
Molecular Signals Controlling the Inhibition of Nodulation by Nitrate in Medicago truncatula

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
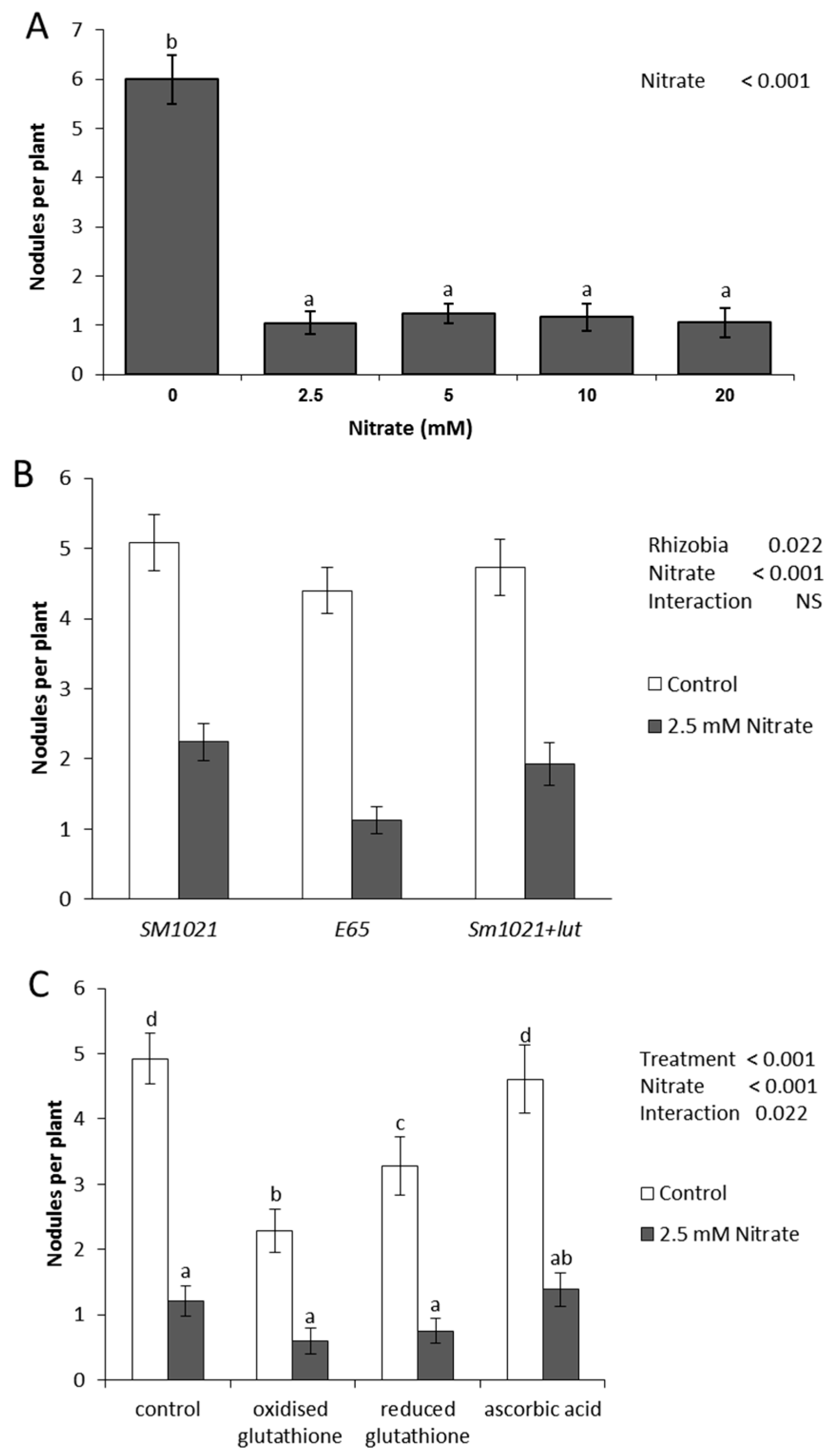
2.1. Nodulation in the Presence of Nitrate
2.2. Proteome Analysis
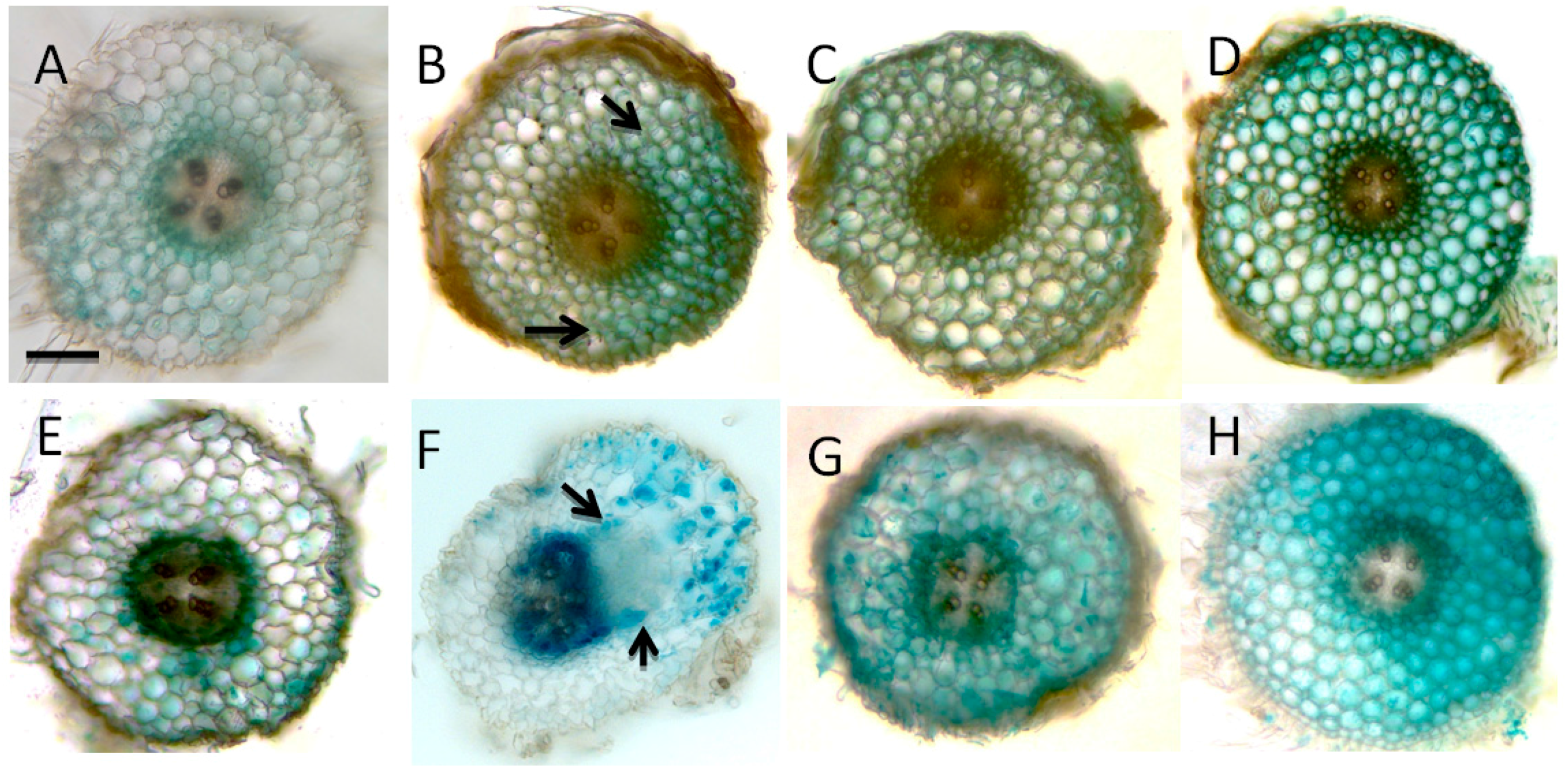
2.3. Involvement of Flavonoids in Nitrate Inhibition of Nodulation
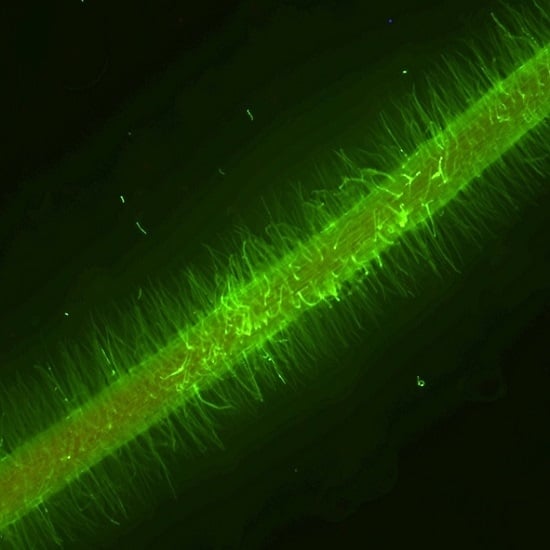
2.4. Involvement of Reactive Oxygen Species in Nitrate Inhibition of Nodulation
3. Discussion
4. Materials and Methods
4.1. Plant Growth
4.2. Microscopy
4.3. Proteome Analysis
4.4. Histochemical GUS Staining
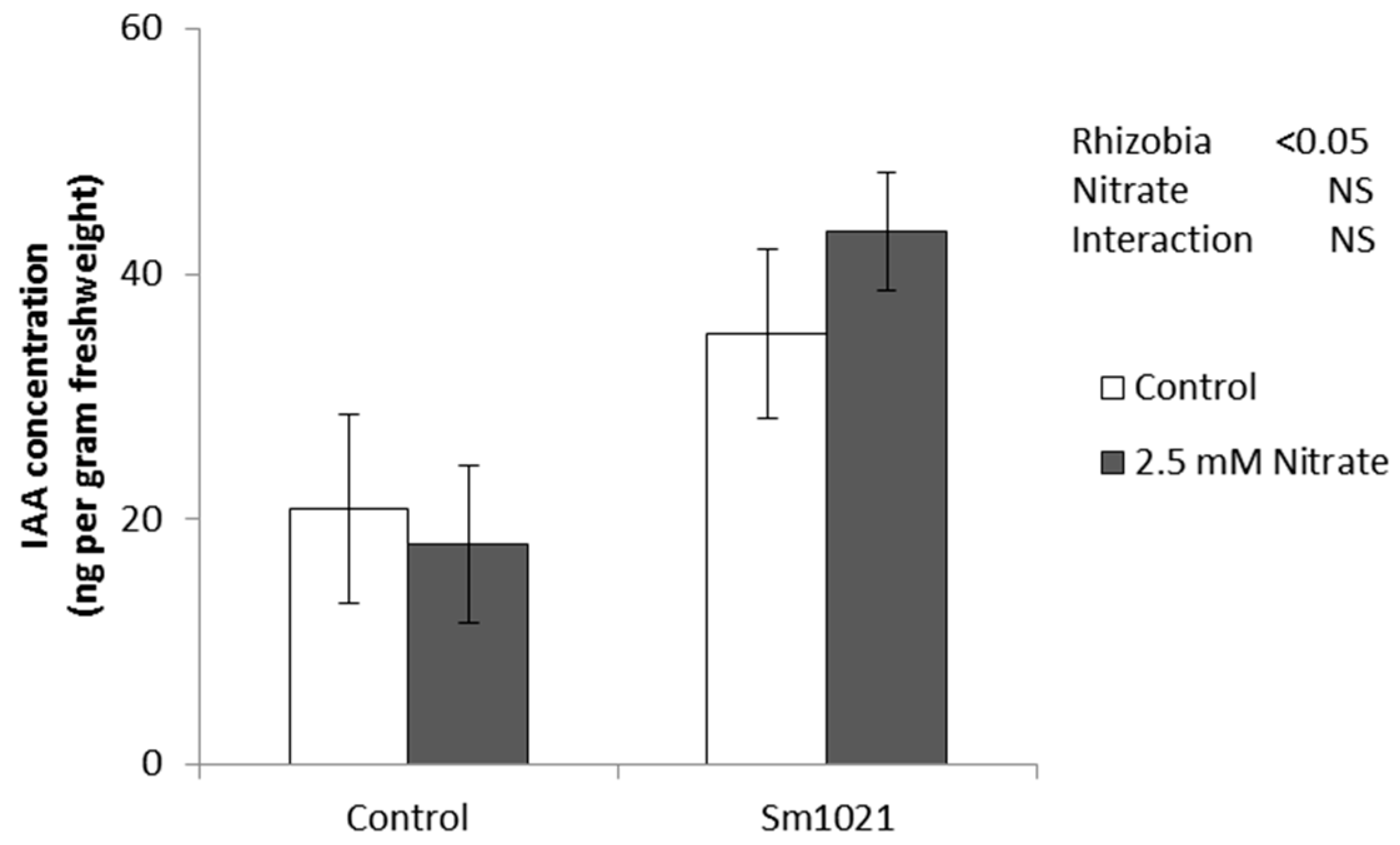
4.5. Quantification of Auxin
4.6. Staining of Reactive Oxygen Species
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oldroyd, G.E.D.; Downie, J.M. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Mortier, V.; Holsters, M.; Goormachtig, S. Never too many? How legumes control nodule numbers. Plant Cell Environ. 2012, 35, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.E.; Ferguson, B.J.; Hayashi, S.; Lin, Y.-H.; Gresshoff, P.M. Molecular mechanisms controlling legume autoregulation of nodulation. Ann. Bot. 2011, 108, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Streeter, J. Inhibition of legume nodule formation and N2 fixation by nitrate. Crit. Rev. Plant Sci. 1988, 7, 1–23. [Google Scholar] [CrossRef]
- Dakora, F.D. Nodule function in symbiotic bambara groundnut (Vigna subterranea L.) and Kersting’s bean (Macrotyloma geocarpum L.) is tolerant of nitrate in the root medium. Ann. Bot. 1998, 82, 687–690. [Google Scholar] [CrossRef]
- Bollman, M.I.; Vessey, J.K. Differential effects of nitrate and ammonium supply on nodule initiation, development, and distribution on roots of pea (Pisum sativum). Can. J. Bot. 2006, 84, 893–903. [Google Scholar] [CrossRef]
- Coronado, C.; Zuanazzi, J.A.S.; Sallaud, C.; Quirion, J.C.; Esnault, R.; Husson, H.P.; Kondorosi, A.; Ratet, P. Medicago sativa root flavonoid production is nitrogen regulated. Plant Physiol. 1995, 108, 533–542. [Google Scholar] [PubMed]
- Barbulova, A.; Rogato, A.; D’Apuzzo, E.; Omrane, S.; Chiurazzi, M. Differential effects of combined N sources on early steps of the Nod factor-dependent transduction pathway in Lotus japonicus. Mol. Plant-Microbe Interact. 2007, 20, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Supanjani, S.; Lee, K.D.; Almaraz, J.J.; Zhou, X.; Smith, D.L. Effect of organic N source on bacterial growth, lipo-chitooligosaccharide production, and early soybean nodulation by Bradyrhizobium japonicum. Can. J. Microbiol. 2006, 52, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Jeudy, C.; Ruffel, S.; Freixes, S.; Tillard, P.; Santoni, A.L.; Morel, S.; Journet, E.-P.; Duc, G.; Gojon, A.; Lepetit, M. Adaptation of Medicago truncatula to nitrogen limitation is modulated via local and systemic nodule developmental responses. New Phytol. 2010, 185, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Day, D.A.; Caroll, B.J.; Delves, A.C.; Gresshoff, P.M. Relationship between autoregulation anf nitrate inhibition in soybeans. Physiol. Plant. 1989, 75, 37–42. [Google Scholar] [CrossRef]
- Okamoto, S.; Ohnishi, E.; Sato, S.; Takahashi, H.; Nakazono, M.; Tabata, S.; Kawaguchi, M. Nod factor/nitrate-induced CLE genes that drive HAR1-mediated systemic regulation of nodulation. Plant Cell Physiol. 2009, 50, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.E.; Ferguson, B.J.; Gresshoff, P.M. Inoculation- and nitrate-induced CLE peptides of soybean control NARK-dependent nodule formation. Mol. Plant-Microbe Interact. 2011, 24, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Caba, J.M.; Recalde, L.; Ligero, F. Nitrate-induced ethylene biosynthesis and the control of nodulation in alfalfa. Plant Cell Environ. 1998, 21, 87–93. [Google Scholar] [CrossRef]
- Ligero, F.; Caba, J.M.; Lluch, C.; Olivares, J. Nitrate inhibition of nodulation can be overcome by the ethylene inhibitor aminoethoxyvinylglycine. Plant Physiol. 1991, 97, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Engstrom, E.M.; Long, S.R. Ethylene inhibits the Nod factor signal transduction pathway of Medicago truncatula. Plant Cell. 2001, 13, 1835–1849. [Google Scholar] [CrossRef] [PubMed]
- Penmetsa, R.V.; Cook, D.R. A legume ethylene-insensitive mutant hyperinfected by its rhizobial symbiont. Science 1997, 275, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Caba, J.M.; Centeno, M.L.; Fernandez, B.; Gresshoff, P.M.; Ligero, F. Inoculation and nitrate alter phytohormone levels in soybean roots: Differences between a supernodulating mutant and the wild type. Planta 2000, 211, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Fei, H.; Vessey, J.K. Involvement of cytokinin in the stimulation of nodulation by low concentrations of ammonium in Pisum sativum. Physiol. Plant. 2003, 118, 447–455. [Google Scholar] [CrossRef]
- Mathesius, U.; Charon, C.; Rolfe, B.G.; Kondorosi, A.; Crespi, M. Temporal and spatial order of events during the induction of cortical cell divisions in white clover by Rhizobium leguminosarum bv. trifolii inoculation or localized cytokinin addition. Mol. Plant-Microbe Interact. 2000, 13, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rizzo, S.; Crespi, M.; Frugier, F. The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell. 2006, 18, 2680–2693. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.D.; Karas, B.J.; Sato, S.; Tabata, S.; Amyot, L.; Szczyglowski, K. A cytokinin perception mutant colonized by Rhizobium in the absence of nodule organogenesis. Science 2007, 315, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Walch-Liu, P.; Filleur, S.; Gan, Y.B.; Forde, B.G. Signaling mechanisms integrating root and shoot responses to changes in the nitrogen supply. Photosyn. Res. 2005, 83, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Jennings, A.; Barlow, P.W.; Forde, B.G. Dual pathways for regulation of root branching by nitrate. Proc. Natl. Acad. Sci. USA 1999, 96, 6529–6534. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Watt, M.; Mathesius, U. The autoregulation gene SUNN mediates changes in root organ formation in response to nitrogen through alteration of shoot-to-root auxin transport. Plant Physiol. 2012, 159, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Fujikake, H.; Yamazaki, A.; Ohtake, N.; Sueyoshi, K.; Matsuhashi, S.; Ito, T.; Mizuniwa, C.; Kume, T.; Hashimoto, S.; Ishioka, N. Quick and reversible inhibition of soybean root nodule growth by nitrate involves a decrease in sucrose supply to nodules. J. Exp. Bot. 2003, 54, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Mathesius, U.; Keijzers, G.; Natera, S.H.A.; Weinman, J.J.; Djordjevic, M.A.; Rolfe, B.G. Establishment of a root proteome reference map for the model legume Medicago truncatula using the expressed sequence tag database for peptide mass fingerprinting. Proteomics 2001, 1, 1424–1440. [Google Scholar] [CrossRef]
- Lei, Z.T.; Elmer, A.M.; Watson, B.S.; Dixon, R.A.; Mendes, P.J.; Sumner, L.W. A two-dimensional electrophoresis proteomic reference map and systematic identification of 1367 proteins from a cell suspension culture of the model legume Medicago truncatula. Molec. Cell. Proteom. 2005, 4, 1812–1825. [Google Scholar] [CrossRef] [PubMed]
- Bestel-Corre, G.; Dumas-Gaudot, E.; Poinsot, V.; Dieu, M.; Dierick, J.-F.; Tuinen, D.v.; Remacle, J.; Gianinazzi-Pearson, V.; Gianinazzi, S. Proteome analysis and identification of symbiosis-related proteins from Medicago truncatula Gaertn. by two-dimensional electrophoresis and mass spectrometry. Electrophoresis 2002, 23, 122–137. [Google Scholar] [CrossRef]
- Larrainzar, E.; Wienkoop, S.; Weckwerth, W.; Ladrera, R.; Arrese-Igor, C.; Gonzalez, E.M. Medicago truncatula root nodule proteome analysis reveals differential plant and bacteroid responses to drought stress. Plant Physiol. 2007, 144, 1495–1507. [Google Scholar] [CrossRef] [PubMed]
- Prayitno, J.; Imin, N.; Rolfe, B.G.; Mathesius, U. Identification of ethylene-mediated protein changes during nodulation in Medicago truncatula using proteome analysis. J. Proteome Res. 2006, 5, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Van Noorden, G.E.; Kerim, T.; Goffard, N.; Wiblin, R.; Pellerone, F.I.; Rolfe, B.G.; Mathesius, U. Overlap of proteome changes in Medicago truncatula in response to auxin and Sinorhizobium meliloti. Plant Physiol. 2007, 144, 1115–1131. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.; Harper, P. Effect of localized nitrate application on isoflavonoid concentration and nodulation in split-root systems of wild-type and nodulation-mutant soybean plants. Plant Physiol. 1991, 95, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Peters, N.K.; Frost, J.W.; Long, S.R. The flavone, luteolin, induces expression of Rhizobium meliloti nodulation genes. Science 1986, 233, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Barnett, M.J.; Tolman, C.J.; Fisher, R.F.; Long, S.R. A dual-genome Symbiosis Chip for coordinate study of signal exchange and development in a prokaryote-host interaction. Proc. Natl. Acad. Sci. USA 2004, 101, 16636–16641. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.L.P.; Hassan, S.; Truong, T.T.; Hocart, C.H.; Laffont, C.; Frugier, F.; Mathesius, U. Flavonoids and auxin transport inhibitors rescue symbiotic nodulation in the Medicago truncatula cytokinin perception mutant cre1. Plant Cell 2015, 27, 2210–2226. [Google Scholar] [CrossRef] [PubMed]
- Pauly, N.; Pucciariello, C.; Mandon, K.; Innocenti, G.; Jamet, A.; Boudouin, E.; Herouart, D.; Frendo, P.; Puppo, A. Reactive oxygen and nitrogen species and glutathione: Key players in the legume-Rhizobium symbiosis. J. Exp. Bot. 2006, 57, 1769–1776. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, K.A.; Jensen, P.E.; Moller, I.M.; Schulz, A. Monitoring reactive oxygen species formation and localisation in living cells by use of the fluorescent probe CM-H2DCFDA and confocal laser microscopy. Physiol. Plant. 2009, 136, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; MArquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: an integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef] [PubMed]
- Carroll, B.J.; Gresshoff, P.M. Nitrate inhibition of nodulation and nitrogen-fixation in white clover. Z. Pflanzenphysiol. 1983, 110, 77–88. [Google Scholar] [CrossRef]
- Carroll, B.J.; McNeil, D.L.; Gresshoff, P.M. Isolation and properties of soybean Glycine max (L) Merr mutants that nodulate in the presence of high nitrate concentrations. Proc. Natl. Acad. Sci. USA 1985, 82, 4162–4166. [Google Scholar] [CrossRef] [PubMed]
- Lohar, D.; Sharopova, N.; Endre, G.; Panuela, S.; Samac, D.; Town, C.; Silverstein, K.; Vanden Bosch, K. Transcript analysis of early nodulation events in Medicago truncatula. Plant Physiol. 2006, 140, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Bouguyon, E.; Gojon, A.; Nacry, P. Nitrate sensing and signaling in plants. Semin. Cell Dev. Biol. 2012, 23, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.-H.; Nicotra, A.B.; Mathesius, U. The presence of nodules on legume root systems can alter phenotypic plasticity in response to internal nitrogen independent of nitrogen fixation. Plant Cell Environ. 2016, 39, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Omrane, S.; Ferrarini, A.; D’Apuzzo, E.; Rogato, A.; Delledonne, M.; Chiurazzi, M. Symbiotic competence in Lotus japonicus is affected by plant nitrogen status: Transcriptomic identification of genes affected by a new signalling pathway. New Phytol. 2009, 183, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Ruffel, S.; Freixes, S.; Balzergue, S.; Tillard, P.; Jeudy, C.; Martin-Magniette, M.L.; van Der Merwe, M.J.; Kakar, K.; Gouzy, J.; Fernie, A.R. Systemic signaling of the plant nitrogen status triggers specific transcriptome responses depending on the nitrogen source in Medicago truncatula. Plant Physiol. 2008, 146, 2020–2035. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.-M.; Koschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation in amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef] [PubMed]
- Borsics, T.; Lados, M. Dodder infection induces the expression of a pathogenesis-related gene of the family PR-10 in alfalfa. J. Exp. Bot. 2002, 53, 1831–1832. [Google Scholar] [CrossRef] [PubMed]
- Colditz, F.; Nyamsuren, O.; Niehaus, K.; Eubel, H.; Braun, H.; Krajinski, F. Proteomic approach: Identification of Medicago truncatula proteins induced in roots after infection with the pathogenic oomycete Aphanomyces euteiches. Plant Mol. Biol. 2004, 55, 109–120. [Google Scholar] [CrossRef] [PubMed]
- De Jong, F.; Mathesius, U.; Imin, N.; Rolfe, B.G. A proteome study of the proliferation of cultured Medicago truncatula protoplasts. Proteomics 2007, 7, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Holmes, P.; Farquharson, R.; Hall, P.J.; Rolfe, B.G. Proteomic analysis of root meristems and the effects of acetohydroxyacid synthase-inhibiting herbicides in the root of Medicago truncatula. J. Proteome Res. 2006, 5, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Imin, N.; Nizamidin, M.; Daniher, D.; Nolan, K.E.; Rose, R.J.; Rolfe, B.G. Proteomic analysis of somatic embryogenesis in Medicago truncatula. Explant cultures grown under 6-benzylaminopurine and 1-naphthaleneacetic acid treatments. Plant Physiol. 2005, 137, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, J.E.; Wimmer, R.; Larsen, J.N.; Spangfort, M.D.; Otzen, D.E. The major birch allergen, Bet v 1, shows affinity for a broad spectrum of physiological ligands. J. Biol. Chem. 2002, 277, 23684–23692. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.J.; Sturgess, F.J.; Farrell, G.G.; Draper, J. The AoPR10 promoter and certain endogenous PR10 genes respond to oxidative signals in Arabidopsis. Mol. Plant Pathol. 2004, 5, 435–451. [Google Scholar] [CrossRef] [PubMed]
- Ramu, S.K.; Peng, H.-M.; Cook, D.R. Nod factor induction of reactive oxygen species production is correlated with expression of the early nodulin gene rip1 in Medicago truncatula. Mol. Plant-Microbe Interact. 2002, 15, 552–528. [Google Scholar] [CrossRef] [PubMed]
- Peleg-Grossman, S.; Volpin, H.; Levine, A. Root hair curling and Rhizobium infection in Medicago truncatula are mediated by phosphatidylinositide-regulated endocytosis and reactive oxygen species. J. Exp. Bot. 2007, 58, 1637–1649. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, M.A.; Oakes, M.; Li, D.X.; Hwang, C.H.; Hocart, C.H.; Gresshoff, P.M. The Glycine max xylem sap and apoplast proteome. J. Proteome Res. 2007, 6, 3771–3779. [Google Scholar] [CrossRef] [PubMed]
- Park, D.-S.; Graham, M.Y.; Graham, T.L. Identification of soybean elicitation competency factor, CF-1, as the soybean Kunitz trypsin inhibitor. Physiol. Mol. Plant Pathol. 2001, 59, 265–273. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Broun, P. Transcriptional control of flavonoid biosynthesis: a complex network of conserved regulators involved in multiple aspects of differentiation in Arabidopsis. Curr. Opin. Plant Biol. 2005, 8, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, N.A.; Glover, B.J. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005, 10, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Wasson, A.P.; Pellerone, F.I.; Mathesius, U. Silencing the flavonoid pathway in Medicago truncatula inhibits root nodule formation and prevents auxin transport regulation by rhizobia. Plant Cell. 2006, 18, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Valera, C.L.; Alexander, M. Reversal of nitrate inhibition of nodulation by indol-3-acetic acid. Nature 1965, 206, 326. [Google Scholar] [CrossRef]
- Fahraeus, G. The infection of clover root hairs by nodule bacteria studied by a simple glass slide technique. J. Gen. Microbiol. 1957, 16, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Medicago truncatula gene index. Available online: http://compbio.dfci.harvard.edu/tgi/cgi-bin/tgi/gimain.pl?gudb=medicago (accessed on 3 May 2013).
- Medicago Gene Expression Atlas. Available online: http://mtgea.noble.org/v3/ (accessed on 23 February 2016).
- Bevan, M.; Bancroft, I.; Bent, E.; Love, K.; Goodman, H.; Dean, C.; Bergkamp, R.; Dirkse, W.; van Staveren, M.; Stiekema, W. Analysis of 1.9 Mb of contiguous sequence from chromosome 4 of Arabidopsis thaliana. Nature 1998, 391, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Kyoto Encyclopedia of Genes and Genomes. Available online: http://www.genome.jp/kegg/ (accessed on 23 February 2016).
- ExPASy SIB Bioinformatics Resource Portal. Available online: http://au.expasy.org/sprot/ (accessed on 23 February 2016).





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Noorden, G.E.; Verbeek, R.; Dinh, Q.D.; Jin, J.; Green, A.; Ng, J.L.P.; Mathesius, U. Molecular Signals Controlling the Inhibition of Nodulation by Nitrate in Medicago truncatula. Int. J. Mol. Sci. 2016, 17, 1060. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071060
Van Noorden GE, Verbeek R, Dinh QD, Jin J, Green A, Ng JLP, Mathesius U. Molecular Signals Controlling the Inhibition of Nodulation by Nitrate in Medicago truncatula. International Journal of Molecular Sciences. 2016; 17(7):1060. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071060
Chicago/Turabian StyleVan Noorden, Giel E., Rob Verbeek, Quy Dung Dinh, Jian Jin, Alexandra Green, Jason Liang Pin Ng, and Ulrike Mathesius. 2016. "Molecular Signals Controlling the Inhibition of Nodulation by Nitrate in Medicago truncatula" International Journal of Molecular Sciences 17, no. 7: 1060. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071060