
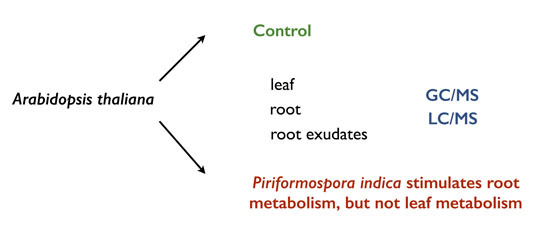
Piriformospora indica Stimulates Root Metabolism of Arabidopsis thaliana

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
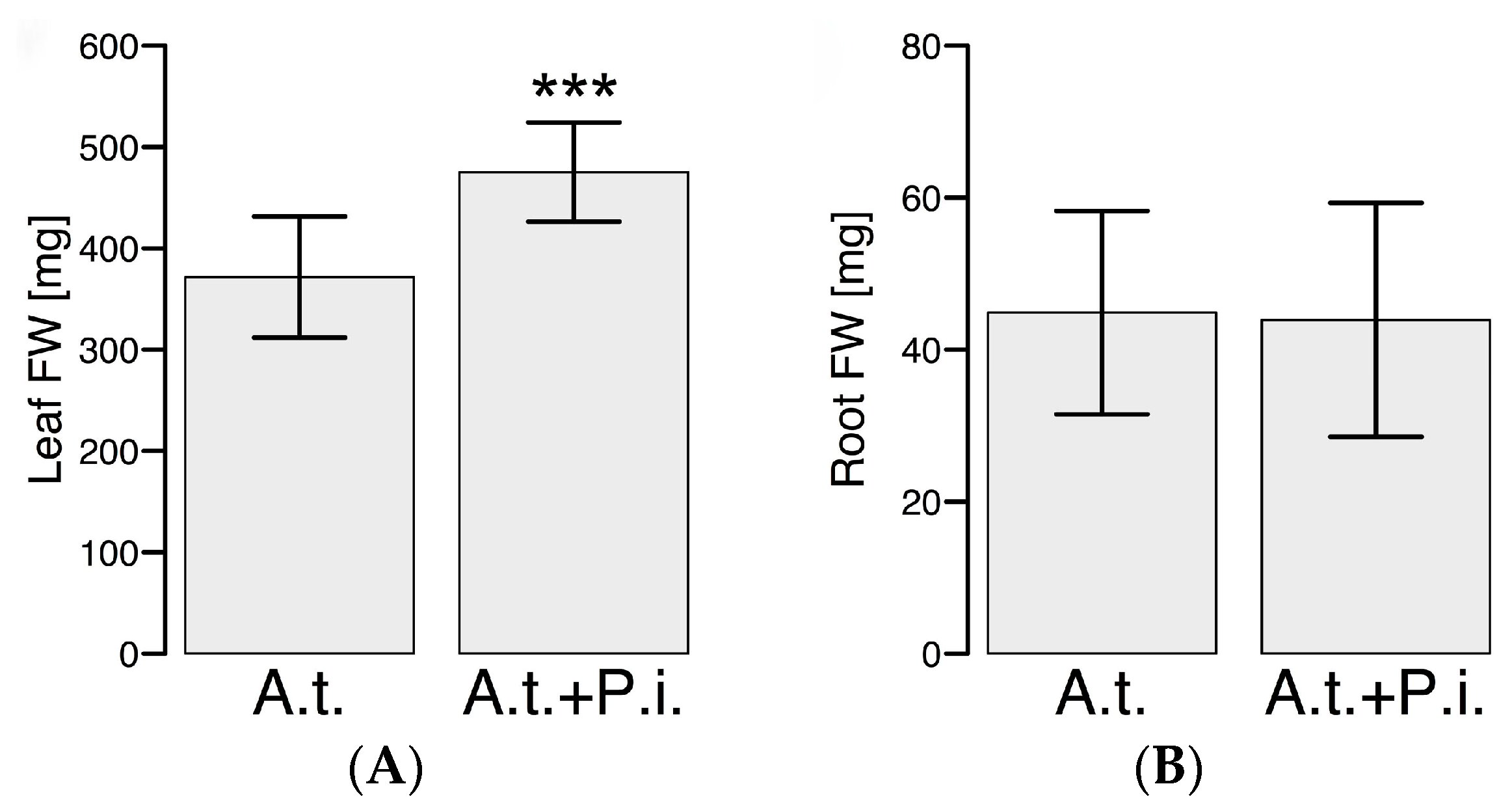
2.1. P. indica Promotes Shoot Growth of A. thaliana under Specific Culturing Conditions in a Hydroponic System after Root Colonization
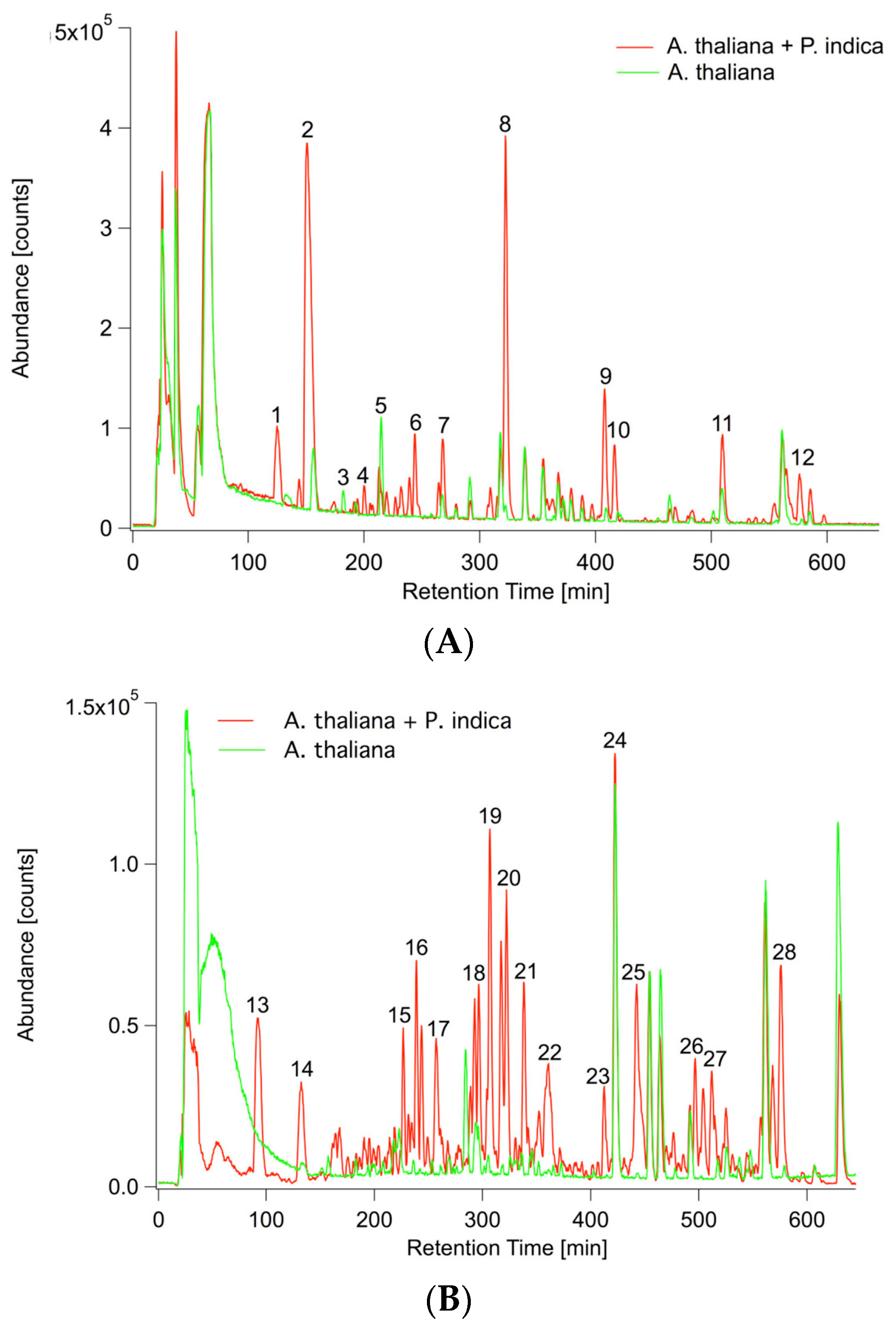
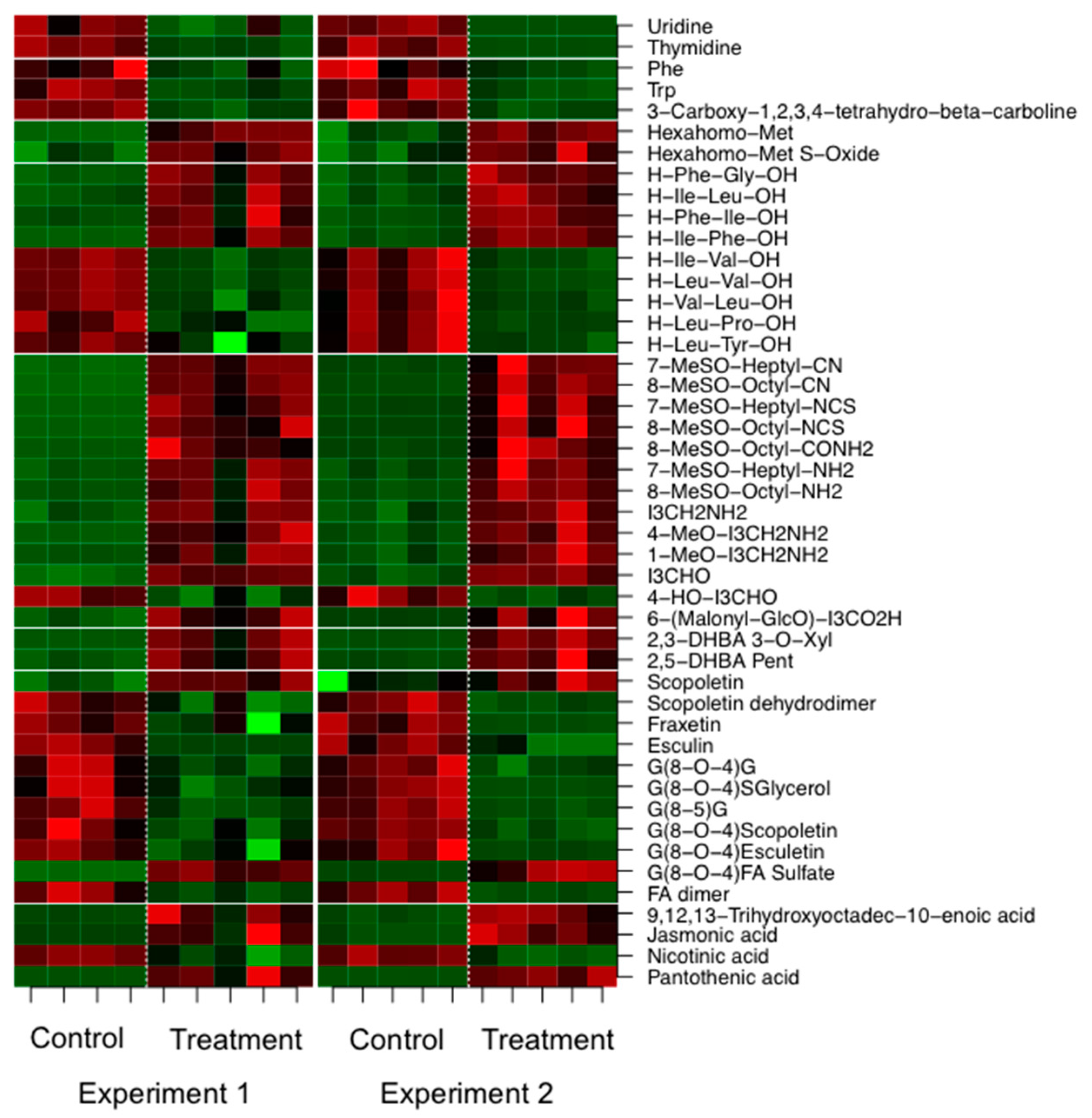
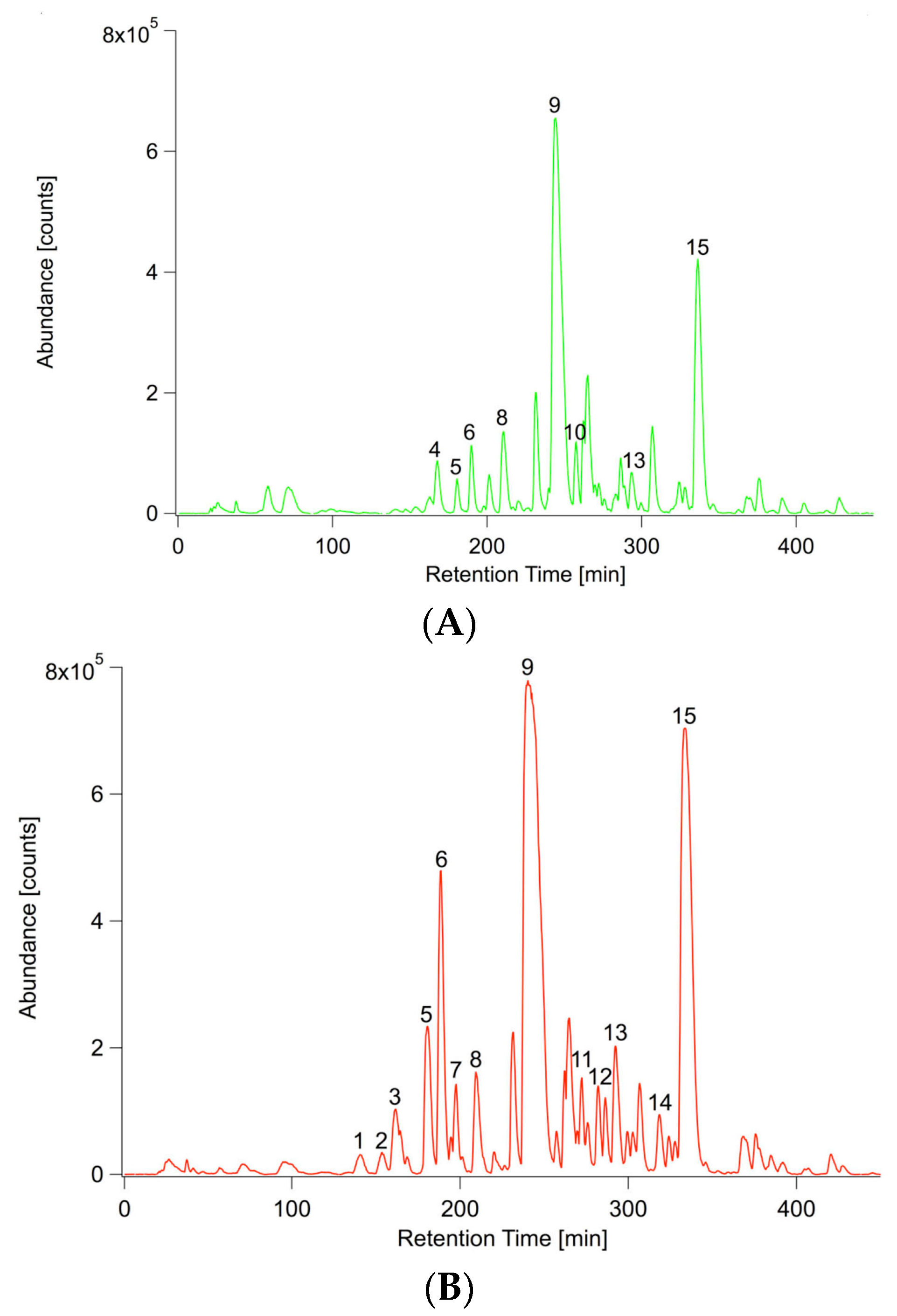
2.2. P. indica Alters the Exudation of Secondary Metabolites by A. thaliana Roots
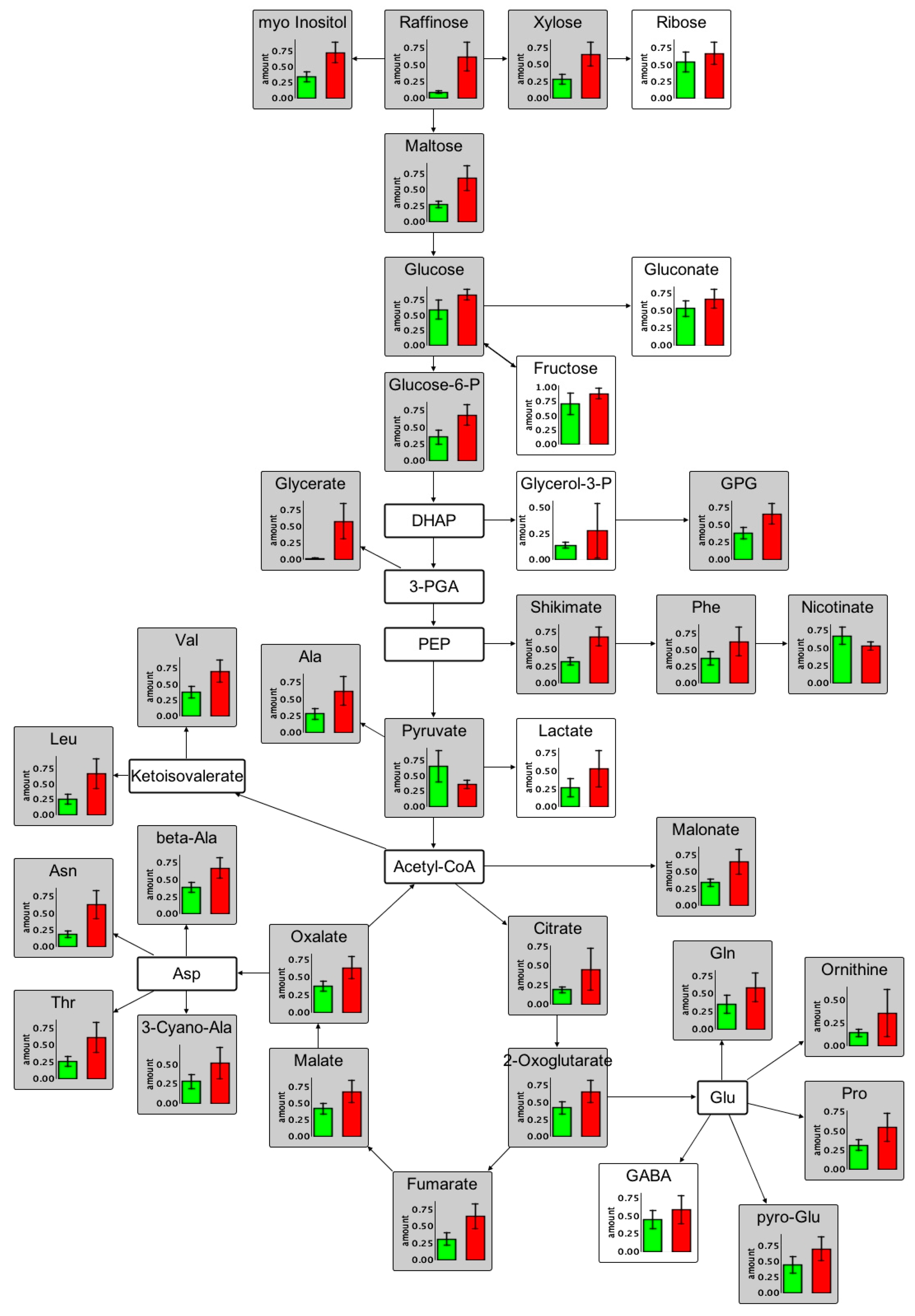
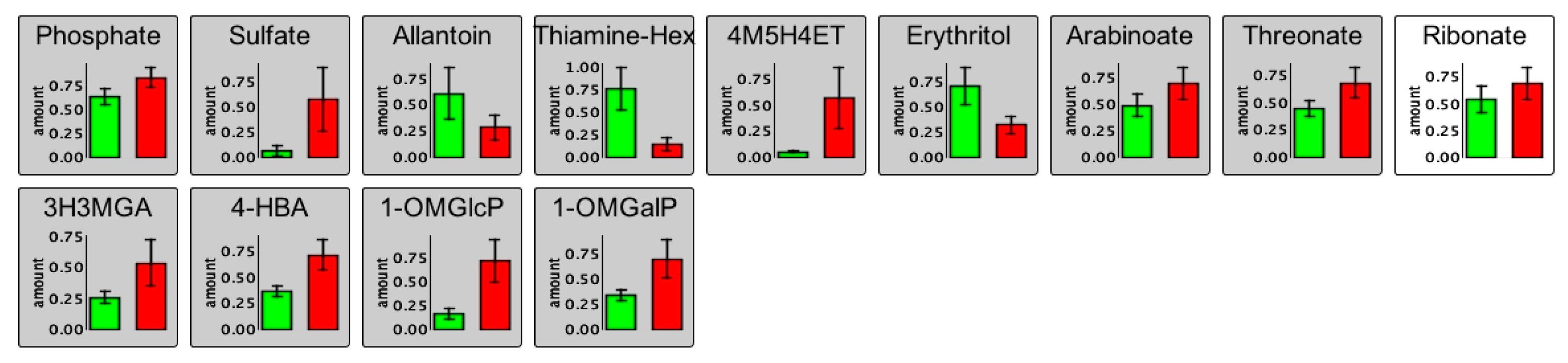
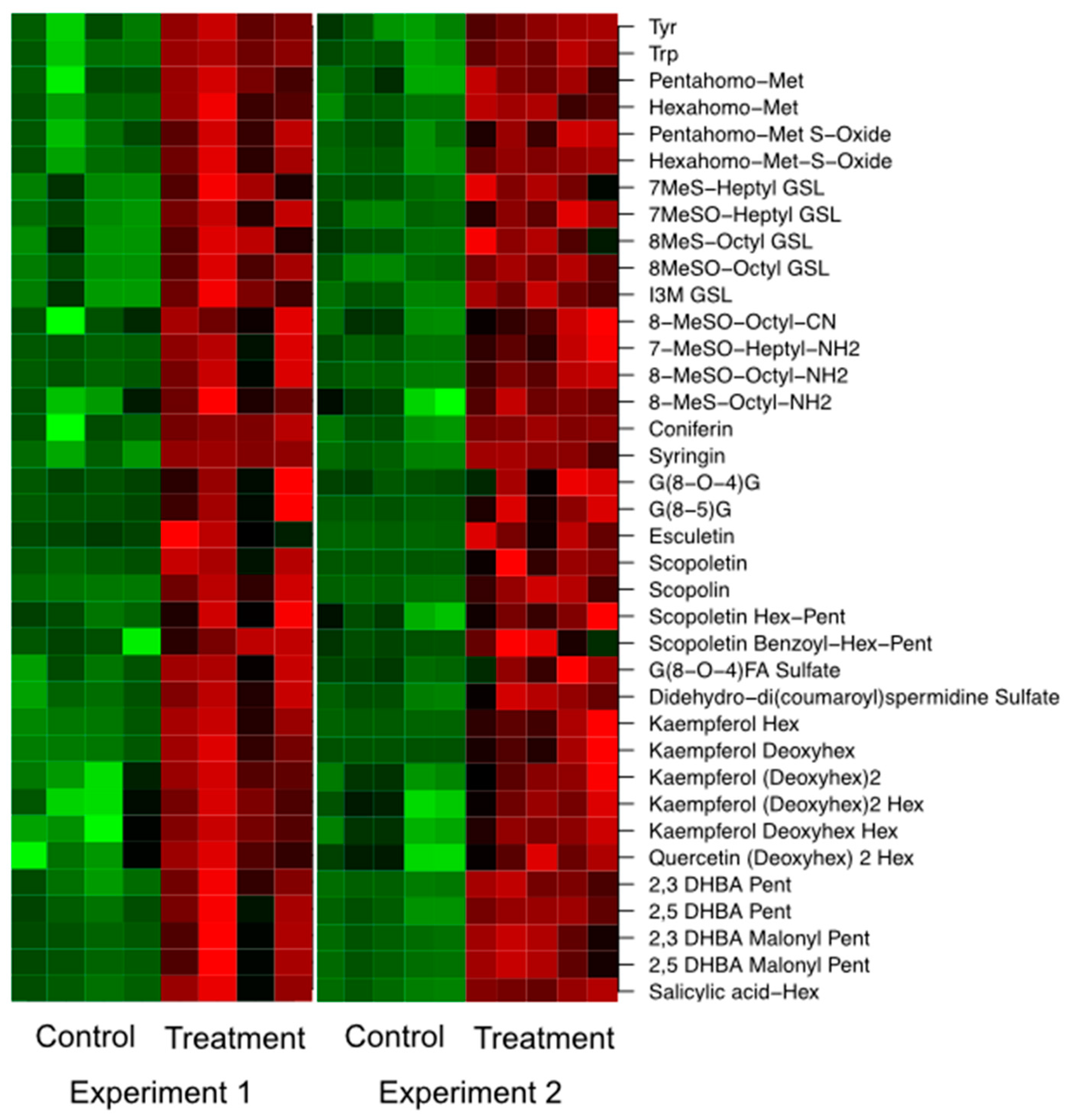
2.3. Changes in the Root Metabolism of A. thaliana
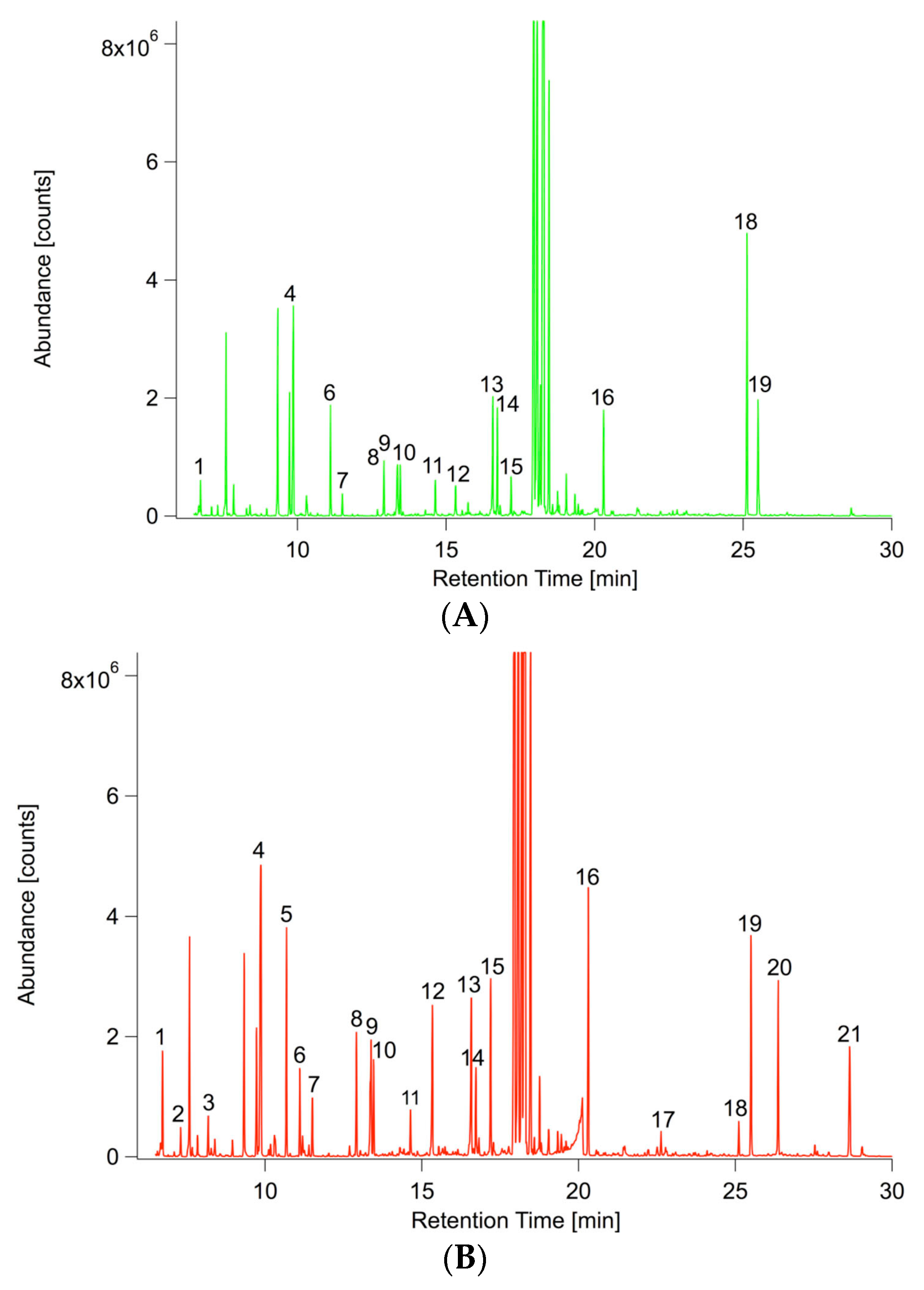
2.3.1. Non-Targeted GC/MS Based Metabolite Profiling Reveals Perturbations in the Primary Root Metabolism
2.3.2. LC/MS Based Non-Targeted Metabolite Profiling Shows an Induction of Aliphatic and Indolic Glucosinolate Metabolism, Flavonoids, and Oligolignols in Roots
3. Materials and Methods
3.1. Chemicals and Standards
3.2. Pre-Cultivation of P. indica
3.3. Conduction of Co-Cultivation Studies and Production of Plant Material
3.4. LC/MS-Based Metabolite Profiling
3.4.1. Preparation of Nutrient Solutions for LC/MS Analysis
3.4.2. Sample Preparation and Profiling of Tissue Material for LC/MS Analysis
3.4.3. Non-Targeted LC/MS-Based Profiling and Data Analysis
3.5. GC/MS Based Metabolite Profiling
3.5.1. Sample Preparation of Tissue Material
3.5.2. Preparation of Samples for Non-Targeted Metabolite Profiling and Analysis of GC/MS Profiles
3.6. Hormone Analysis
3.7. Microscope Images
3.8. Transcript Enrichment Analysis
3.9. Data Availability
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| A. thaliana | Arabidopsis thaliana |
| P. indica | Piriformospora indica |
| GC | Gas chromatography |
| UPLC | Ultraperformance liquid chromatography |
| ESI | Electrospray ionisation |
| QTOF | Quadupole time of flight mass spectrometer |
| SD | Standard deviation |
| ET | Ethylene |
| JA | Jasmonic acid |
| GSL | Glucosinolate |
Appendix A (Media for Co-Cultivation Studies)
Appendix B (UPLC/ESI-QTOFMS)
Appendix C (GC/EI-QMS)
Appendix D (Hormone Analysis)
Profiling of Root Tissue
Profiling of Root Exudates
References
- Verma, S.; Varma, A.; Rexer, K.H.; Hassel, A.; Kost, G.; Sarbhoy, A.; Bisen, P.; Bütehorn, B.; Franken, P. Piriformospora indica, gen. et sp. nov., a new root-colonizing fungus. Mycologia 1998, 90, 896–903. [Google Scholar] [CrossRef]
- Weiss, M.; Selosse, M.A.; Rexer, K.H.; Urban, A.; Oberwinkler, F. Sebacinales: A hitherto overlooked cosm of heterobasidiomycetes with a broad mycorrhizal potential. Mycol. Res. 2004, 108, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.; Bakshi, M.; Lou, B.; Hartmann, A.; Oelmueller, R. Piriformospora indica: A Novel Plant Growth-Promoting Mycorrhizal Fungus. Agric. Res. 2012, 1, 117–131. [Google Scholar] [CrossRef]
- Sun, C.; Shao, Y.; Vahabi, K.; Lu, J.; Bhattacharya, S.; Dong, S.; Yeh, K.-W.; Sherameti, I.; Lou, B.; Baldwin, I.T.; et al. The beneficial fungus Piriformospora indica protects Arabidopsis from Verticillium dahliae infection by downregulation plant defense responses. BMC Plant Biol. 2014, 14, 268. [Google Scholar] [CrossRef] [PubMed]
- Daneshkhah, R.; Cabello, S.; Rozanska, E.; Sobczak, M.; Grundler, F.M.W.; Wieczorek, K.; Hofmann, J. Piriformospora indica antagonizes cyst nematode infection and development in Arabidopsis roots. J. Exp. Bot. 2013, 64, 3763–3774. [Google Scholar] [CrossRef] [PubMed]
- Camehl, I.; Sherameti, I.; Seebald, E.; Johnson, J.M.; Oelmüller, R. Role of Defense Compounds in the Beneficial Interaction Between Arabidopsis thalina and Piriformospora indica. In Piriformospora indica: Sebacinales and Their Biotechnological Applications; Varma, A., Kost, G., Oelmüller, R., Eds.; Springer-Verlag Berlin Heidelberg: New York, NY, USA, 2013; Volume 33, pp. 239–250. [Google Scholar]
- Gill, S.S.; Gill, R.; Trivedi, D.K.; Anjum, N.A.; Sharma, K.K.; Ansari, M.W.; Ansari, A.A.; Johri, A.K.; Prasad, R.; Pereira, E.; et al. Piriformospora indica: Potential and Significance in Plant Stress Tolerance. Front. Microbiol. 2016, 7, 332. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Kamal, S.; Shakil, N.A.; Sherameti, I.; Oelmueller, R.; Dua, M.; Tuteja, N.; Johri, A.K.; Varma, A. The root endophyte fungus Piriformospora indica leads to early flowering, higher biomass and altered secondary metabolites of the medicinal plant, Coleus forskohlii. Plant Signal. Behav. 2012, 7, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Kamal, S.; Sharma, P.K.; Oelmueller, R.; Varma, A. Root endophyte Piriformospora indica DSM 11827 alters plant morphology, enhances biomass and antioxidant activity of medicinal plant Bacopa monniera. J. Basic Microbiol. 2013, 53, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.; Verma, S.; Sudha; Sahay, N.; Butehorn, B.; Franken, P. Piriformospora indica, a cultivable plant-growth-promoting root endophyte. Appl. Environ. Microbiol. 1999, 65, 2741–2744. [Google Scholar] [PubMed]
- Oelmueller, R.; Sherameti, I.; Tripathi, S.; Varma, A. Piriformospora indica, a cultivable root endophyte with multiple biotechnological applications. Symbiosis 2009, 49, 1–17. [Google Scholar] [CrossRef]
- Yadav, V.; Kumar, M.; Deep, D.K.; Kumar, H.; Sharma, R.; Tripathi, T.; Tuteja, N.; Saxena, A.K.; Johri, A.K. A phosphate transporter from the root endophytic fungus Piriformospora indica plays a role in phosphate transport to the host plant. J. Biol. Chem. 2010, 285, 26532–26544. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, M.; Vahabi, K.; Sherameti, I.; Oelmuller, R.; Bhattacharya, S.; Baldwin, I.; Varma, A.; Yeh, K.-W.; Johri, A.K. WRKY6 restricts Piriformospora indica-stimulated and phosphate-induced root development in Arabidopsis. BMC Plant Biol. 2015, 15, 305. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Yadav, V.; Kumar, H.; Sharma, R.; Singh, A.; Tuteja, N.; Johri, A.K. Piriformospora indica enhances plant growth by transferring phosphate. Plant Signal. Behav. 2011, 6, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Lahrmann, U.; Ding, Y.; Banhara, A.; Rath, M.; Hajirezaei, M.R.; Doehlemann, S.; von Wiren, N.; Parniske, M.; Zuccaro, A. Host-related metabolic cues affect colonization strategies of a root endophyte. Proc. Natl. Acad. Sci. USA 2013, 110, 13965–13970. [Google Scholar] [CrossRef] [PubMed]
- Vahabi, K.; Sherameti, I.; Bakshi, M.; Mrozinska, A.; Ludwig, A.; Reichelt, M.; Oelmuller, R. The interaction of Arabidopsis with Piriformospora indica shifts from initial transient stress induced by fungus-released chemical mediators to a mutualistic interaction after physical contact of the two symbionts. BMC Plant Biol. 2015, 15, 58. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, M.; Johnson, J.M.; Sherameti, I.; Hieno, A.; Tokizawa, M.; Yamamoto, Y.Y.; Nomoto, M.; Tada, Y.; Godfrey, R.; Obokata, J.; et al. High REDOX RESPONSIVE TRANSCRIPTION FACTOR1 Levels Result in Accumulation of Reactive Oxygen Species in Arabidopsis thaliana Shoots and Roots. Mol. Plant 2015, 8, 1253–1273. [Google Scholar] [CrossRef] [PubMed]
- Rafiqi, M.; Jelonek, L.; Akum, N.F.; Zhang, F.; Kogel, K.-H. Effector candidates in the secretome of Piriformospora indica, a ubiquitous plant-associated fungus. Front. Plant Sci. 2013, 4, 228. [Google Scholar] [CrossRef] [PubMed]
- Vadassery, J.; Oelmuller, R. Calcium signaling in pathogenic and beneficial plant microbe interactions what can we learn from the interaction between Piriformospora indica and Arabidopsis thaliana. Plant Signal. Behav. 2009, 4, 1024–1027. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.; Oelmueller, R. Agony to harmony—what decides? calcium signaling in beneficial and pathogenic plant—fungus interactions—what we can learn from the Arabidopsis/Piriformospora indica symbiosis. Mol. Microb. Ecol. Rhizosphere 2013, 2, 833–850. [Google Scholar]
- Camehl, I.; Sherameti, I.; Venus, Y.; Bethke, G.; Varma, A.; Lee, J.; Oelmueller, R. Ethylene signalling and ethylene-targeted transcription factors are required to balance beneficial and nonbeneficial traits in the symbiosis between the endophytic fungus Piriformospora indica and Arabidopsis thaliana. New Phytol. 2010, 185, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Camehl, I.; Oelmueller, R. Do ethylene response factors-9 and -14 repress PR gene expression in the interaction between Piriformospora indica and Arabidopsis? Plant Signal. Behav. 2010, 5, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Vadassery, J.; Ritter, C.; Venus, Y.; Camehl, I.; Varma, A.; Shahollari, B.; Novak, O.; Strnad, M.; Ludwig-Mueller, J.; Oelmueller, R. The role of auxins and cytokinins in the mutualistic interaction between Arabidopsis and Piriformospora indica. Mol. Plant Microbe Interact. 2008, 21, 1371–1383. [Google Scholar] [CrossRef] [PubMed]
- Lahrmann, U.; Strehmel, N.; Langen, G.; Frerigmann, H.; Leson, L.; Ding, Y.; Scheel, D.; Herklotz, S.; Hilbert, M.; Zuccaro, A. Mutualistic root endophytism is not associated with the reduction of saprotrophic traits and requires a noncompromised plant innate immunity. New Phytol. 2015, 207, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Vahabi, K.; Camehl, I.; Sherameti, I.; Oelmueller, R. Growth of Arabidopsis seedlings on high fungal doses of Piriformospora indica has little effect on plant performance, stress, and defense gene expression in spite of elevated jasmonic acid and jasmonic acid-isoleucine levels in the roots. Plant Signal. Behav. 2013, 8, e26301. [Google Scholar] [CrossRef] [PubMed]
- Nongbri, P.L.; Johnson, J.M.; Sherameti, I.; Glawischnig, E.; Halkier, B.A.; Oelmueller, R. Indole-3-acetaldoxime-derived compounds restrict root colonization in the beneficial interaction between Arabidopsis roots and the endophyte Piriformospora indica. Mol. Plant Microbe Interact. 2012, 25, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Zuccaro, A.; Lahrmann, U.; Gueldener, U.; Langen, G.; Pfiffi, S.; Biedenkopf, D.; Wong, P.; Samans, B.; Grimm, C.; Basiewicz, M.; et al. Endophytic life strategies decoded by genome and transcriptome analyses of the mutualistic root symbiont Piriformospora indica. PLoS Pathog. 2011, 7, e1002290. [Google Scholar] [CrossRef] [PubMed]
- Lahrmann, U.; Zuccaro, A. Opprimo ergo sum-evasion and suppression in the root endophytic fungus Piriformospora indica. Mol. Plant Microbe Interact. 2012, 25, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, C.; Chapman, A.; Fellermeier, F.; Choudhary, M.; Scheel, D.; Glawischnig, E. The Biosynthetic Pathway of Indole-3-Carbaldehyde and Indole-3-Carboxylic Acid Derivatives in Arabidopsis. Plant Physiol. 2014, 165, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Peskan-Berghoefer, T.; Shahollari, B.; Giong, P.H.; Hehl, S.; Markert, C.; Blanke, V.; Kost, G.; Varma, A.; Oelmueller, R. Association of Piriformospora indica with Arabidopsis thaliana roots represents a novel system to study beneficial plant-microbe interactions and involves early plant protein modifications in the endoplasmic reticulum and at the plasma membrane. Physiol. Plant. 2004, 122, 465–477. [Google Scholar] [CrossRef]
- Peskan-Berghoefer, T.; Vilches-Barro, A.; Mueller, T.M.; Glawischnig, E.; Reichelt, M.; Gershenzon, J.; Rausch, T. Sustained exposure to abscisic acid enhances the colonization potential of the mutualist fungus Piriformospora indica on Arabidopsis thaliana roots. New Phytol. 2015, 208, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Cosme, M.; Wurst, S.; Cosme, M.; Franken, P.; Cosme, M.; Lu, J.; Erb, M.; Lu, J.; Erb, M.; Stout, M.J.; et al. A fungal endophyte helps plants to tolerate root herbivory through changes in gibberellin and jasmonate signaling. New Phytol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Khatabi, B.; Schafer, P. Ethylene in mutualistic symbioses. Plant Signal. Behav. 2012, 7, 1634–1638. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, P.; Pfiffi, S.; Voll, L.M.; Zajic, D.; Chandler, P.M.; Waller, F.; Scholz, U.; Pons-Kuehnemann, J.; Sonnewald, S.; Sonnewald, U.; et al. Manipulation of plant innate immunity and gibberellin as factor of compatibility in the mutualistic association of barley roots with Piriformospora indica. Plant J. 2009, 59, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Komarova, N.Y.; Thor, K.; Gubler, A.; Meier, S.; Dietrich, D.; Weichert, A.; Suter Grotemeyer, M.; Tegeder, M.; Rentsch, D. AtPTR1 and AtPTR5 transport dipeptides in planta. Plant Physiol. 2008, 148, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.; Schaarschmidt, S. The role of jasmonates in mutualistic symbioses between plants and soil-born microorganisms. Phytochemistry 2009, 70, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O'Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- MetaboLights. Available online: http://www.ebi.ac.uk/metabolights/MTBLS341 (accessed on 3 July 2016).
- Lommen, A. MetAlign: interface-driven, versatile metabolomics tool for hyphenated full-scan mass spectrometry data preprocessing. Anal. Chem. 2009, 81, 3079–3086. [Google Scholar] [CrossRef] [PubMed]
- Luedemann, A.; Strassburg, K.; Erban, A.; Kopka, J. TagFinder for the quantitative analysis of gas chromatography-mass spectrometry (GC-MS)-based metabolite profiling experiments. Bioinformatics 2008, 24, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Golm Metabolome Database. Available online: http://gmd.mpimp-golm.mpg.de/.
- Ziegler, J.; Qwegwer, J.; Schubert, M.; Erickson, J.L.; Schattat, M.; Burstenbinder, K.; Grubb, C.D.; Abel, S. Simultaneous analysis of apolar phytohormones and 1-aminocyclopropan-1-carboxylic acid by high performance liquid chromatography/electrospray negative ion tandem mass spectrometry via 9-fluorenylmethoxycarbonyl chloride derivatization. J. Chromatogr. A 2014, 1362, 102–109. [Google Scholar] [CrossRef] [PubMed]
- DAVID Bioinformatics Resources 6.7. Available online: https://david.ncifcrf.gov/.
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2008, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.W.; Gill, S.S.; Tuteja, N. Piriformospora Indica a Powerful Tool for Crop Improvement. Proc. Indian Natl. Sci. Acad. 2014, 80, 317–324. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Fold Enrichment | p-Value * |
|---|---|---|
| ath00966: Glucosinolate biosynthesis | 10.4 | 8.89 × 10−8 |
| ath00940: Phenylpropanoid biosynthesis | 3.8 | 5.84 × 10−7 |
| ath00360: Phenylalanine metabolism | 3.7 | 6.21 × 10−5 |
| ath00903: Limonene and pinene degradation | 3.8 | 1.12 × 10−4 |
| ath00680: Methane metabolism | 3.5 | 1.70 × 10−4 |
| ath00945: Stilbenoid, diarylheptanoid and gingerol biosynthesis | 3.7 | 2.20 × 10−4 |
| ath00910: Nitrogen metabolism | 3.9 | 5.56 × 10−3 |
| ath00260: Glycine, serine and threonine metabolism | 3.7 | 7.07 × 10−3 |
| ath00460: Cyanoamino acid metabolism | 5.0 | 1.25 × 10−2 |
| ath00960: Tropane, piperidine and pyridine alkaloid biosynthesis | 5.5 | 2.05 × 10−2 |
| ath01070: Biosynthesis of plant hormones | 1.6 | 3.54 × 10−2 |
| ath00400: Phenylalanine, tyrosine and tryptophan biosynthesis | 3.3 | 4.44 × 10−2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strehmel, N.; Mönchgesang, S.; Herklotz, S.; Krüger, S.; Ziegler, J.; Scheel, D. Piriformospora indica Stimulates Root Metabolism of Arabidopsis thaliana. Int. J. Mol. Sci. 2016, 17, 1091. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071091
Strehmel N, Mönchgesang S, Herklotz S, Krüger S, Ziegler J, Scheel D. Piriformospora indica Stimulates Root Metabolism of Arabidopsis thaliana. International Journal of Molecular Sciences. 2016; 17(7):1091. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071091
Chicago/Turabian StyleStrehmel, Nadine, Susann Mönchgesang, Siska Herklotz, Sylvia Krüger, Jörg Ziegler, and Dierk Scheel. 2016. "Piriformospora indica Stimulates Root Metabolism of Arabidopsis thaliana" International Journal of Molecular Sciences 17, no. 7: 1091. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071091