
Biodegradation of Single-Walled Carbon Nanotubes in Macrophages through Respiratory Burst Modulation

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
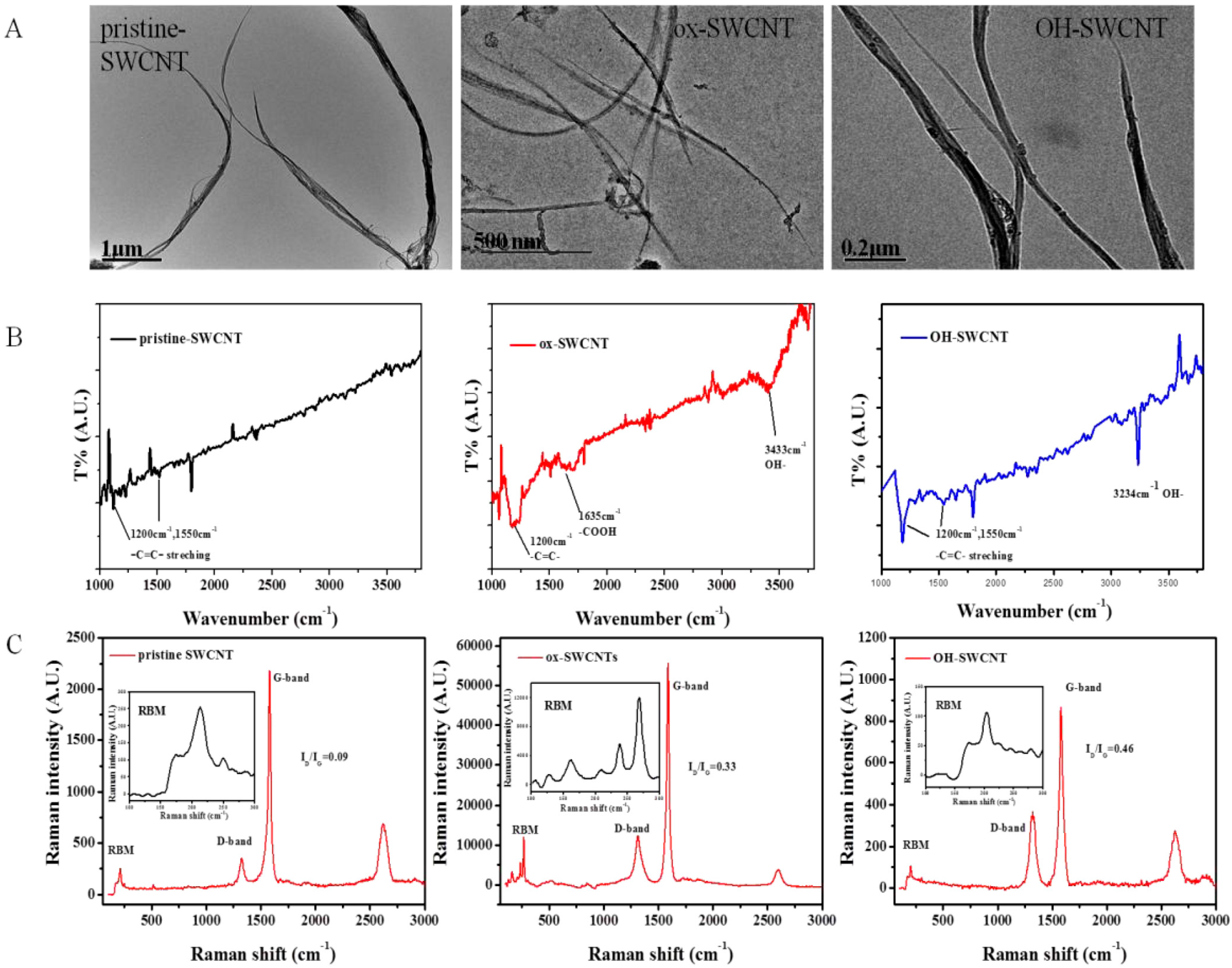
2.1. Characterization of Single-Walled Carbon Nanotubes (SWCNTs)
2.2. Biodegradation of SWCNTs in Macrophages
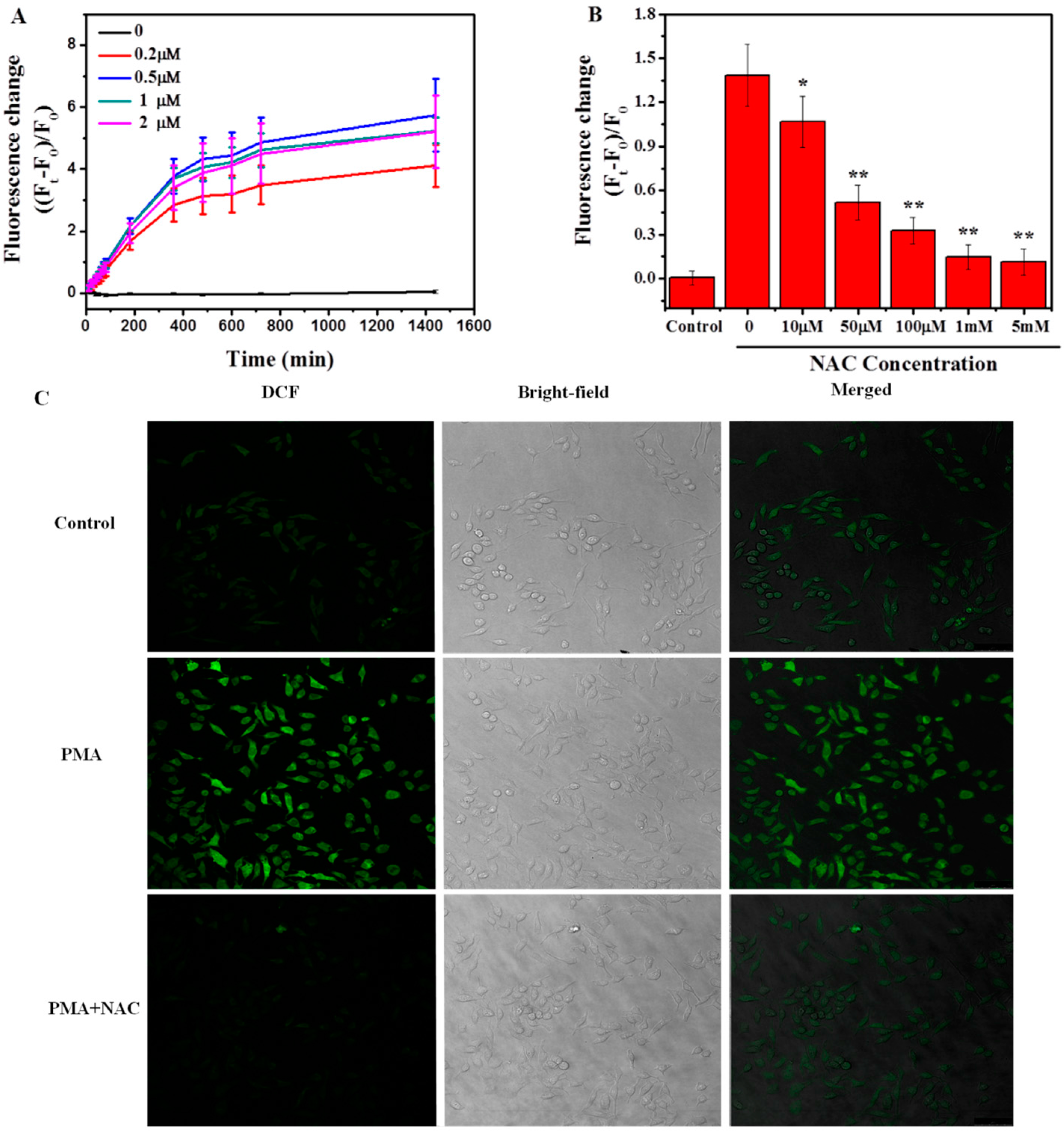
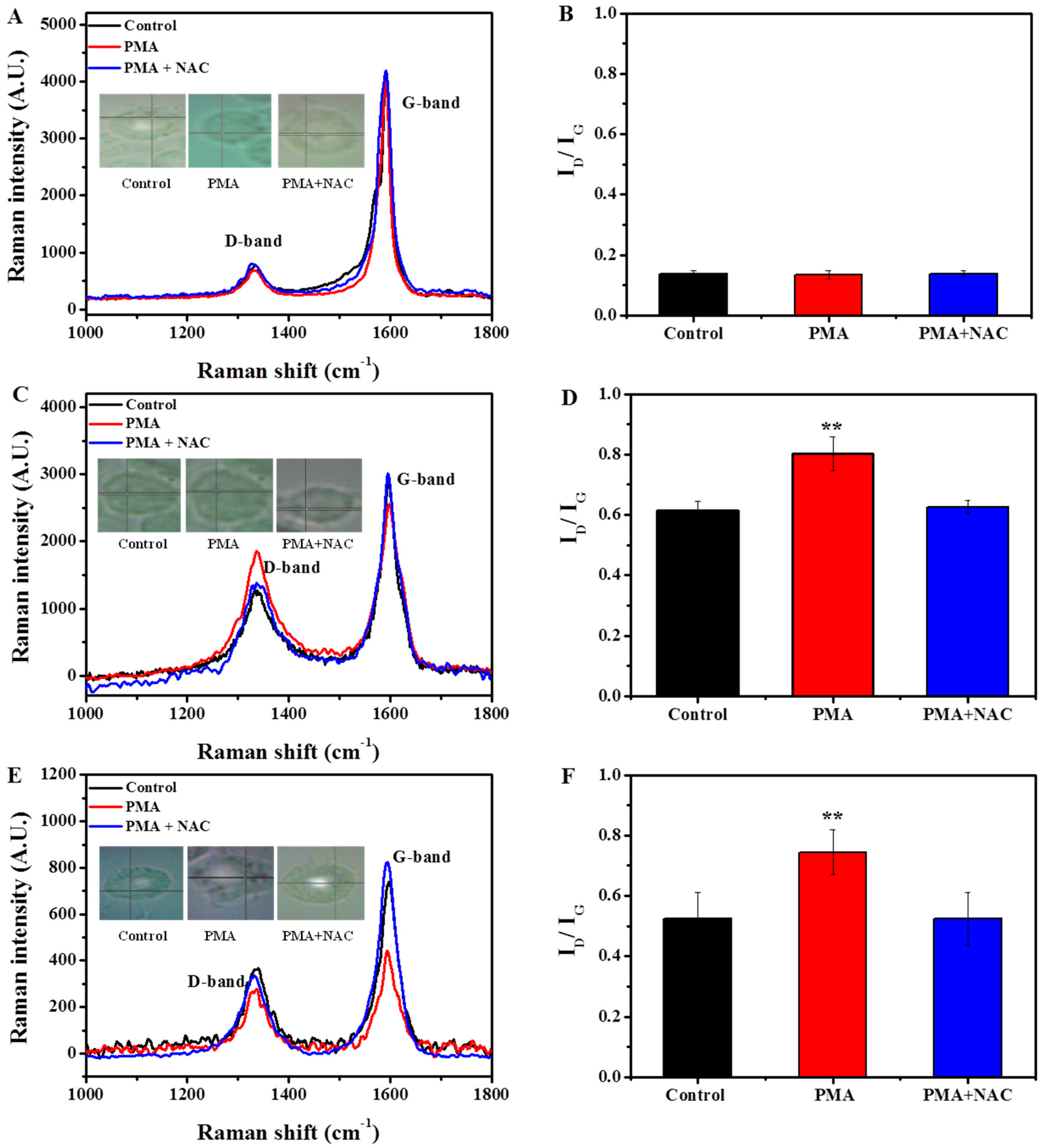
2.2.1. Induction of Respiratory Burst in Macrophages
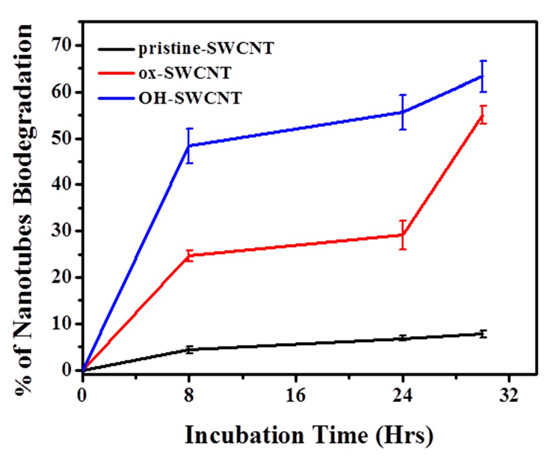
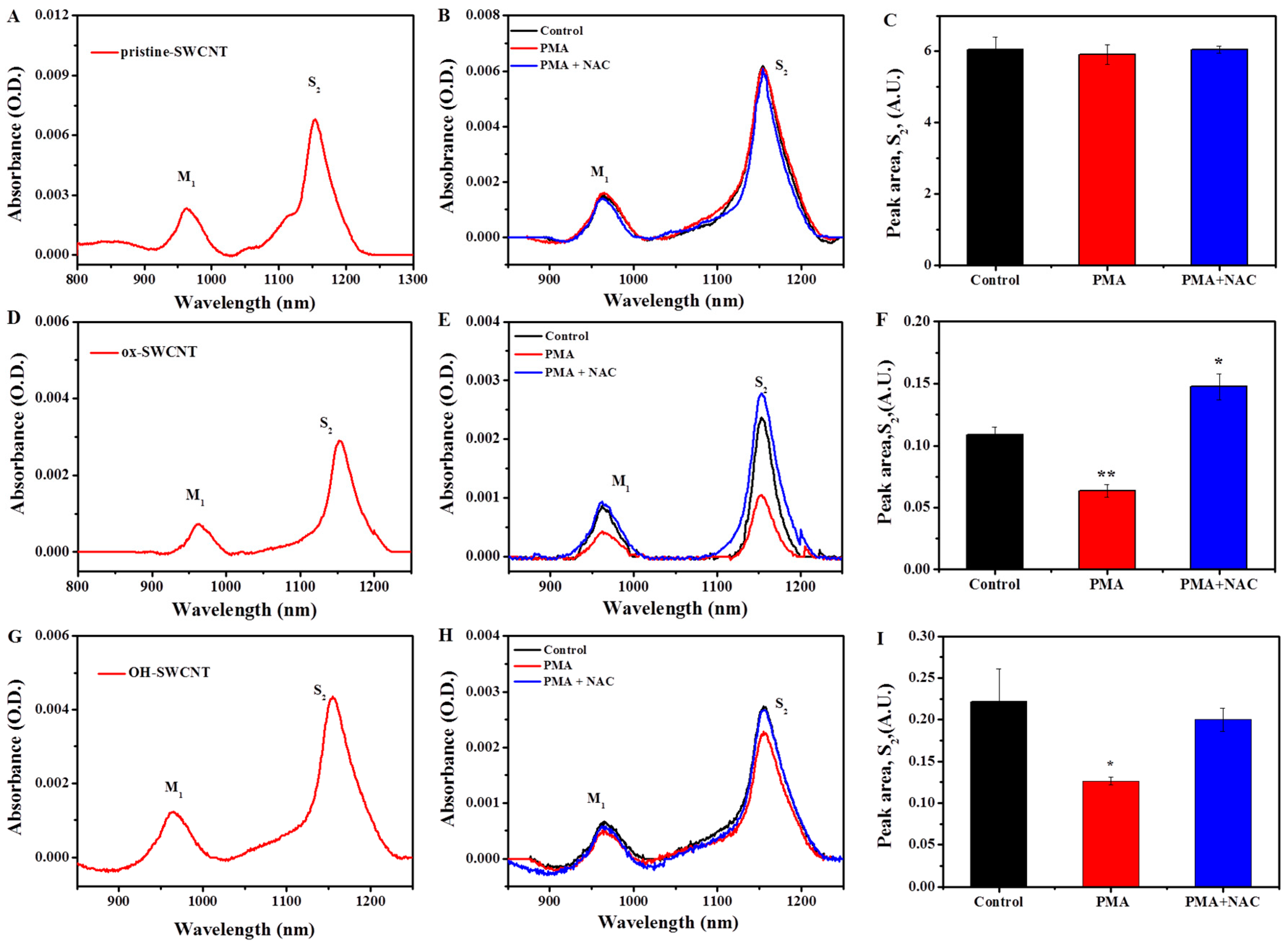
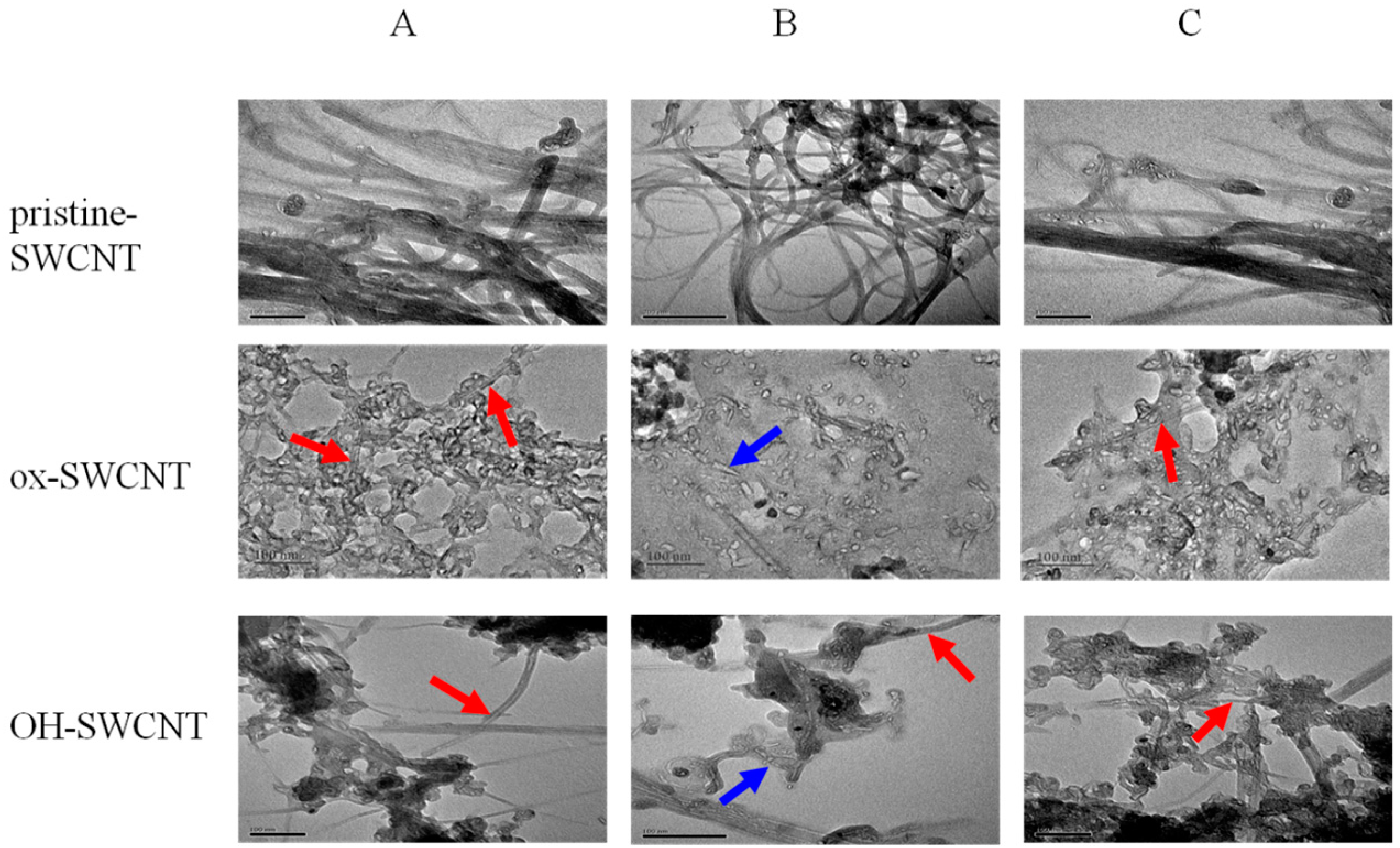
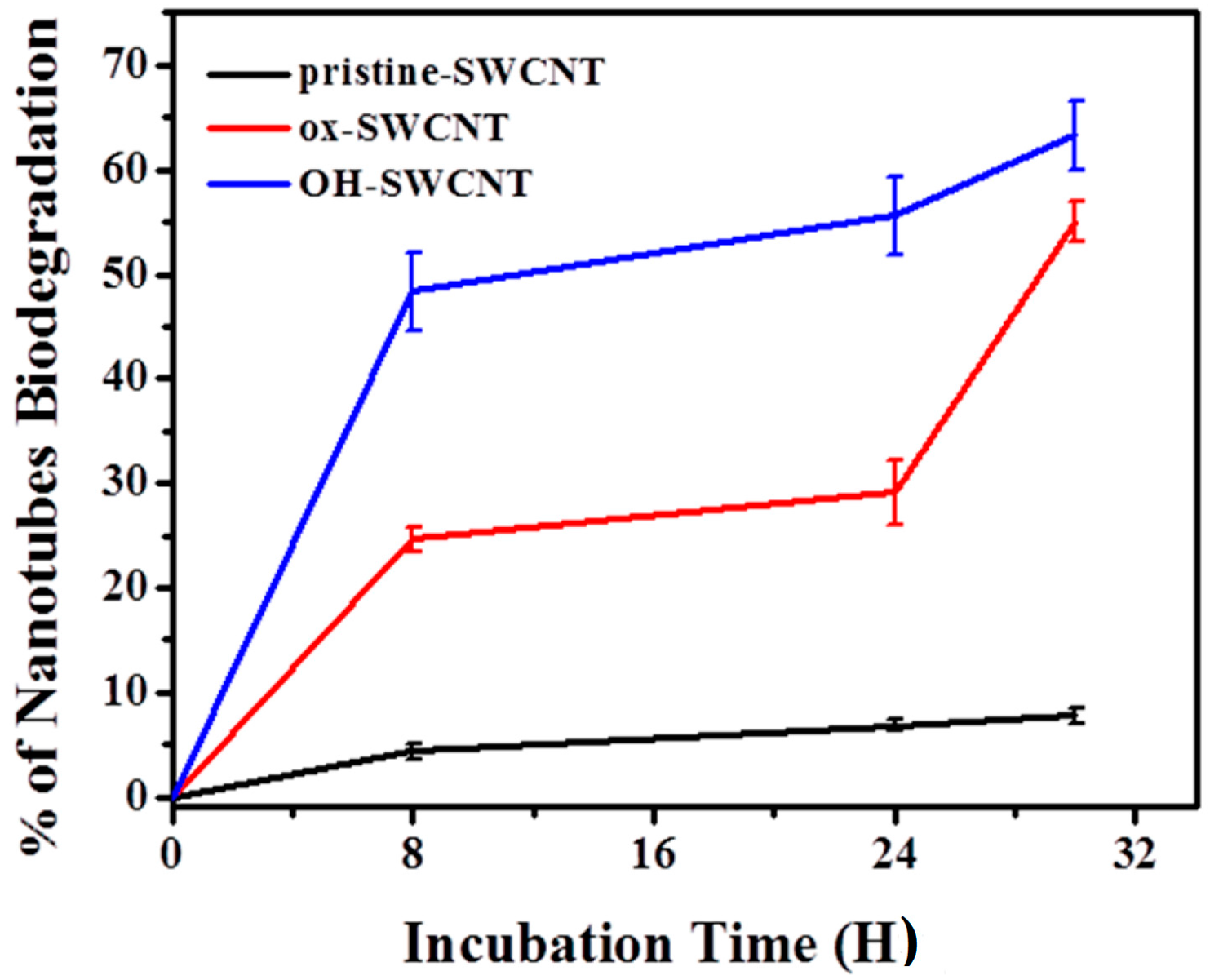
2.2.2. Characterization of SWCNTs Degradation in Macrophages
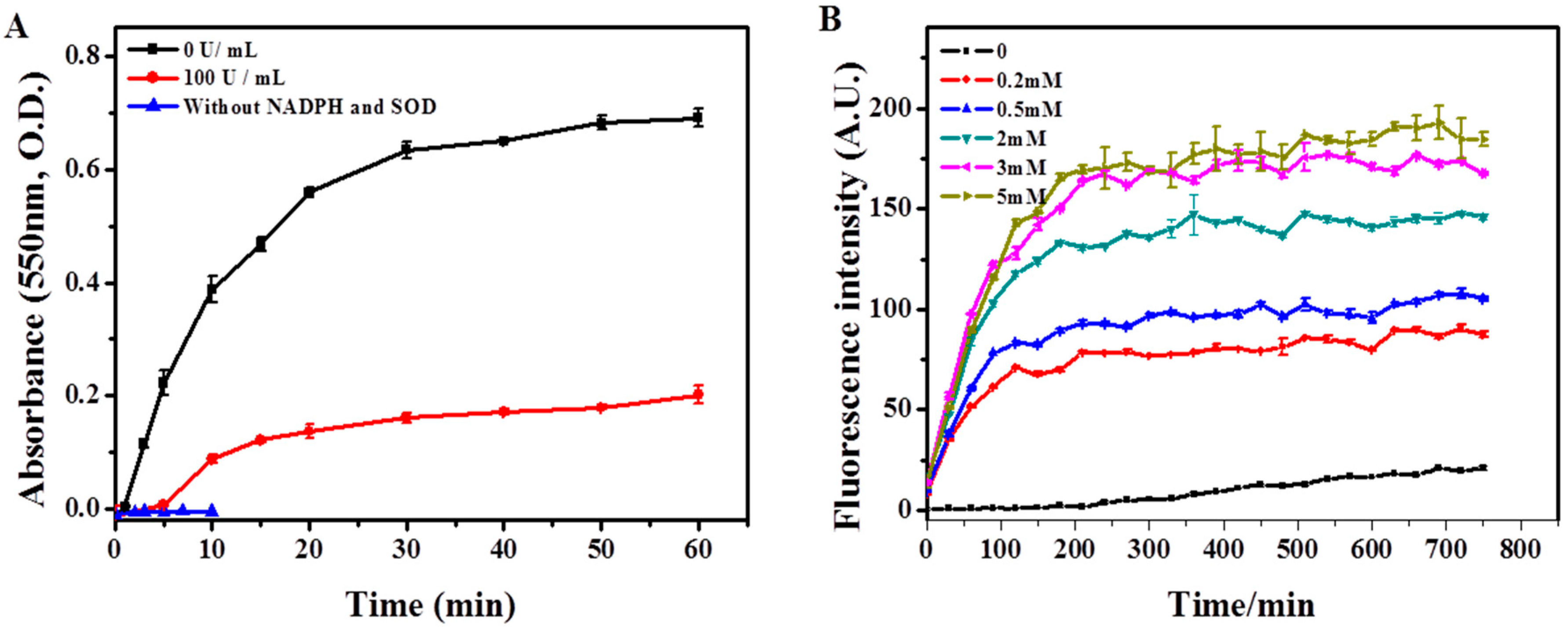
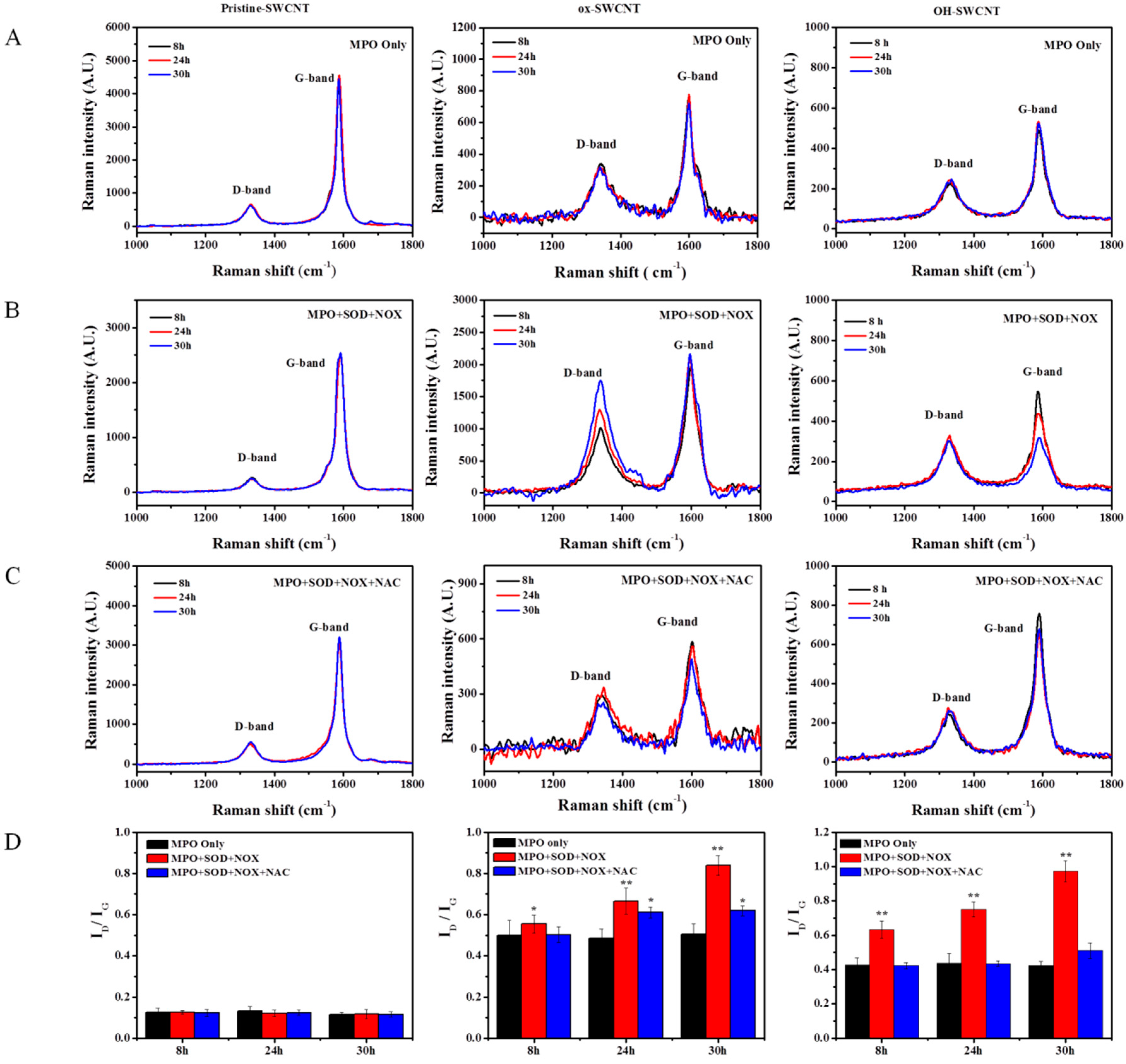
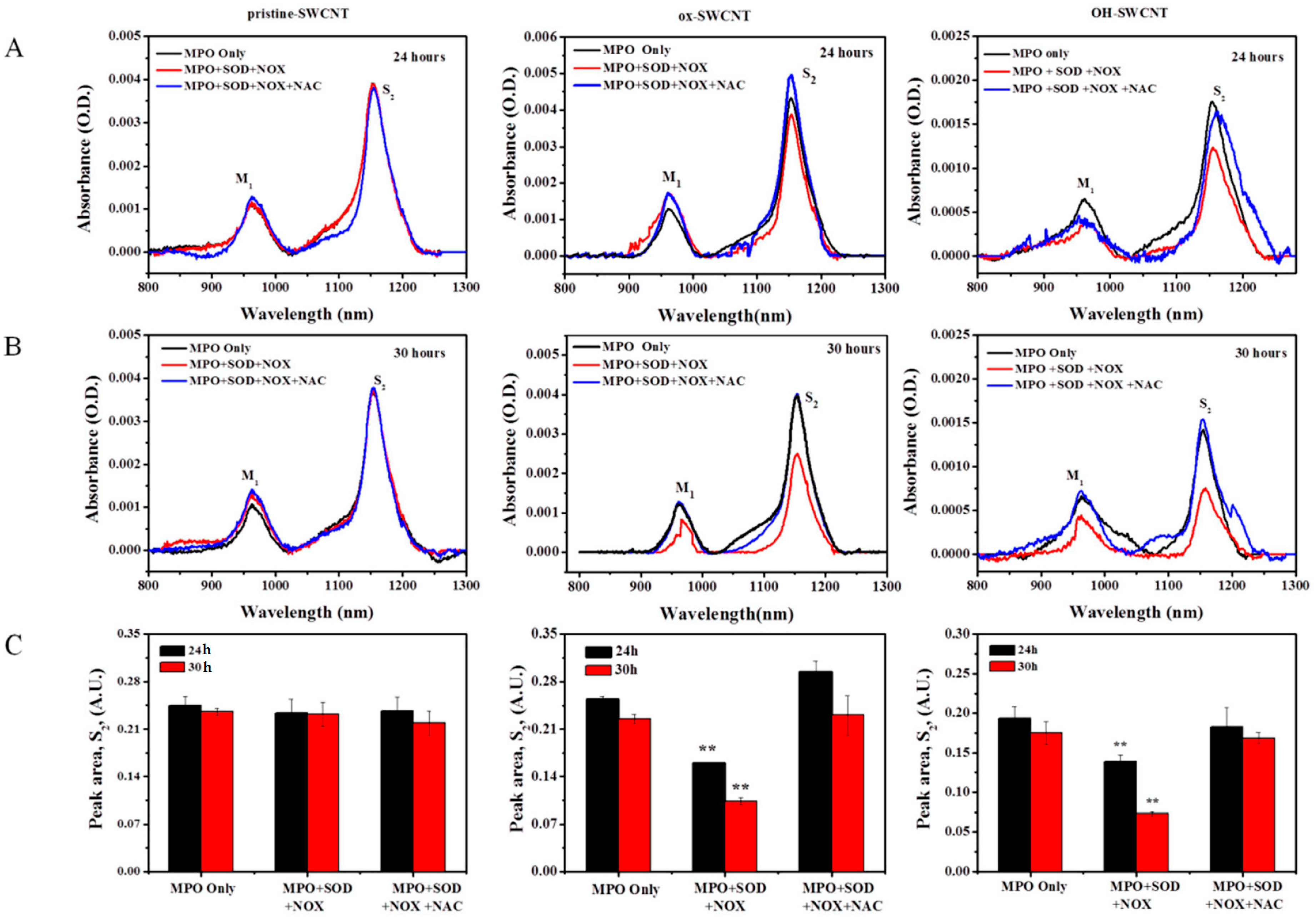
2.3. Biodegradation of SWCNTs in Respiratory Burst Enzymatic System in Vitro
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of SWCNTs Solutions and Characterization
4.3. Cell Culture and Treatment
4.4. Construction of in Vitro Respiratory Burst Enzymatic System and Verification
4.5. Assessment of SWCNT Degradation
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SWCNT | Single-walled carbon nanotube |
| HRP | Horse radish peroxidase |
| MPO | Myeloperoxidase |
| EPO | Eosinophil peroxidase |
| NETs | Neutrophil extracellular traps |
| PMN | Polymorphonuclear neutrophils |
| O2*− | Superoxide anion |
| NOX | Nicotinamide adenine dinucleotide phosphate (NADPH)-oxidase |
| SOD | Superoxide dismutase |
| PMA | Phorbol myristate acetate |
| LPS | Lipopolysaccharide |
| NAC | N-acetyl-l-cysteine |
| ROS | Reactive oxygen species |
| DCF-DA | 2′,7′-Dichlorofluorescein diacetate |
| TEM | Transmission electron microscopy |
| UV-vis-NIR | Ultraviolet-visible-near-infrared |
| FAD | Flavin adenine dinucleotide disodium salt hydrate |
References
- De Volder, M.F.L.; Tawfick, S.H.; Baughman, R.H.; Hart, A.J. Carbon nanotubes: Present and future commercial applications. Science 2013, 339, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Marchesan, S.; Kostarelos, K.; Bianco, A.; Prato, M. The winding road for carbon nanotubes in nanomedicine. Mater. Today 2015, 18, 12–19. [Google Scholar] [CrossRef]
- Marchesan, S.; Melchionna, M.; Prato, M. Wire up on carbon nanostructures! How to play a winning game. ACS Nano 2015, 9, 9441–9450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, M.; Solati, N.; Amiri, M.; Mirshekari, H.; Mohamed, E.; Taheri, M.; Hashemkhani, M.; Saeidi, A.; Estiar, M.A.; Kiani, P.; et al. Carbon nanotubes part I: Preparation of a novel and versatile drug-delivery vehicle. Expert Opin. Drug Deliv. 2015, 12, 1071–1087. [Google Scholar] [CrossRef] [PubMed]
- Martincic, M.; Tobias, G. Filled carbon nanotubes in biomedical imaging and drug delivery. Expert Opin. Drug Deliv. 2015, 12, 563–581. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kis, A.; Zettl, A.; Bertozzi, C.R. A cell nanoinjector based on carbon nanotubes. Proc. Natl. Acad. Sci. USA 2007, 104, 8218–8222. [Google Scholar] [CrossRef] [PubMed]
- Malarkey, E.B.; Parpura, V. Applications of carbon nanotubes in neurobiology. Neurodegener. Dis. 2007, 4, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.; Solati, N.; Ghasemi, A.; Estiar, M.A.; Hashemkhani, M.; Kiani, P.; Mohamed, E.; Saeidi, A.; Taheri, M.; Avci, P.; et al. Carbon nanotubes part II: A remarkable carrier for drug and gene delivery. Expert Opin. Drug Deliv. 2015, 12, 1089–1105. [Google Scholar] [CrossRef] [PubMed]
- Prato, M.; Kostarelos, K.; Bianco, A. Functionalized carbon nanotubes in drug design and discovery. Acc. Chem. Res. 2008, 41, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.S.; Yoong, S.L.; Jagusiak, A.; Panczyk, T.; Ho, H.K.; Ang, W.H.; Pastorin, G. Carbon nanotubes for delivery of small molecule drugs. Adv. Drug Deliv. Rev. 2013, 65, 1964–2015. [Google Scholar] [CrossRef] [PubMed]
- Lamberti, M.; Zappavigna, S.; Sannolo, N.; Porto, S.; Caraglia, M. Advantages and risks of nanotechnologies in cancer patients and occupationally exposed workers. Expert Opin. Drug Deliv. 2014, 11, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Schulte, P.A.; Kuempel, E.D.; Zumwalde, R.D.; Geraci, C.L.; Schubauer-Berigan, M.K.; Castranova, V.; Hodson, L.; Murashov, V.; Dahm, M.M.; Ellenbecker, M. Focused actions to protect carbon nanotube workers. Am. J. Ind. Med. 2012, 55, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Maynard, A.D.; Kuempel, E.D. Airborne nanostructured particles and occupational health. J. Nanopart. Res. 2005, 7, 587–614. [Google Scholar] [CrossRef]
- Porter, A.E.; Gass, M.; Muller, K.; Skepper, J.N.; Midgley, P.A.; Welland, M. Direct imaging of single-walled carbon nanotubes in cells. Nat. Nanotechnol. 2007, 2, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Liu, Y.; Dolash, B.D.; Jung, Y.; Slipchenko, M.N.; Bergstrom, D.E.; Cheng, J.X. Label-free imaging of semiconducting and metallic carbon nanotubes in cells and mice using transient absorption microscopy. Nat. Nanotechnol. 2012, 7, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Al-Jamal, K.T.; Nunes, A.; Methven, L.; Ali-Boucetta, H.; Li, S.; Toma, F.M.; Herrero, M.A.; Al-Jamal, W.T.; ten Eikelder, H.M.M.; Foster, J.; Mather, S.; Prato, M.; et al. Degree of chemical functionalization of carbon nanotubes determines tissue distribution and excretion profile. Angew. Chem. Int. Ed. 2012, 51, 6389–6393. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.W.; James, J.T.; McCluskey, R.; Hunter, R.L. Pulmonary toxicity of single-wall carbon nanotubes in mice 7 and 90 days after intratracheal instillation. Toxicol. Sci. 2004, 77, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.H.; von dem Bussche, A.; Hurt, R.H.; Kane, A.B.; Gao, H.J. Cell entry of one-dimensional nanomaterials occurs by tip recognition and rotation. Nat. Nanotechnol. 2011, 6, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Wang, H.F.; Yan, L.; Wang, X.; Pei, R.J.; Yan, T.; Zhao, Y.L.; Guo, X.B. Cytotoxicity of carbon nanomaterials: Single-wall nanotube, multi-wall nanotube, and fullerene. Environ. Sci. Technol. 2005, 39, 1378–1383. [Google Scholar] [CrossRef] [PubMed]
- Magrez, A.; Kasas, S.; Salicio, V.; Pasquier, N.; Seo, J.W.; Celio, M.; Catsicas, S.; Schwaller, B.; Forro, L. Cellular toxicity of carbon-based nanomaterials. Nano Lett. 2006, 6, 1121–1125. [Google Scholar] [CrossRef] [PubMed]
- Firme, C.P.; Bandaru, P.R. Toxicity issues in the application of carbon nanotubes to biological systems. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Warheit, D.B.; Laurence, B.R.; Reed, K.L.; Roach, D.H.; Reynolds, G.A.M.; Webb, T.R. Comparative pulmonary toxicity assessment of single-wall carbon nanotubes in rats. Toxicol. Sci. 2004, 77, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Shvedova, A.A.; Kisin, E.; Murray, A.R.; Johnson, V.J.; Gorelik, O.; Arepalli, S.; Hubbs, A.F.; Mercer, R.R.; Keohavong, P.; Sussman, N.; et al. Inhalation vs. aspiration of single-walled carbon nanotubes in C57BL/6 mice: Inflammation, fibrosis, oxidative stress, and mutagenesis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L552–L565. [Google Scholar] [CrossRef] [PubMed]
- Ali-Boucetta, H.; Nunes, A.; Sainz, R.; Herrero, M.A.; Tian, B.; Prato, M.; Bianco, A.; Kostarelos, K. Asbestos-like pathogenicity of long carbon nanotubes alleviated by chemical functionalization. Angew. Chem. Int. Ed. 2013, 52, 2274–2278. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Tyurina, Y.Y.; Tyurin, V.A.; Konduru, N.V.; Potapovich, A.I.; Osipov, A.N.; Kisin, E.R.; Schwegler-Berry, D.; Mercer, R.; Castranova, V.; et al. Direct and indirect effects of single walled carbon nanotubes on raw 264.7 macrophages: Role of iron. Toxicol. Lett. 2006, 165, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Kisin, E.R.; Murray, A.R.; Keane, M.J.; Shi, X.C.; Schwegler-Berry, D.; Gorelik, O.; Arepalli, S.; Castranova, V.; Wallace, W.E.; Kagan, V.E.; et al. Single-walled carbon nanotubes: Geno- and cytotoxic effects in lung fibroblast v79 cells. J. Toxicol. Environ. Health A 2007, 70, 2071–2079. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Thakare, V.S.; Das, M.; Godugu, C.; Jain, A.K.; Mathur, R.; Chuttani, K.; Mishra, A.K. Toxicity of multiwalled carbon nanotubes with end defects critically depends on their functionalization density. Chem. Res. Toxicol. 2011, 24, 2028–2039. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Zhang, D.; Li, L.; Shen, X.; Deng, X.; Dong, L.; Wu, M.; Liu, Y. The hepattoxicity of multi-walled carbon nanotubes in mice. Nanotechnology 2009, 20. [Google Scholar] [CrossRef] [PubMed]
- Sureshbabu, A.R.; Kurapati, R.; Russier, J.; Menard-Moyon, C.; Bartolini, I.; Meneghetti, M.; Kostarelos, K.; Bianco, A. Degradation-by-design: Surface modification with functional substrates that enhance the enzymatic degradation of carbon nanotubes. Biomaterials 2015, 72, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Kotchey, G.P.; Zhao, Y.; Kagan, V.E.; Star, A. Peroxidase-mediated biodegradation of carbon nanotubes in vitro and in vivo. Adv. Drug Deliv. Rev. 2013, 65, 1921–1932. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.L.; Kichambare, P.D.; Gou, P.; Vlasova, I.I.; Kapralov, A.A.; Konduru, N.; Kagan, V.E.; Star, A. Biodegradation of single-walled carbon nanotubes through enzymatic catalysis. Nano Lett. 2008, 8, 3899–3903. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.L.; Kotchey, G.P.; Chen, Y.; Yanamala, N.V.K.; Klein-Seetharaman, J.; Kagan, V.E.; Star, A. Mechanistic investigations of horseradish peroxidase-catalyzed degradation of single-walled carbon nanotubes. J. Am. Chem. Soc. 2009, 131, 17194–17205. [Google Scholar] [CrossRef] [PubMed]
- Russier, J.; Menard-Moyon, C.; Venturelli, E.; Gravel, E.; Marcolongo, G.; Meneghetti, M.; Doris, E.; Bianco, A. Oxidative biodegradation of single- and multi-walled carbon nanotubes. Nanoscale 2011, 3, 893–896. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Hurt, R.H.; Kane, A.B. Biodurability of single-walled carbon nanotubes depends on surface functionalization. Carbon 2010, 48, 1961–1969. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Konduru, N.V.; Feng, W.; Allen, B.L.; Conroy, J.; Volkov, Y.; Vlasova, I.I.; Belikova, N.A.; Yanamala, N.; Kapralov, A.; et al. Carbon nanotubes degraded by neutrophil myeloperoxidase induce less pulmonary inflammation. Nat. Nanotechnol. 2010, 5, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Andon, F.T.; Kapralov, A.A.; Yanamala, N.; Feng, W.; Baygan, A.; Chambers, B.J.; Hultenby, K.; Ye, F.; Toprak, M.S.; Brandner, B.D.; et al. Biodegradation of single-walled carbon nanotubes by eosinophil peroxidase. Small 2013, 9, 2721–2729. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, I.I.; Sokolov, A.V.; Chekanov, A.V.; Kostevich, V.A.; Vasilyev, V.B. Myeloperoxidase-induced biodegradation of single-walled carbon nanotubes is mediated by hypochlorite. Russ. J. Bioorg. Chem. 2011, 37, 453–463. [Google Scholar] [CrossRef]
- Chandrasekaran, G.; Choi, S.K.; Lee, Y.C.; Kim, G.J.; Shin, H.J. Oxidative biodegradation of single-walled carbon nanotubes by partially purified lignin peroxidase from Sparassis latifolia mushroom. J. Ind. Eng. Chem. 2013, 20, 3367–3374. [Google Scholar] [CrossRef]
- Zhang, L.W.; Petersen, E.J.; Habteselassie, M.Y.; Mao, L.; Huang, Q.G. Degradation of multiwall carbon nanotubes by bacteria. Environ. Pollut. 2013, 181, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Farrera, C.; Bhattacharya, K.; Lazzaretto, B.; Andon, F.T.; Hultenby, K.; Kotchey, G.P.; Star, A.; Fadeel, B. Extracellular entrapment and degradation of single-walled carbon nanotubes. Nanoscale 2014, 6, 6974–6983. [Google Scholar] [CrossRef] [PubMed]
- Teeguarden, J.G.; Webb-Robertson, B.J.; Waters, K.M.; Murray, A.R.; Kisin, E.R.; Varnum, S.M.; Jacobs, J.M.; Pounds, J.G.; Zanger, R.C.; Shvedova, A.A. Comparative proteomics and pulmonary toxicity of instilled single-walled carbon nanotubes, crocidolite asbestos, and ultrafine carbon black in mice. Toxicol. Sci. 2011, 120, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Elgrabli, D.; Floriani, M.; Abella-Gallart, S.; Meunier, L.; Gamez, C.; Delalain, P.; Rogerieux, F.; Boczkowski, J.; Lacroix, G. Biodistribution and clearance of instilled carbon nanotubes in rat lung. Part. Fibre Toxicol. 2008, 5. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.; Bussy, C.; Gherardini, L.; Meneghetti, M.; Herrero, M.A.; Bianco, A.; Prato, M.; Pizzorusso, T.; Al-Jamal, K.T.; Kostarelos, K. In vivo degradation of functionalized carbon nanotubes after stereotactic administration in the brain cortex. Nanomedicine 2012, 7, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M.; Lambeth, J.D.; Nauseef, W. The neutrophil NADPH oxidase. Arch. Biochem. Biophys. 2002, 397, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Inoguchi, T.; Sonta, T.; Tsubouchi, H.; Etoh, T.; Kakimoto, M.; Sonoda, N.; Sato, N.; Sekiguchi, N.; Kobayashi, K.; Sumimoto, H.; et al. Protein kinase c-dependent increase in reactive oxygen species (ROS) production in vascular tissues of diabetes: Role of vascular NAD(P)H oxidase. J. Am. Soc. Nephrol. 2003, 14, S227–S232. [Google Scholar] [CrossRef] [PubMed]
- Curulli, A.; Cesaro, S.N.; Coppe, A.; Silvestri, C.; Palleschi, G. Functionalization and dissolution of single-walled carbon nanotubes by chemical-physical and electrochemical treatments. Microchim. Acta 2006, 152, 225–232. [Google Scholar] [CrossRef]
- Ahmad, M.N.; Xie, J.Y.; Ma, Y.H.; Yang, W.T. Surface functionalization of single-walled carbon nanotubes using photolysis for enhanced dispersion in an organic solvent. New Carbon Mater. 2010, 25, 134–140. [Google Scholar] [CrossRef]
- Zhang, L.; Kiny, V.U.; Peng, H.Q.; Zhu, J.; Lobo, R.F.M.; Margrave, J.L.; Khabashesku, V.N. Sidewall functionalization of single-walled carbon nanotubes with hydroxyl group-terminated moieties. Chem. Mater. 2004, 16, 2055–2061. [Google Scholar] [CrossRef]
- Octavia, Y.; Brunner-La Rocca, H.P.; Moens, A.L. NADPH oxidase-dependent oxidative stress in the failing heart: From pathogenic roles to therapeutic approach. Free Radic. Biol. Med. 2012, 52, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Morel, F.; Doussiere, J.; Vignais, P.V. The superoxide-generating oxidase of phagocytic-cells—Physiological, molecular and pathological aspects. Eur. J. Biochem. 1991, 201, 523–546. [Google Scholar] [CrossRef] [PubMed]
- Green, S.P.; Phillips, W.A. Activation of the macrophage respiratory burst by phorbol-myristate acetate—Evidence for both tyrosine kinase-dependent and kinase-independent pathways. Biochim. Biophys. Acta Mol. Cell Res. 1994, 1222, 241–248. [Google Scholar] [CrossRef]
- Decker, J.E.; Hight Walker, A.R.; Bosnick, K.; Clifford, C.A.; Dai, L.; Fagan, J.; Hooker, S.; Jakubek, Z.J.; Kingston, C.; Makar, J.; et al. Sample preparation protocols for realization of reproducible characterization of single-wall carbon nanotubes. Metrologia 2009, 46, 682–692. [Google Scholar] [CrossRef]
- Hamon, M.A.; Itkis, M.E.; Niyogi, S.; Alvaraez, T.; Kuper, C.; Menon, M.; Haddon, R.C. Effect of rehybridization on the electronic structure of single-walled carbon nanotubes. J. Am. Chem. Soc. 2001, 123, 11292–11293. [Google Scholar] [CrossRef] [PubMed]
- Bromberg, Y.; Pick, E. Activation of NADPH-dependent superoxide production in a cell-free system by sodium dodecyl-sulfate. J. Biol. Chem. 1985, 260, 3539–3545. [Google Scholar]
- Green, T.R.; Schaefer, R.E. Intrinsic dichlorophenolindophenol reductase-activity associated with the superoxide-generating oxidoreductase of human-granulocytes. Biochemistry 1981, 20, 7483–7487. [Google Scholar] [CrossRef] [PubMed]
- Gabig, T.G.; English, D.; Akard, L.P.; Schell, M.J. Regulation of neutrophil NADPH oxidase activation in a cell-free system by guanine nucleotides and fluoride. Evidence for participation of a pertussis and cholera toxin-insensitive g protein. J. Biol. Chem. 1987, 262, 1685–1690. [Google Scholar] [PubMed]
- Babior, B.M. NADPH oxidase: An update. Blood 1999, 93, 1464–1476. [Google Scholar] [PubMed]
- Nauseef, W.M. Nox enzymes in immune cells. Semin. Immunopathol. 2008, 30, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Bedard, K.; Krause, K.H. The nox family of ROS-generating NADPH oxidaes: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M. NADPH oxidase. Curr. Opin. Immunol 2004, 16, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Kotchey, G.P.; Gaugler, J.A.; Kapralov, A.A.; Kagan, V.E.; Star, A. Effect of antioxidants on enzyme-catalysed biodegradation of carbon nanotubes. J. Mat. Chem. B 2013, 1, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Paravicini, T.M.; Touyz, R.M. NADPH oxidases, reactive oxygen species, and hypertension: Clinical implications and therapeutic possibilities. Diabetes Care 2008, 31, S170–S180. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Kapralov, A.A.; St Croix, C.M.; Watkins, S.C.; Kisin, E.R.; Kotchey, G.P.; Balasubramanian, K.; Vlasova, I.I.; Yu, J.; Kim, K.; et al. Lung macrophages “digest” carbon nanotubes using a superoxide/peroxynitrite oxidative pathway. ACS Nano 2014, 8, 5610–5621. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Allen, B.L.; Star, A. Enzymatic degradation of multiwalled carbon nanotubes. J. Phys. Chem. A 2011, 115, 9536–9544. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Rinzler, A.G.; Dai, H.J.; Hafner, J.H.; Bradley, R.K.; Boul, P.J.; Lu, A.; Iverson, T.; Shelimov, K.; Huffman, C.B.; et al. Fullerene pipes. Science 1998, 280, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.X.; Wan, B.; Guo, L.H. In vitro toxicity of acid-functionalized single-walled carbon nanotubes: Effects on murine macrophages and gene expression profiling. Nanotoxicology 2012, 6, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Wan, B.; Wang, Z.X.; Lv, Q.Y.; Dong, P.X.; Zhao, L.X.; Yang, Y.; Guo, L.H. Single-walled carbon nanotubes and graphene oxides induce autophagosome accumulation and lysosome impairment in primarily cultured murine peritoneal macrophages. Toxicol. Lett. 2013, 221, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.H.; Liu, Z.X.; Shen, X. Roles of phospholipase d in phorbol myristate acetate-stimulated neutrophil respiratory burst. J. Cell. Mol. Med. 2011, 15, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Bellavite, P.; Berton, G.; Dri, P.; Soranzo, M.R. Enzymatic basis of the respiratory burst of guinea-pig resident peritoneal-macrophages. J. Reticuloendothel. Soc. 1981, 29, 47–60. [Google Scholar] [PubMed]









© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, J.; Wan, B.; Yang, Y.; Ren, X.-M.; Guo, L.-H.; Liu, J.-F. Biodegradation of Single-Walled Carbon Nanotubes in Macrophages through Respiratory Burst Modulation. Int. J. Mol. Sci. 2016, 17, 409. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030409
Hou J, Wan B, Yang Y, Ren X-M, Guo L-H, Liu J-F. Biodegradation of Single-Walled Carbon Nanotubes in Macrophages through Respiratory Burst Modulation. International Journal of Molecular Sciences. 2016; 17(3):409. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030409
Chicago/Turabian StyleHou, Jie, Bin Wan, Yu Yang, Xiao-Min Ren, Liang-Hong Guo, and Jing-Fu Liu. 2016. "Biodegradation of Single-Walled Carbon Nanotubes in Macrophages through Respiratory Burst Modulation" International Journal of Molecular Sciences 17, no. 3: 409. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17030409