
Microarray Study of Pathway Analysis Expression Profile Associated with MicroRNA-29a with Regard to Murine Cholestatic Liver Injuries

 ,
,
Abstract
:
1. Introduction
2. Results
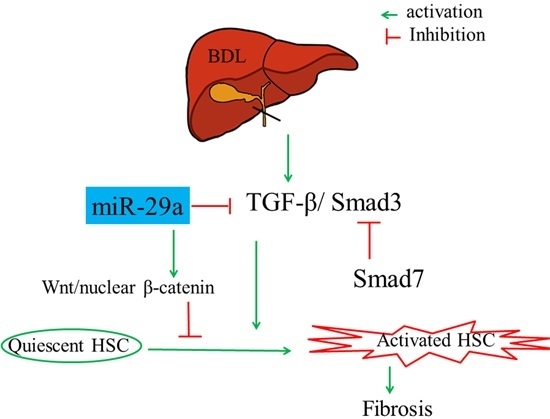
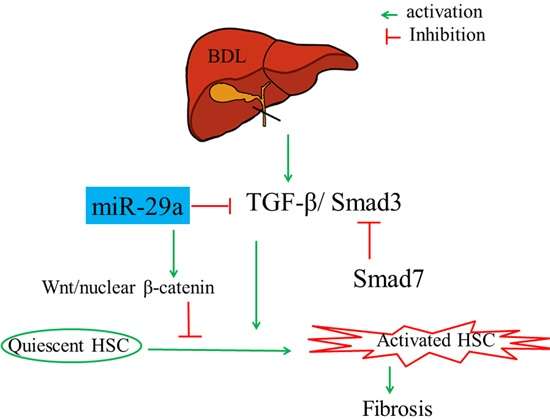
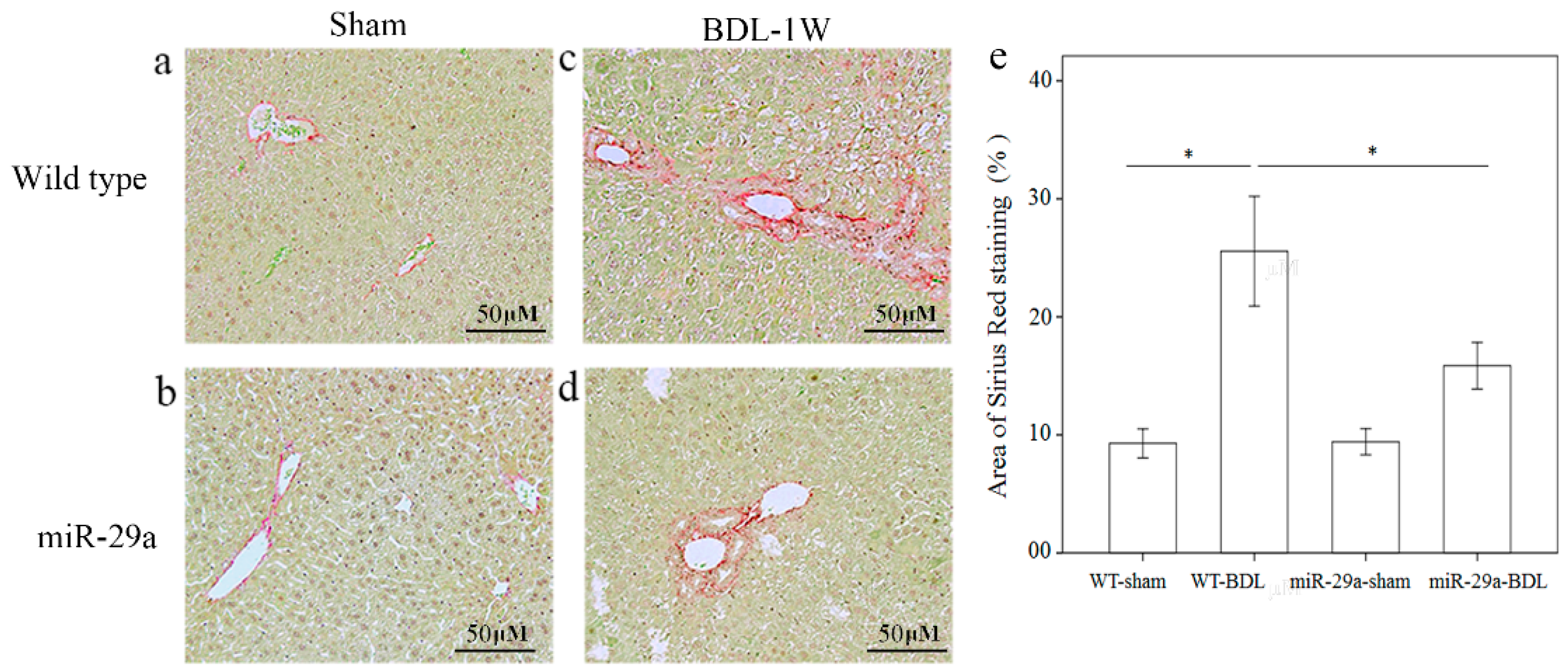
2.1. Gain of miR-29a Signaling Considerably Mitigates Liver Fibrosis in Cholestatic Livers
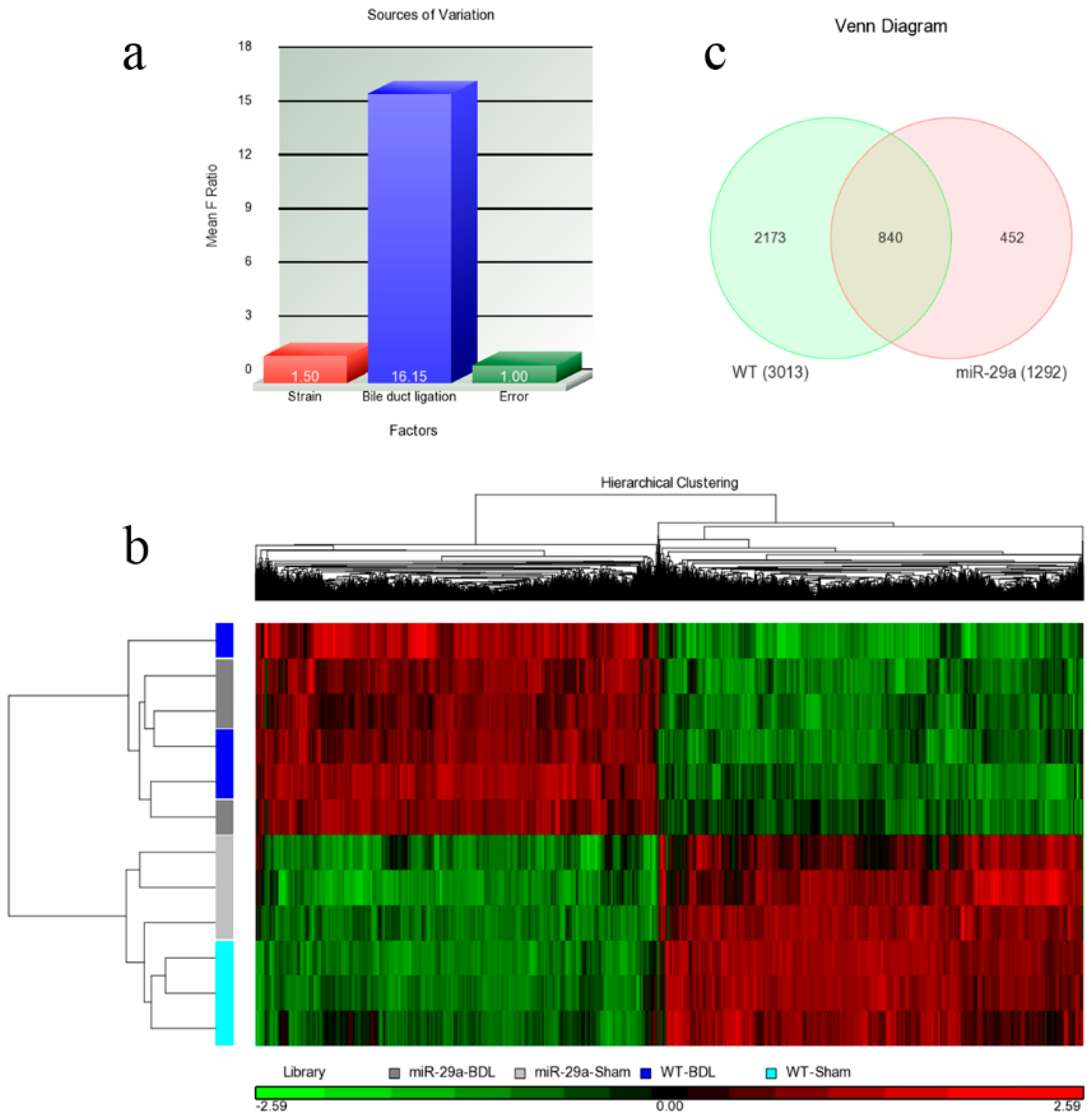
2.2. Analyses of Microarray Data
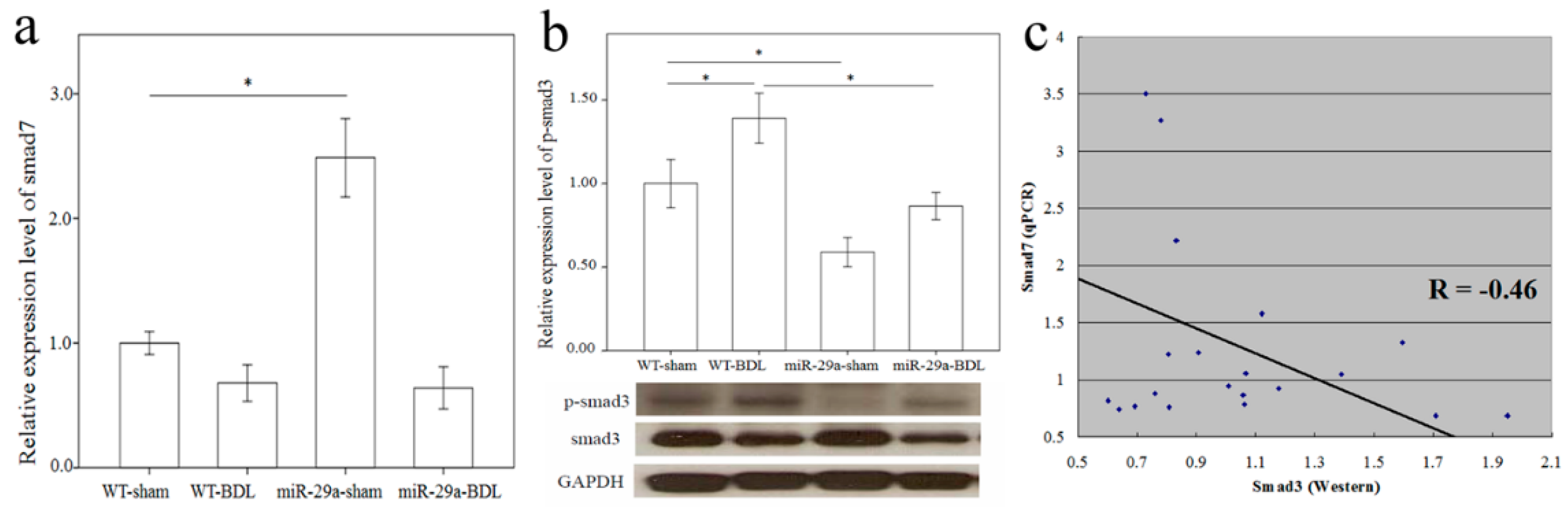
2.3. Overexpression of miR-29a Considerably Hinders TGF-β Signaling Pathways in Cholestatic Livers
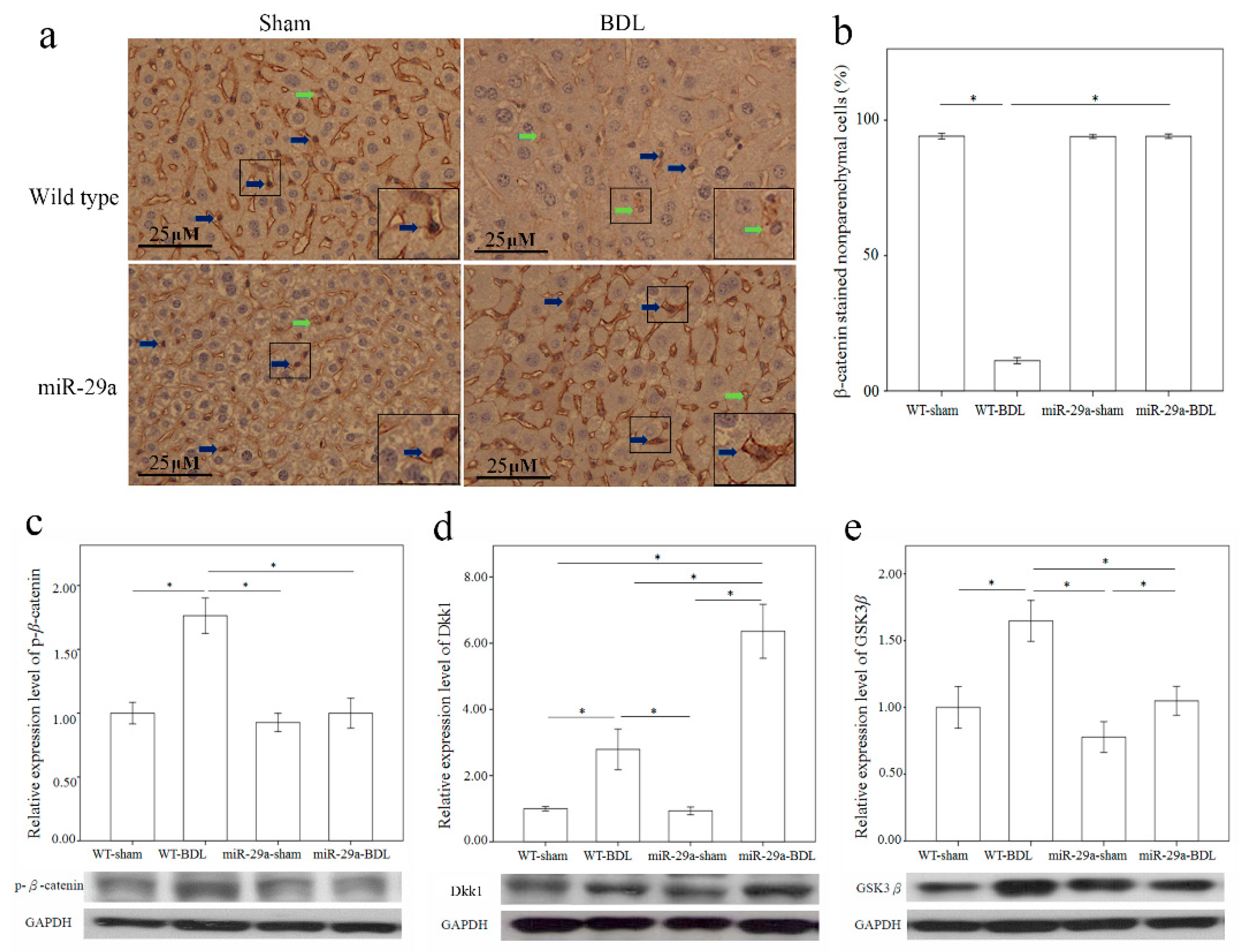
2.4. Overexpression of miR-29a Considerably Increased Wnt/β-Catenin in Cholestatic Livers
2.5. Overexpression of miR-29a Considerably Increased Dickkopf-Related Protein 1 (Dkk1) Expression in Cholestatic Livers
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Greation of miR-29a Transgenic Mice
4.3. Animal Model and Experimental Protocol
4.4. Histological Analysis
4.5. Microarray Samples
4.6. mRNA Expression Detected by Real-Time Quantitative RT-PCR
4.7. Immunoblotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Friedman, S.L. Mechanisms of hepatic fibrogenesis. Gastroenterology 2008, 134, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L. Evolving challenges in hepatic fibrosis. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Tiao, M.M.; Huang, L.T.; Chuang, J.H.; Kuo, K.C.; Yang, Y.L.; Wang, F.S. Activation of Mir-29a in activated hepatic stellate cells modulates its profibrogenic phenotype through inhibition of histone deacetylases 4. PLoS ONE 2015, 10, e0136453. [Google Scholar] [CrossRef] [PubMed]
- Tiao, M.M.; Wang, F.S.; Huang, L.T.; Chuang, J.H.; Kuo, H.C.; Yang, Y.L.; Huang, Y.H. MicroRNA-29a protects against acute liver injury in a mouse model of obstructive jaundice via inhibition of the extrinsic apoptosis pathway. Apoptosis 2014, 19, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Selbach, M.; Schwanhausser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Roderburg, C.; Urban, G.W.; Bettermann, K.; Vucur, M.; Zimmermann, H.; Schmidt, S.; Janssen, J.; Koppe, C.; Knolle, P.; Castoldi, M.; et al. Micro-RNA profiling reveals a role for miR-29 in human and murine liver fibrosis. Hepatology 2011, 53, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Gressner, A.M.; Weiskirchen, R.; Breitkopf, K.; Dooley, S. Roles of TGF-β in hepatic fibrosis. Front. Biosci. 2002, 7, d793–d807. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.F.; Fu, W.; Kim-Kaneyama, J.R.; Omoto, T.; Miyazaki, T.; Li, B.; Miyazaki, A. Hic-5 deficiency attenuates the activation of hepatic stellate cells and liver fibrosis through upregulation of Smad7 in mice. J. Hepatol. 2016, 64, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Afrakhte, M.; Moren, A.; Nakayama, T.; Christian, J.L.; Heuchel, R.; Itoh, S.; Kawabata, M.; Heldin, N.E.; Heldin, C.H.; et al. Identification of Smad7, a TGFβ-inducible antagonist of TGF-β signalling. Nature 1997, 389, 631–635. [Google Scholar] [PubMed]
- Yoshida, K.; Murata, M.; Yamaguchi, T.; Matsuzaki, K. TGF-β/Smad signaling during hepatic fibro-carcinogenesis (review). Int. J. Oncol. 2014, 45, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xiao, L.; Peng, Y.; Sun, L.; Liu, F. Wnt/β-catenin signaling: A promising new target for fibrosis diseases. Physiol. Res. 2012, 61, 337–346. [Google Scholar] [PubMed]
- Monga, S.P. β-Catenin Signaling and Roles in Liver Homeostasis, Injury, and Tumorigenesis. Gastroenterology 2015, 148, 1294–1310. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.L.; Khoo, P.L.; de Young, R.A.; Steiner, K.; Wilcock, C.; Mukhopadhyay, M.; Westphal, H.; Jamieson, R.V.; Robb, L.; Tam, P.P. Dkk1 and Wnt3 interact to control head morphogenesis in the mouse. Development 2008, 135, 1791–1801. [Google Scholar] [CrossRef] [PubMed]
- Habas, R.; Dawid, I.B. Dishevelled and Wnt signaling: Is the nucleus the final frontier? J. Biol. 2005, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, C.Y.; Nusse, R. The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 2004, 20, 781–810. [Google Scholar] [CrossRef] [PubMed]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. TGF-β signaling in fibrosis. Growth Factors 2011, 29, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Friedman, R.C.; Marquez, R.T.; Keck, K.; Kong, B.; Icardi, M.S.; Brown, K.E.; Burge, C.B.; Schmidt, W.N.; Wang, Y.; et al. Hepatitis C virus infection and hepatic stellate cell activation downregulate miR-29: miR-29 overexpression reduces hepatitis C viral abundance in culture. J. Infect. Dis. 2011, 203, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Gordillo, K.; Shah, R.; Arellanes-Robledo, J.; Hernandez-Nazara, Z.; Rincon-Sanchez, A.R.; Inagaki, Y.; Rojkind, M.; Lakshman, M.R. Mechanisms of action of acetaldehyde in the up-regulation of the human alpha2(I) collagen gene in hepatic stellate cells: Key roles of Ski, SMAD3, SMAD4, and SMAD7. Am. J. Pathol. 2014, 184, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Chung, A.C.; Huang, X.R.; Meng, X.M.; Hui, D.S.; Yu, C.M.; Sung, J.J.; Lan, H.Y. TGF-β/Smad3 signaling promotes renal fibrosis by inhibiting miR-29. J. Am. Soc. Nephrol. 2011, 22, 1462–1474. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.M.; Tang, P.M.; Li, J.; Lan, H.Y. TGF-β/Smad signaling in renal fibrosis. Front. Physiol. 2015, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Meng, X.M.; Huang, X.R.; Chung, A.C.; Feng, Y.L.; Hui, D.S.; Yu, C.M.; Sung, J.J.; Lan, H.Y. miR-29 inhibits bleomycin-induced pulmonary fibrosis in mice. Mol. Ther. 2012, 20, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.R. The Wnts. Genome Biol. 2002, 3, 3001.1–3001.15. [Google Scholar]
- Cheng, J.H.; She, H.; Han, Y.P.; Wang, J.; Xiong, S.; Asahina, K.; Tsukamoto, H. Wnt antagonism inhibits hepatic stellate cell activation and liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G39–G49. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhu, C.; Chen, X.; Li, Y.; Gao, R.; Wu, Q. Pokeweed antiviral protein down-regulates Wnt/β-catenin signalling to attenuate liver fibrogenesis in vitro and in vivo. Dig. Liver Dis. 2011, 43, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Kordes, C.; Sawitza, I.; Haussinger, D. Canonical Wnt signaling maintains the quiescent stage of hepatic stellate cells. Biochem. Biophys. Res. Commun. 2008, 367, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dai, C.; Li, Y.; Liu, Y. Canonical Wnt/β-catenin signaling mediates transforming growth factor-β1-driven podocyte injury and proteinuria. Kidney Int. 2011, 80, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Sato, M. Upregulation of the Wnt/β-catenin pathway induced by transforming growth factor-β in hypertrophic scars and keloids. Acta Derm. Venereol. 2006, 86, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.S.; Chung, P.C.; Lin, C.L.; Chen, M.W.; Ke, H.J.; Chang, Y.H.; Chen, Y.S.; Wu, S.L.; Ko, J.Y. MicroRNA-29a protects against glucocorticoid-induced bone loss and fragility in rats by orchestrating bone acquisition and resorption. Arthritis Rheum. 2013, 65, 1530–1540. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.C.; Chang, H.H.; Tsai, C.Y.; Jong, Y.J.; Horng, L.S.; Lin, S.F.; Tsai, T.F. Coat color-tagged green mouse with EGFP expressed from the RNA polymerase II promoter. Genesis 2004, 39, 122–129. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway Name | p Value | # of Genes in the Pathway | Pathway ID |
|---|---|---|---|
| Metabolic pathways | 0.000001 | 145 | kegg_pathway_235 |
| Peroxisome | 0.000229 | 19 | kegg_pathway_157 |
| Cell cycle | 0.000548 | 24 | kegg_pathway_53 |
| Retinol metabolism | 0.000743 | 16 | kegg_pathway_57 |
| Steroid hormone biosynthesis | 0.000904 | 14 | kegg_pathway_294 |
| Protein digestion and absorption | 0.001179 | 16 | kegg_pathway_206 |
| Arachidonic acid metabolism | 0.002021 | 15 | kegg_pathway_38 |
| Biosynthesis of antibiotics | 0.002177 | 31 | kegg_pathway_274 |
| Lysine degradation | 0.003177 | 13 | kegg_pathway_203 |
| Fatty acid degradation | 0.004790 | 11 | kegg_pathway_125 |
| TGF-β signaling pathway | 0.009331 | 15 | kegg_pathway_93 |
| Linoleic acid metabolism | 0.010929 | 8 | kegg_pathway_263 |
| Amoebiasis | 0.011037 | 18 | kegg_pathway_111 |
| Tryptophan metabolism | 0.012916 | 9 | kegg_pathway_88 |
| Progesterone-mediated oocyte maturation | 0.014029 | 16 | kegg_pathway_200 |
| ECM-receptor interaction | 0.015281 | 14 | kegg_pathway_147 |
| Chemical carcinogenesis | 0.018625 | 12 | kegg_pathway_229 |
| Butanoate metabolism | 0.025619 | 6 | kegg_pathway_248 |
| Bile secretion | 0.027883 | 12 | kegg_pathway_291 |
| Oocyte meiosis | 0.029539 | 17 | kegg_pathway_230 |
| Small cell lung cancer | 0.030371 | 14 | kegg_pathway_190 |
| PPAR signaling pathway | 0.033269 | 13 | kegg_pathway_78 |
| Regulation of lipolysis in adipocytes | 0.037877 | 10 | kegg_pathway_255 |
| Histidine metabolism | 0.038958 | 5 | kegg_pathway_165 |
| Drug metabolism—other enzymes | 0.039071 | 8 | kegg_pathway_36 |
| Fatty acid metabolism | 0.046776 | 9 | kegg_pathway_27 |
| Fatty acid biosynthesis | 0.048322 | 4 | kegg_pathway_89 |
| Mineral absorption | 0.049102 | 8 | kegg_pathway_236 |
| Pathway Name | p Value | # of Genes in the Pathway | Pathway ID |
|---|---|---|---|
| Steroid hormone biosynthesis | 0.000021 | 8 | kegg_pathway_294 |
| Leukocyte transendothelial migration | 0.000029 | 11 | kegg_pathway_276 |
| Natural killer cell mediated cytotoxicity | 0.000347 | 10 | kegg_pathway_253 |
| Chemical carcinogenesis | 0.000356 | 7 | kegg_pathway_229 |
| Fc epsilon RI signaling pathway | 0.000914 | 7 | kegg_pathway_100 |
| Platelet activation | 0.001147 | 9 | kegg_pathway_23 |
| B cell receptor signaling pathway | 0.001389 | 7 | kegg_pathway_65 |
| Osteoclast differentiation | 0.002132 | 9 | kegg_pathway_135 |
| Arrhythmogenic right ventricular cardiomyopathy (ARVC) | 0.002642 | 6 | kegg_pathway_1 |
| Retinol metabolism | 0.002727 | 6 | kegg_pathway_57 |
| Bile secretion | 0.002771 | 6 | kegg_pathway_291 |
| Phagosome | 0.003856 | 10 | kegg_pathway_194 |
| Hypertrophic cardiomyopathy (HCM) | 0.005919 | 6 | kegg_pathway_279 |
| Energy metabolism | 0.011458 | 8 | kegg_pathway_16 |
| Pentose and glucuronate interconversions | 0.021195 | 3 | kegg_pathway_285 |
| Metabolic pathways | 0.021733 | 31 | kegg_pathway_235 |
| Focal adhesion | 0.022124 | 9 | kegg_pathway_187 |
| Inositol phosphate metabolism | 0.022784 | 4 | kegg_pathway_49 |
| Viral carcinogenesis | 0.024087 | 9 | kegg_pathway_216 |
| Axon guidance | 0.025580 | 7 | kegg_pathway_264 |
| Antigen processing and presentation | 0.025778 | 5 | kegg_pathway_269 |
| Viral myocarditis | 0.026037 | 5 | kegg_pathway_47 |
| Linoleic acid metabolism | 0.027978 | 3 | kegg_pathway_263 |
| Dilated cardiomyopathy | 0.029280 | 5 | kegg_pathway_277 |
| Autoimmune thyroid disease | 0.039891 | 4 | kegg_pathway_64 |
| Biosynthesis of antibiotics | 0.040668 | 8 | kegg_pathway_274 |
| Fc gamma R-mediated phagocytosis | 0.041882 | 5 | kegg_pathway_120 |
| 2-Oxocarboxylic acid metabolism | 0.046164 | 2 | kegg_pathway_35 |
| HTLV-I infection | 0.047823 | 10 | kegg_pathway_189 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.-C.; Wang, F.-S.; Yang, Y.-L.; Tiao, M.-M.; Chuang, J.-H.; Huang, Y.-H. Microarray Study of Pathway Analysis Expression Profile Associated with MicroRNA-29a with Regard to Murine Cholestatic Liver Injuries. Int. J. Mol. Sci. 2016, 17, 324. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030324
Li S-C, Wang F-S, Yang Y-L, Tiao M-M, Chuang J-H, Huang Y-H. Microarray Study of Pathway Analysis Expression Profile Associated with MicroRNA-29a with Regard to Murine Cholestatic Liver Injuries. International Journal of Molecular Sciences. 2016; 17(3):324. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030324
Chicago/Turabian StyleLi, Sung-Chou, Feng-Sheng Wang, Ya-Ling Yang, Mao-Meng Tiao, Jiin-Haur Chuang, and Ying-Hsien Huang. 2016. "Microarray Study of Pathway Analysis Expression Profile Associated with MicroRNA-29a with Regard to Murine Cholestatic Liver Injuries" International Journal of Molecular Sciences 17, no. 3: 324. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030324