
MicroRNA-15b Modulates Molecular Mediators of Blood Induced Arthropathy in Hemophilia Mice

Abstract
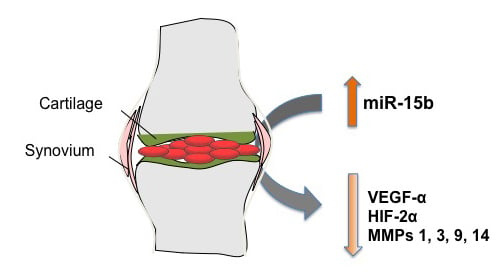
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. miR-15b Expression Is Altered during the Development of Arthropathy in Vivo
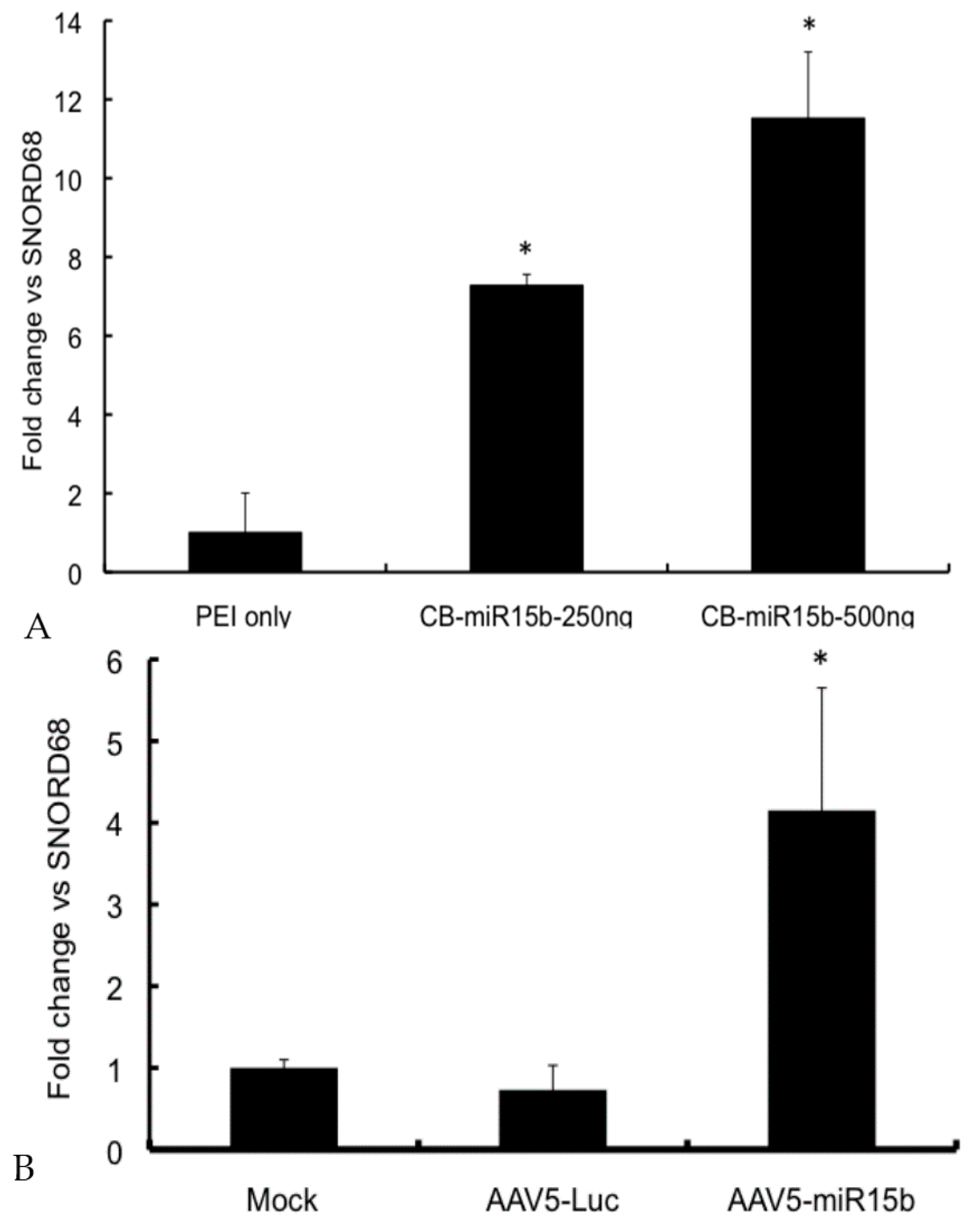
2.2. miR-15b Vectors Are Functional in Vitro
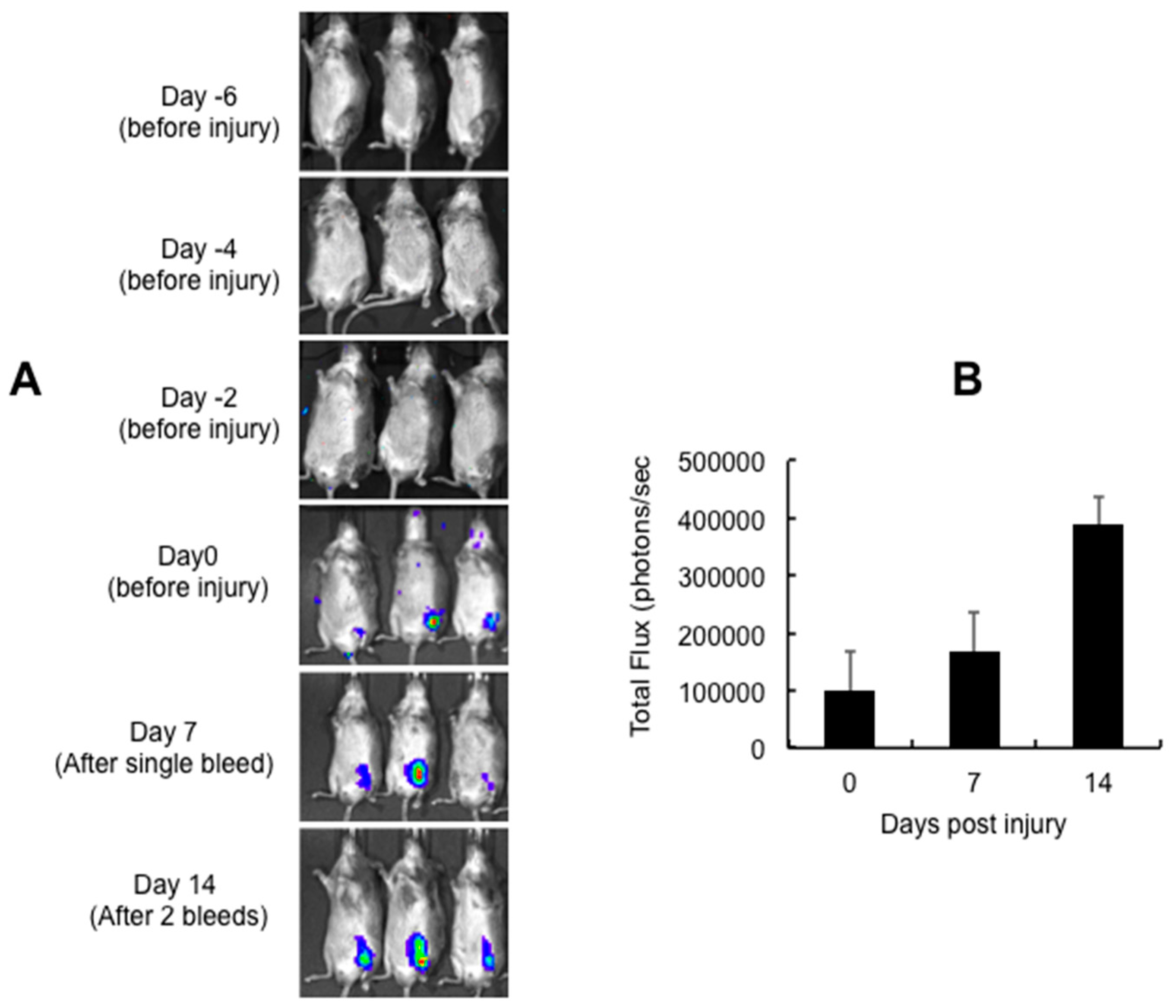
2.3. Kinetics of Adeno-Associated Virus (AAV) Mediated Transgene Expression in Joint Tissue of Hemarthrosis Model
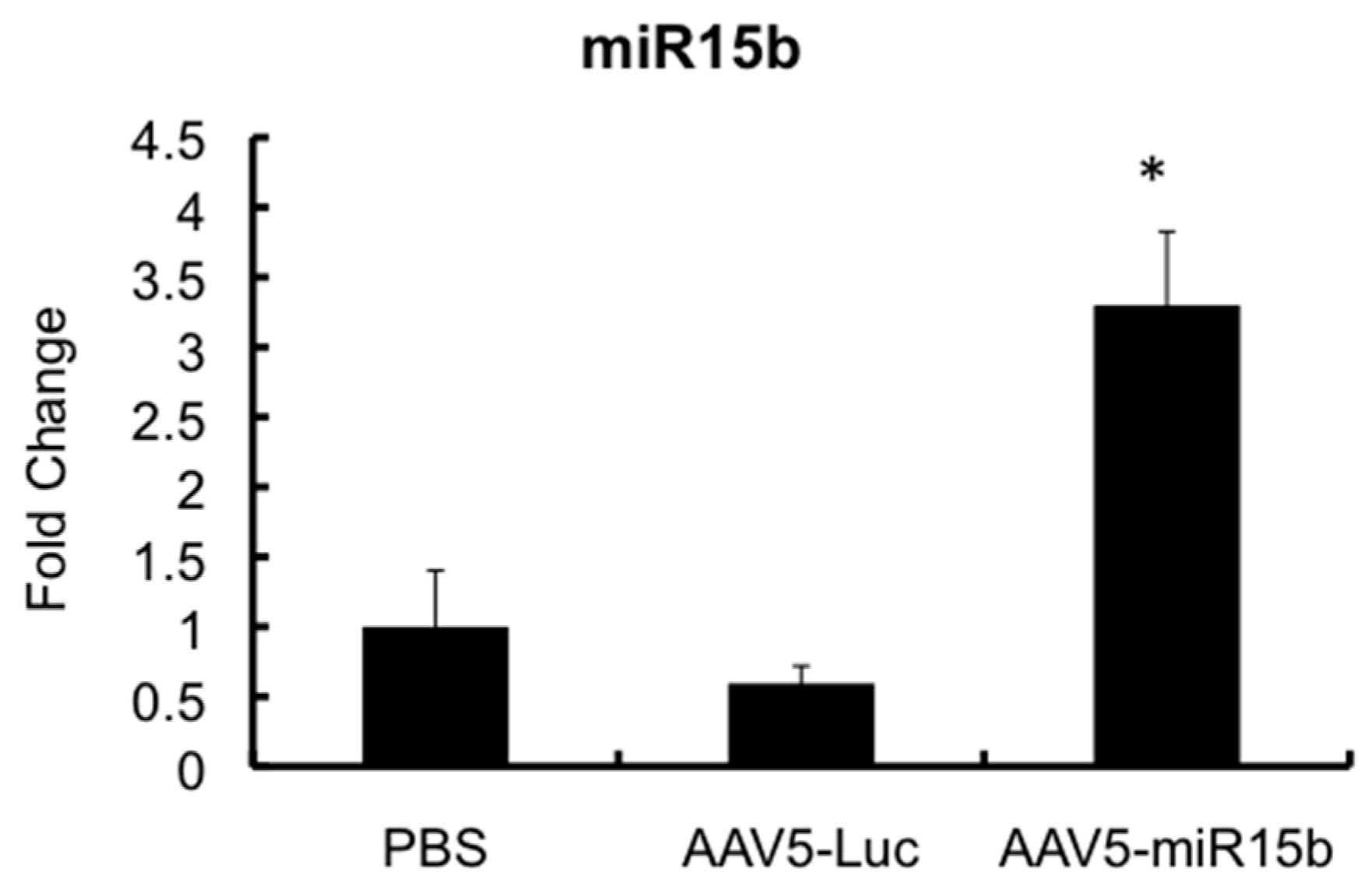
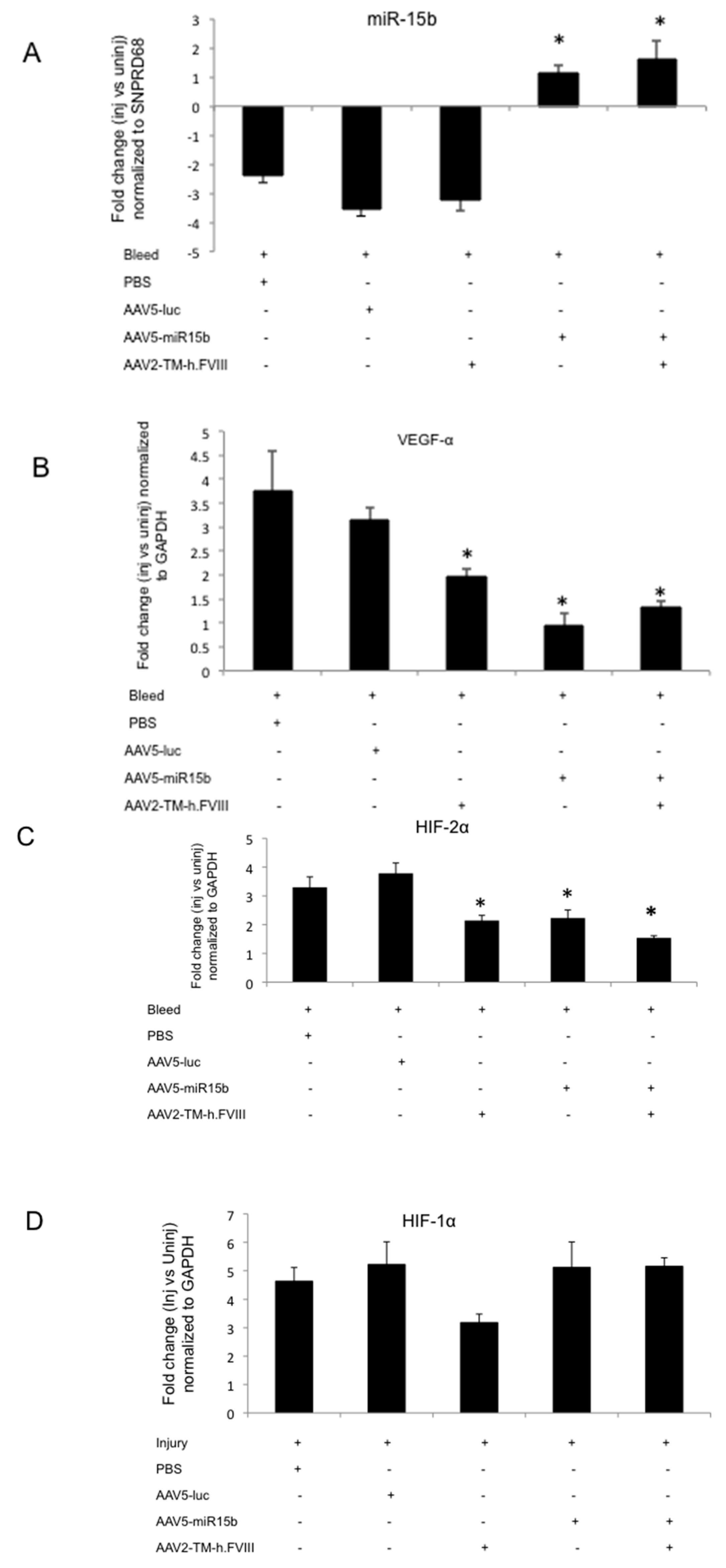
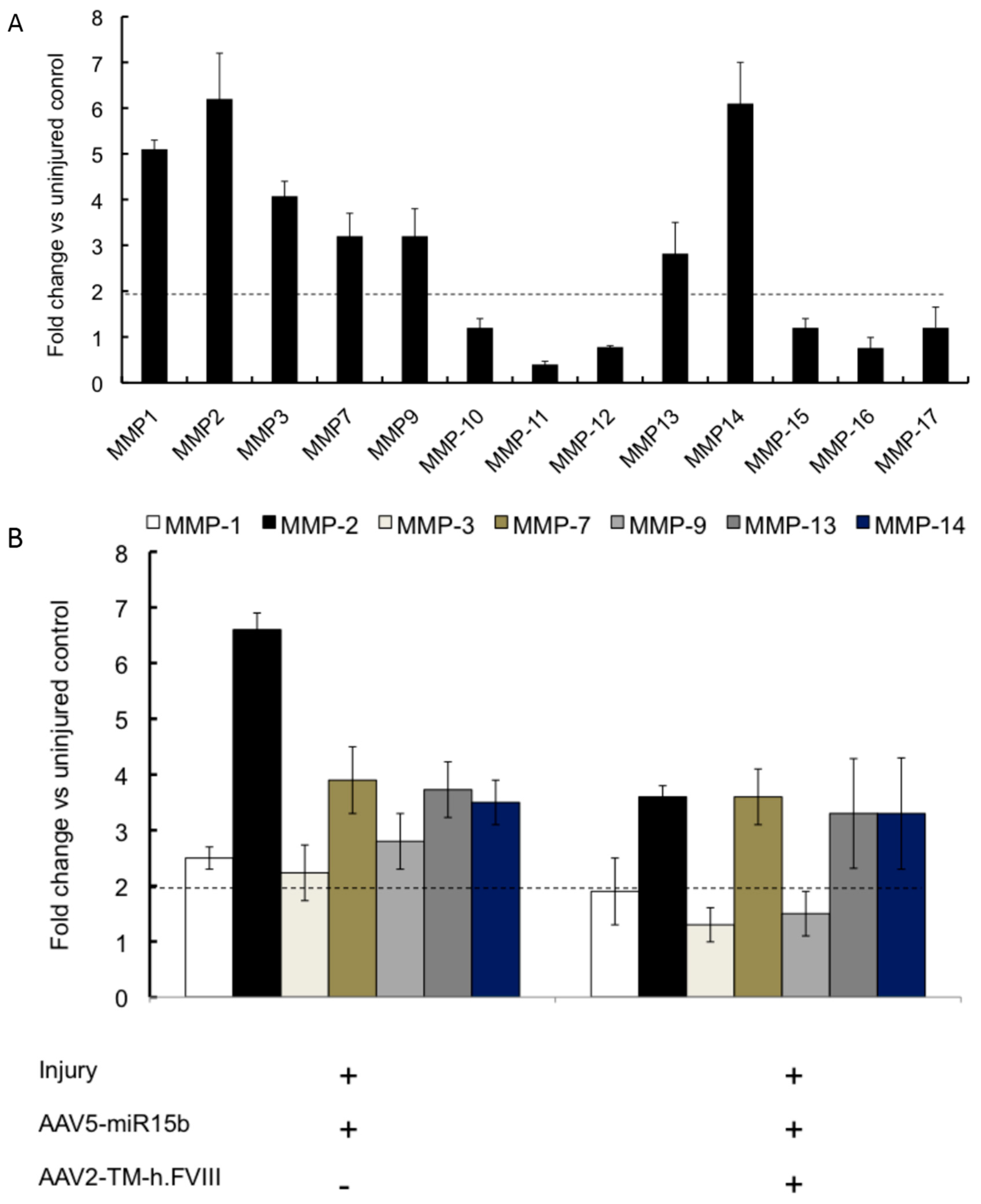
2.4. Intra-Articular Administration of AAV-miR-15b Attenuates Molecular Mediators of Arthropathy
3. Discussion
4. Experimental Section
4.1. Hemarthrosis Model
4.2. Vector Construction
4.3. Generation of AAV Vectors
4.4. Validation of miR-15b Vectors
4.5. In Vitro Studies
4.6. In Vivo Gene Delivery: Optimization of Intra-Articular Vector Delivery in Hemarthrosis Model
4.7. Systemic h.FVIII and Intra-Articular miR-15b Gene Transfer
4.8. Quantitative PCR
4.9. Immunoblotting
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Valentino, L.A.; Hakobyan, N.; Enockson, C.; Simpson, M.L.; Kakodkar, N.C.; Cong, L.; Song, X. Exploring the biological basis of haemophilic joint disease: Experimental studies. Haemophilia 2012, 18, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Roosendaal, G.; Vianen, M.E.; Marx, J.J.; van den Berg, H.M.; Lafeber, F.P.; Bijlsma, J.W. Blood-induced joint damage: A human in vitro study. Arthritis Rheum. 1999, 42, 1025–1032. [Google Scholar] [CrossRef]
- Stein, H.; Duthie, R.B. The pathogenesis of chronic haemophilic arthropathy. J. Bone Jt. Surg. Br. 1981, 63B, 601–609. [Google Scholar]
- Hooiveld, M.J.; Roosendaal, G.; Jacobs, K.M.; Vianen, M.E.; van den Berg, H.M.; Bijlsma, J.W.; Lafeber, F.P. Initiation of degenerative joint damage by experimental bleeding combined with loading of the joint: A possible mechanism of hemophilic arthropathy. Arthritis Rheum. 2004, 50, 2024–2031. [Google Scholar] [CrossRef] [PubMed]
- Handelsman, J.E. The knee joint in hemophilia. Orthop. Clin. N. Am. 1979, 10, 139–173. [Google Scholar]
- Van den Berg, H.M.; de Groot, P.H.; Fischer, K. Phenotypic heterogeneity in severe hemophilia. J. Thromb. Haemost. 2007, 5, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Sen, D.; Chapla, A.; Walter, N.; Daniel, V.; Srivastava, A.; Jayandharan, G.R. Nuclear factor (NF)-κB and its associated pathways are major molecular regulators of blood-induced joint damage in a murine model of hemophilia. J. Thromb. Haemost. 2013, 11, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Acharya, S.S.; Kaplan, R.N.; Macdonald, D.; Fabiyi, O.T.; DiMichele, D.; Lyden, D. Neoangiogenesis contributes to the development of hemophilic synovitis. Blood 2011, 117, 2484–2493. [Google Scholar] [CrossRef] [PubMed]
- Ovlisen, K.; Kristensen, A.T.; Jensen, A.L.; Tranholm, M. IL-1β, IL-6, KC and MCP-1 are elevated in synovial fluid from haemophilic mice with experimentally induced haemarthrosis. Haemophilia 2009, 15, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Medina, P.P.; Slack, F.J. microRNAs and cancer: An overview. Cell Cycle 2008, 7, 2485–2492. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.J.; Pan, Q.; Li, D.G.; Sun, H.; Liu, B.W. miR-15b and miR-16 are implicated in activation of the rat hepatic stellate cell: An essential role for apoptosis. J. Hepatol. 2009, 50, 766–778. [Google Scholar] [CrossRef] [PubMed]
- An, F.; Gong, B.; Wang, H.; Yu, D.; Zhao, G.; Lin, L.; Tang, W.; Yu, H.; Bao, S.; Xie, Q. miR-15b and miR-16 regulate TNF mediated hepatocyte apoptosis via BCL2 in acute liver failure. Apoptosis 2012, 17, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Lv, Q.; Ye, W.; Wong, C.K.; Cai, G.; Gu, D.; Ji, Y.; Zhao, C.; Wang, J.; Yang, B.B.; et al. miRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS ONE 2006, 1, e116. [Google Scholar] [CrossRef] [PubMed]
- Nicoloso, M.S.; Spizzo, R.; Shimizu, M.; Rossi, S.; Calin, G.A. MicroRNAs—The micro steering wheel of tumour metastases. Nat. Rev. Cancer 2009, 9, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, H.; Kimura, T.; Dalal, S.; Okada, Y.; D'Armiento, J. Joint diseases and matrix metalloproteinases: A role for MMP-13. Curr. Pharm. Biotechnol. 2008, 9, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Hakobyan, N.; Valentino, L.A.; Feldman, B.L.; Samulski, R.J.; Monahan, P.E. Intraarticular factor IX protein or gene replacement protects against development of hemophilic synovitis in the absence of circulating factor IX. Blood 2008, 112, 4532–4541. [Google Scholar] [CrossRef] [PubMed]
- Tas, S.W.; Adriaansen, J.; Hajji, N.; Bakker, A.C.; Firestein, G.S.; Vervoordeldonk, M.J.; Tak, P.P. Amelioration of arthritis by intraarticular dominant negative Ikkβ gene therapy using adeno-associated virus type 5. Hum. Gene Ther. 2006, 17, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Adriaansen, J.; Khoury, M.; de Cortie, C.J.; Fallaux, F.J.; Bigey, P.; Scherman, D.; Gould, D.J.; Chernajovsky, Y.; Apparailly, F.; Jorgensen, C.; et al. Reduction of arthritis following intra-articular administration of an adeno-associated virus serotype 5 expressing a disease-inducible TNF-blocking agent. Ann. Rheum. Dis. 2007, 66, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Adriaansen, J.; Tas, S.W.; Klarenbeek, P.L.; Bakker, A.C.; Apparailly, F.; Firestein, G.S.; Jorgensen, C.; Vervoordeldonk, M.J.; Tak, P.P. Enhanced gene transfer to arthritic joints using adeno-associated virus type 5: Implications for intra-articular gene therapy. Ann. Rheum. Dis. 2005, 64, 1677–1684. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chopp, M.; Lu, Y.; Buller, B.; Jiang, F. miR-15b and miR-152 reduce glioma cell invasion and angiogenesis via NRP-2 and MMP-3. Cancer Lett. 2013, 329, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Houseman, M.; Potter, C.; Marshall, N.; Lakey, R.; Cawston, T.; Griffiths, I.; Young-Min, S.; Isaacs, J.D. Baseline serum MMP-3 levels in patients with Rheumatoid Arthritis are still independently predictive of radiographic progression in a longitudinal observational cohort at 8 years follow up. Arthritis Res. Ther. 2012, 14, R30. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Choi, H.M.; Lee, Y.A.; Choi, I.A.; Lee, S.H.; Hong, S.J.; Yang, H.I.; Yoo, M.C. Expression levels and association of gelatinases MMP-2 and MMP-9 and collagenases MMP-1 and MMP-13 with VEGF in synovial fluid of patients with arthritis. Rheumatol. Int. 2011, 31, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.R.; Schoenfeld, L.W.; Ruby, J.G.; Auyeung, V.C.; Spies, N.; Baek, D.; Johnston, W.K.; Russ, C.; Luo, S.; Babiarz, J.E.; et al. Mammalian microRNAs: Experimental evaluation of novel and previously annotated genes. Genes Dev. 2010, 24, 992–1009. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Wei, T.; Janson, P.C.; Saaf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A.; et al. MicroRNAs: Novel regulators involved in the pathogenesis of psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef] [PubMed]
- Stanczyk, J.; Pedrioli, D.M.; Brentano, F.; Sanchez-Pernaute, O.; Kolling, C.; Gay, R.E.; Detmar, M.; Gay, S.; Kyburz, D. Altered expression of microRNA in synovial fibroblasts and synovial tissue in rheumatoid arthritis. Arthritis Rheum. 2008, 58, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Malizos, K.N.; Oikonomou, P.; Tsezou, A. Integrative microRNA and proteomic approaches identify novel osteoarthritis genes and their collaborative metabolic and inflammatory networks. PLoS ONE 2008, 3, e3740. [Google Scholar] [CrossRef] [PubMed]
- Padgett, K.A.; Lan, R.Y.; Leung, P.C.; Lleo, A.; Dawson, K.; Pfeiff, J.; Mao, T.K.; Coppel, R.L.; Ansari, A.A.; Gershwin, M.E. Primary biliary cirrhosis is associated with altered hepatic microRNA expression. J. Autoimmun. 2009, 32, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Huang, Y.S.; Tang, M.; Lv, T.Y.; Hu, C.X.; Tan, Y.H.; Xu, Z.M.; Yin, Y.B. Microarray analysis of microRNA expression in peripheral blood cells of systemic lupus erythematosus patients. Lupus 2007, 16, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zikusoka, M.; Trindade, A.; Dassopoulos, T.; Harris, M.L.; Bayless, T.M.; Brant, S.R.; Chakravarti, S.; Kwon, J.H. MicroRNAs are differentially expressed in ulcerative colitis and alter expression of macrophage inflammatory peptide-2α. Gastroenterology 2008, 135, 1624–1635. [Google Scholar] [CrossRef] [PubMed]
- Kulshreshtha, R.; Davuluri, R.V.; Calin, G.A.; Ivan, M. A microRNA component of the hypoxic response. Cell Death Differ. 2008, 15, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Augustsson, J.; Neovius, M.; Cullinane-Carli, C.; Eksborg, S.; van Vollenhoven, R.F. Patients with rheumatoid arthritis treated with tumour necrosis factor antagonists increase their participation in the workforce: Potential for significant long-term indirect cost gains (data from a population-based registry). Ann. Rheum. Dis. 2010, 69, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Abba, M.; Patil, N.; Allgayer, H. MicroRNAs in the Regulation of MMPs and Metastasis. Cancers (Basel) 2014, 6, 625–645. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Meng, X.; Han, J.; Zhang, Z.; Wang, B.; Bai, X.; Zhang, X. Anti-cancer activity of DHA on gastric cancer—An in vitro and in vivo study. Tumour Biol. 2013, 34, 3791–3800. [Google Scholar] [CrossRef] [PubMed]
- Heishima, K.; Mori, T.; Ichikawa, Y.; Sakai, H.; Kuranaga, Y.; Nakagawa, T.; Tanaka, Y.; Okamura, Y.; Masuzawa, M.; Sugito, N.; et al. MicroRNA-214 and microRNA-126 are potential biomarkers for malignant endothelial proliferative diseases. Int. J. Mol. Sci. 2015, 16, 25377–25391. [Google Scholar] [CrossRef] [PubMed]
- Hanin, G.; Shenhar-Tsarfaty, S.; Yayon, N.; Yau, Y.H.; Bennett, E.R.; Sklan, E.H.; Rao, D.C.; Rankinen, T.; Bouchard, C.; Geifman-Shochat, S.; et al. Competing targets of microRNA-608 affect anxiety and hypertension. Hum. Mol. Genet. 2014, 23, 4569–4580. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.; Lenting, P.J.; Rosales, C.; Lee, D.; Rabbanian, S.; Raj, D.; Patel, N.; Tuddenham, E.G.; Christophe, O.D.; McVey, J.H.; et al. Therapeutic levels of FVIII following a single peripheral vein administration of rAAV vector encoding a novel human factor VIII variant. Blood 2013, 121, 3335–3344. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, N.; Hareendran, S.; Sen, D.; Gadkari, R.A.; Sudha, G.; Selot, R.; Hussain, M.; Dhaksnamoorthy, R.; Samuel, R.; Srinivasan, N.; et al. Bioengineering of AAV2 capsid at specific serine, threonine, or lysine residues improves its transduction efficiency in vitro and in vivo. Hum. Gene Ther. Methods 2013, 24, 80–93. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sen, D.; Jayandharan, G.R. MicroRNA-15b Modulates Molecular Mediators of Blood Induced Arthropathy in Hemophilia Mice. Int. J. Mol. Sci. 2016, 17, 492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17040492
Sen D, Jayandharan GR. MicroRNA-15b Modulates Molecular Mediators of Blood Induced Arthropathy in Hemophilia Mice. International Journal of Molecular Sciences. 2016; 17(4):492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17040492
Chicago/Turabian StyleSen, Dwaipayan, and Giridhara R. Jayandharan. 2016. "MicroRNA-15b Modulates Molecular Mediators of Blood Induced Arthropathy in Hemophilia Mice" International Journal of Molecular Sciences 17, no. 4: 492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17040492