via different vaccination routes. Fish Shellfish Immunol. 2014, 37, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Joosten, P.M.H.; Kruijer, W.J.; Rambout, J.H.W.M. Anal immunization of carp and rainbow trout with different fractions of a Vibrio anguillarum bacterin. Fish Shellfish Immunol. 1996, 6, 541–551. [Google Scholar] [CrossRef]
- Joosten, P.H.M.; Tiemersma, E.; Threels, A.; Caumartin-Dhieux, C.; Rombout, J.H.W.M. Oral vaccination of fish against Vibrio anguillarum using alginate microparticles. Fish Shellfish Immunol. 1997, 7, 471–485. [Google Scholar] [CrossRef]
- Estevez, J.; Leiro, J.; Toranzo, A.E.; Barja, J.L.; Ubeira, F.M. Kinetics of antibody production against Vibrio anguillarum antigens in turbot. Aquaculture 1994, 123, 191–196. [Google Scholar] [CrossRef]
- Estevez, J.; Leiro, J.; Toranzo, A.E.; Barja, J.L.; Ubeira, F.M. Role of serum antibodies in protection of vaccinated turbot (Scophthalmus maximus) against vibriosis. Aquaculture 1994, 123, 197–204. [Google Scholar] [CrossRef]
- Stromsheim, A.; Eide, D.M.; Hofgaard, P.O.; Larsen, H.J.S.; Refstie, T.; Roed, K.H. Genetic variation in the humoral immune response against Vibrio salmonicida and in antibody titer against Vibrio anguillarum and total IgM in Atlantic salmon (Salmo salar). Veterin. Immunol. Immunopathol. 1994, 44, 85–95. [Google Scholar] [CrossRef]
- Gudmundsdottir, S.; Magnadottir, B.; Bjornsdottir, B.; Arnadottir, H.; Gudmundsdottir, B.K. Specific and natural antibody response of cod juveniles vaacinated against Vibrio anguillarum. Fish Shellfish Immunol. 2009, 26, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Santarem, M.M.; Figueras, A. Leukocyte numbers and phagocytic activity in turbot Scophthalmus maximus L. following immunization with Vibrio damsela and Pasteurella piscicida O-antigen bacterins. Dis. Aquat. Organ. 1995, 23, 213–220. [Google Scholar] [CrossRef]
- Solem, S.T.; Jorgensen, J.B.; Roberson, B. Stimulation of respiratory burst and phagocytic activity in Atlantic salmon (Salmo salar L.) macrophages by lipopolysaccharide. Fish Shellfish Immunol. 1995, 5, 475–491. [Google Scholar] [CrossRef]
- Magarinos, B.; Romalde, J.L.; Santos, Y.; Casal, J.F.; Barja, J.L.; Toranzo, A.E. Vaccination trials on gilthead sea bream (Sparus aurata) against Pasteurella iscicida. Aquaculture 1994, 120, 201–208. [Google Scholar] [CrossRef]
- Li, J.; Feng, J.; Liu, X.; Li, Q.F.; Woo, N.Y.S.; Xu, H.S. Vibrio alginolyticus, a pathogen of silver sea bream, Sparus sarba, cultured in Hong Kong. J. Fish. Chin. 1998, 22, 275–278. [Google Scholar]
- Li, J.; Zhou, L.R.; Woo, N.Y.S. Invasion route and pathogenic mechanisms of Vibrio alginolyticus to silver sea bream (Sparus sarba). J. Aquat. Anim. Health 2003, 15, 302–313. [Google Scholar] [CrossRef]
- Li, J.; Yie, J.; Foo, R.W.T.; Ling, J.M.L.; Xu, H.; Woo, N.Y.S. Antibiotic resistance and plasmid profiles of vibrio isolates from cultured silver sea bream, Sparus sarba. Mar. Pollut. Bull. 1999, 39, 245–249. [Google Scholar] [CrossRef]
- Plant, K.P.; LaPatra, S.E. Advances in fish vaccine delivery. Dev. Comp. Immunol. 2011, 35, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Gudding, R. Vaccination as a protective measure. In Fish Vaccination, 1st ed.; Gudding, R., Lillehaug, A., Evensen, O., Eds.; John Wiley & Sons, Ltd.: San Francisco, CA, USA, 2014; pp. 12–21. [Google Scholar]
- Gutierrez, M.A.; Miyazaki, T. Responses of Japanese eel to oral challenge with Edwardsiella. tarda after vaccination with formalin-killed cells or lipopolysaccharide of the bacterium. J. Aquat. Anim. Health 1994, 6, 110–117. [Google Scholar] [CrossRef]
- Dalmo, R.A.; Seljelid, R. The immunomodulatory effect of LPS, laminaran and sulphated laminaran β(1,3)-d-glucan on Atlantic salmon, Salmo salar L., macrophages in vitro. J. Fish Dis. 1995, 18, 175–185. [Google Scholar] [CrossRef]
- Dalmo, R.A.; Ingebrigtsen, K.; Bogwald, J. Non-specific defense mechanisms in fish, with particular reference to the reticuloendothelial system (RES). J. Fish Dis. 1997, 20, 241–273. [Google Scholar] [CrossRef]
- Angelidis, P.; Baudin-Laurencin, F.; Youinou, P. Stress in rainbow trout, Salmo gairdneri: Effects upon phagocyte chemiluminescence, circulating leukocytes and susceptibility to Aeromonas salmonicida. J. Fish Biol. 1987, 31 (Suppl. A), S113–S122. [Google Scholar] [CrossRef]
- Sakai, M.; Konishi, M.; Atsuta, S.; Kobayashi, M. The chemiluminescent response of leukocytes from anterior kidney of rainbow trout Oncorhynchus mykiss vaccinated with Vibrio anguillarum, Streptococcus sp. or Renibacterium Salmoninarum. Nippon Suisan Gakkaishi 1991, 57, 237–241. [Google Scholar] [CrossRef]
- Qin, Q.; Pan, J. Safety and efficacy of Vibrio vulnificus bacterins vaccinated by injection administration against vibriosis in Epinephelus awoara. Trop. Oceanol. 1996, 15, 7–12. [Google Scholar]
- Anderson, D.; Ross, A.J. Comparative study of Hagerman redmouth disease oral bacterins. Prog. Fish Cult. 1972, 34, 226–228. [Google Scholar] [CrossRef]
- Kakizawa, S.; Kaneko, T.; Hirano, T. Elevation of plasma somatolactin concentrations during acidosis in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 1996, 199, 1043–1051. [Google Scholar] [PubMed]
- Aakre, R.; Wergeland, H.I.; Aasjord, P.M.; Endresen, C. Enhanced antibody response in Atlantic salmon (Salmo salar L.) to Aeromonas salmonicida cell wall antigen using a bacteria containing β-1,3-m-glucan as adjuvant. Fish Shellfish Immunol. 1994, 4, 47–61. [Google Scholar] [CrossRef]
- Boesen, H.T.; Pedersen, K.; Koch, C.; Larsen, J.L. Immune response of rainbow trout (Oncorhynchus mykiss) to antigenic preparations from Vibrio anguillarum serogroup O1. Fish Shellfish Immunol. 1997, 7, 543–553. [Google Scholar] [CrossRef]
- Velji, M.I.; Albright, L.J.; Evelyn, T.P.T. Immunogenicity of various Vibrio ordalii lipopolysaccharide fractions in Coho salmon, Oncorhynchus kisutch. Dis. Aquat. Organ. 1992, 12, 97–101. [Google Scholar]
- Espelid, S.; Hjelmeland, K.; Jorgensen, T. The specificity of Atlantic salmon antibodies made against the fish pathogen Vibriosalmonicida, establishing the surface protein VS-P1 as the dominating antigen. Dev. Comp. Immunol. 1987, 11, 529–537. [Google Scholar] [CrossRef]
- Hjelmeland, K.; Stensvag, K.; Jorgensen, T.; Espelid, S. Isolation and characterization of a surface layer antigen from Vibrio salmonicida. J. Fish Dis. 1988, 10, 85–90. [Google Scholar] [CrossRef]
- Thune, R.L.; Collins, L.A.; Pena, M.P. A comparison of immersion, immersion/oral combination and injection methods for the vaccination of channel catfish Ictalurus punctatus against Edwardsiella ictaluri. J. World Aquac. Soc. 1997, 28, 193–201. [Google Scholar] [CrossRef]
- Hjeltnes, B.; Andersen, K.; Ellingsen, H.M. Vaccination against Vibrio salmonicida: the effect of different routes of administration and of revaccination. Aquaculture 1989, 83, 1–6. [Google Scholar] [CrossRef]
- Nordmo, R.; Sevatdal, S.; Ramstad, A. Experimental infection with Vibrio salmonicida in Atlantic salmon (Salmo salar L.): An evaluation of three different challenge methods. Aquaculture 1997, 158, 23–32. [Google Scholar] [CrossRef]
- Secombes, C.J. Enhancement of fish phagocyte activity. Fish Shellfish Immunol. 1994, 4, 421–436. [Google Scholar] [CrossRef]
- Deane, E.E.; Li, J.; Woo, N.Y.S. Hormonal status and phagocytic activity of sea bream infected with vibriosis. Comp. Biochem. Physiol. 2001, 129, 687–693. [Google Scholar] [CrossRef]
- Ellis, A.E. Recent development in oral vaccine delivery systems. Fish Pathol. 1995, 30, 293–300. [Google Scholar] [CrossRef]
- Wong, G.; Kaattari, S.L.; Christensen, J.M. Effectiveness of an oral enteric coated Vibrio vaccines for use in salmonid fish. Immunol. Investig. 1992, 21, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Bogwald, J.; Stensvag, K.; Hoffman, J.; Holm, K.O.; Jorgensen, T.Ø. Vaccination of Atlantic salmon, Salmon salar L., with particulate lipopolysaccharide antigens from Vibrio salmonicida and Vibrio anguillarum. Fish Shellfish Immunol. 1992, 2, 251–261. [Google Scholar] [CrossRef]
- Rombout, J.H.W.M.; Bot, H.E.; Taverne-Thiele, J.J. Immunological importance of the second gut segment of carp III. Systemic and/or mucosal immune responses after imunization with soluble or particulate antigen. J. Fish Biol. 1989, 25, 179–186. [Google Scholar] [CrossRef]
- Zhang, Y.A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Parra, D.; Gómez, D.; Salinas, I.; Zhang, Y.A.; von Gersdorff Jørgensen, L.; Heinecke, R.D.; Buchmann, K.; LaPatra, S.; Sunyer, J.O. Teleost skin, an ancient mucosal surface that elicits gut-like immune response. Proc. Natl. Acad. Sci. USA 2013, 110, 13097–13102. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.S.; Rohovec, J.S.; Fryer, J.L. Tissue location of Vibrio bacterin delivered by intraperitoneal injection, immersion and oral routes to Salmo gairdneri. Fish Pathol. 1985, 19, 263–269. [Google Scholar] [CrossRef]
- Woo, N.Y.S.; Kelly, S.P. Effects of salinity and nutritional status on growth and metabolism of Sparus sarba in a closed seawater system. Aquaculture 1995, 135, 229–238. [Google Scholar] [CrossRef]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides: Extraction with phenol-water and further applications of the procedure. In Methods in Carbohydrate Chemistry; Whistler, R.N., Bemiller, J.N., Wolffrom, M.L., Eds.; Academic Press: New York, NY, USA, 1965; pp. 83–91. [Google Scholar]
- Amend, D.F. Potency testing of fish vaccines. Developm. Biol. Standard. 1981, 49, 447–454. [Google Scholar]
- Roberson, B.S. Bacterial agglutination. In Techniques in Fish Immunology; Stolen, J.S., Flectcher, T.C., Anderson, D.P., Roberson, B.S., van Muiswinkel, W.B., Eds.; SOS Publications: Fair Haven, NJ, USA, 1990; pp. 81–86. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ma, S.; Woo, N.Y.S. Vaccination of Silver Sea Bream (Sparus sarba) against Vibrio alginolyticus: Protective Evaluation of Different Vaccinating Modalities. Int. J. Mol. Sci. 2016, 17, 40. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010040
Li J, Ma S, Woo NYS. Vaccination of Silver Sea Bream (Sparus sarba) against Vibrio alginolyticus: Protective Evaluation of Different Vaccinating Modalities. International Journal of Molecular Sciences. 2016; 17(1):40. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010040
Chicago/Turabian StyleLi, Jun, Siyuan Ma, and Norman Y. S. Woo. 2016. "Vaccination of Silver Sea Bream (Sparus sarba) against Vibrio alginolyticus: Protective Evaluation of Different Vaccinating Modalities" International Journal of Molecular Sciences 17, no. 1: 40. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010040