
Enhancement of Anti-Hypoxic Activity and Differentiation of Cardiac Stem Cells by Supernatant Fluids from Cultured Macrophages that Phagocytized Dead Mesenchymal Stem Cells


Abstract
:1. Introduction
2. Results
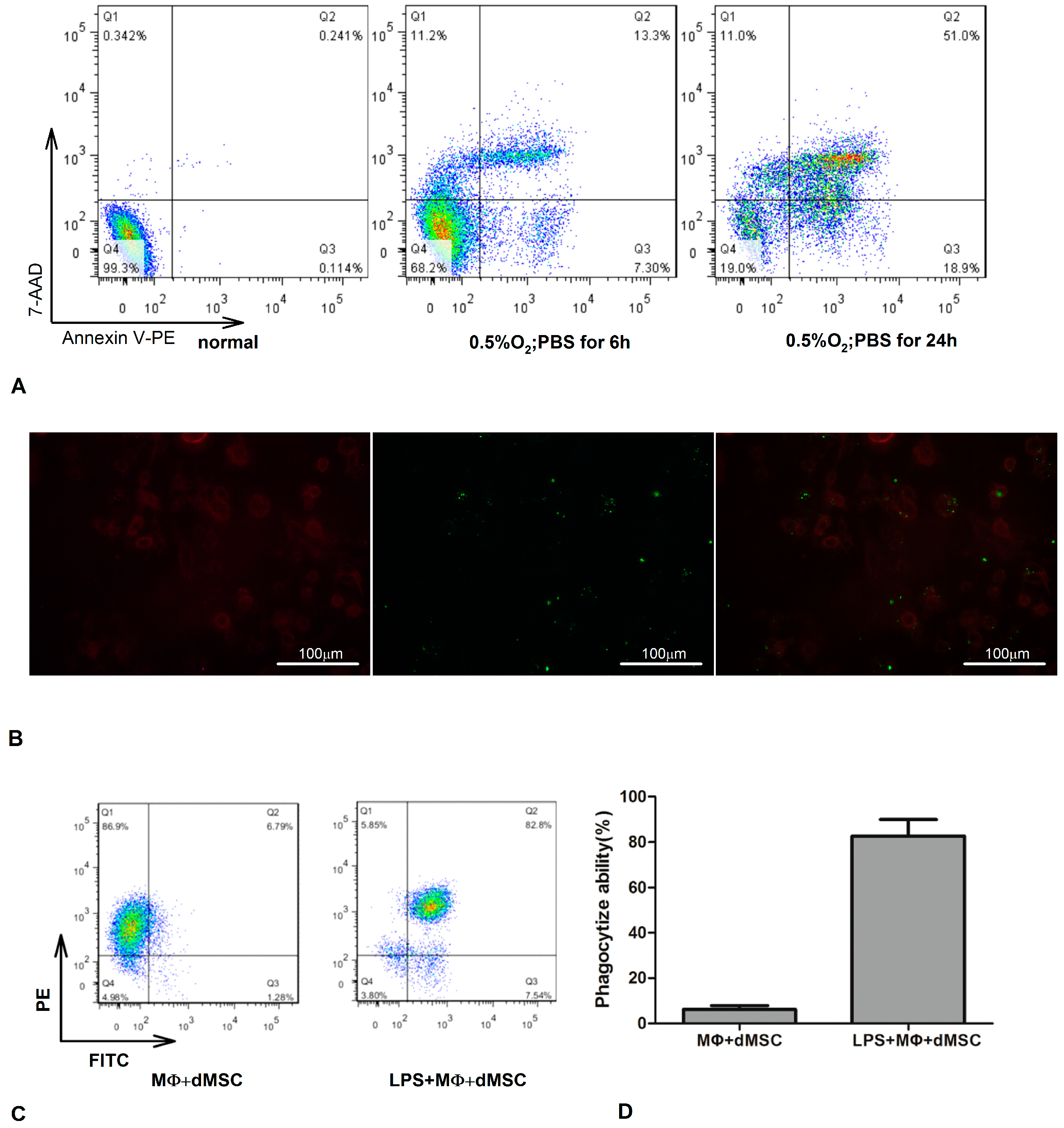
2.1. Lipopolysaccharide (LPS)-Stimulated Macrophages Phagocytosed Dead Mesenchymal Stem Cells (dMSCs)
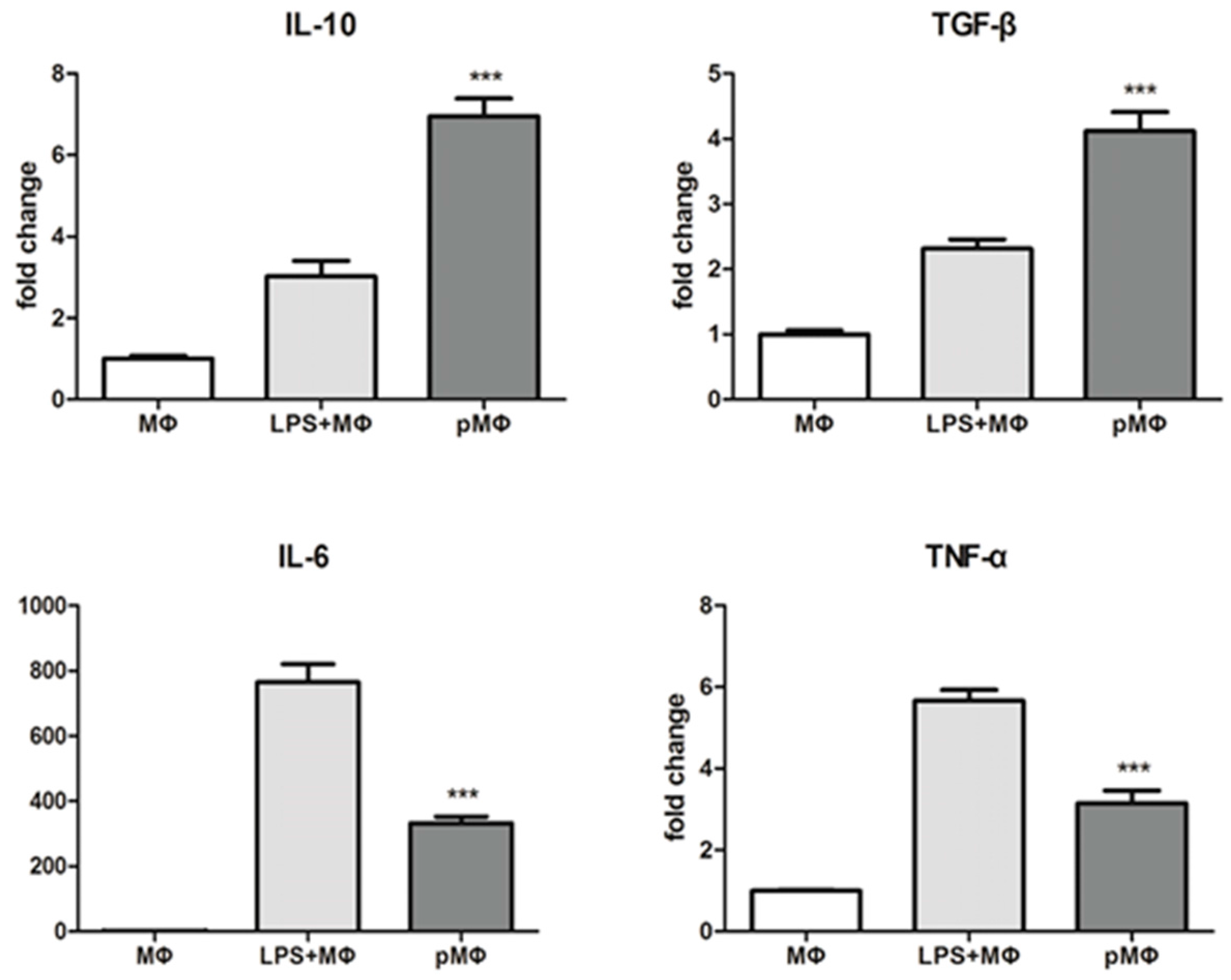
2.2. Inflammatory Cytokines Released by Macrophages (MΦ), Lipopolysaccharide (LPS) + MΦ, Phagocytized by Macrophages (pMΦ)
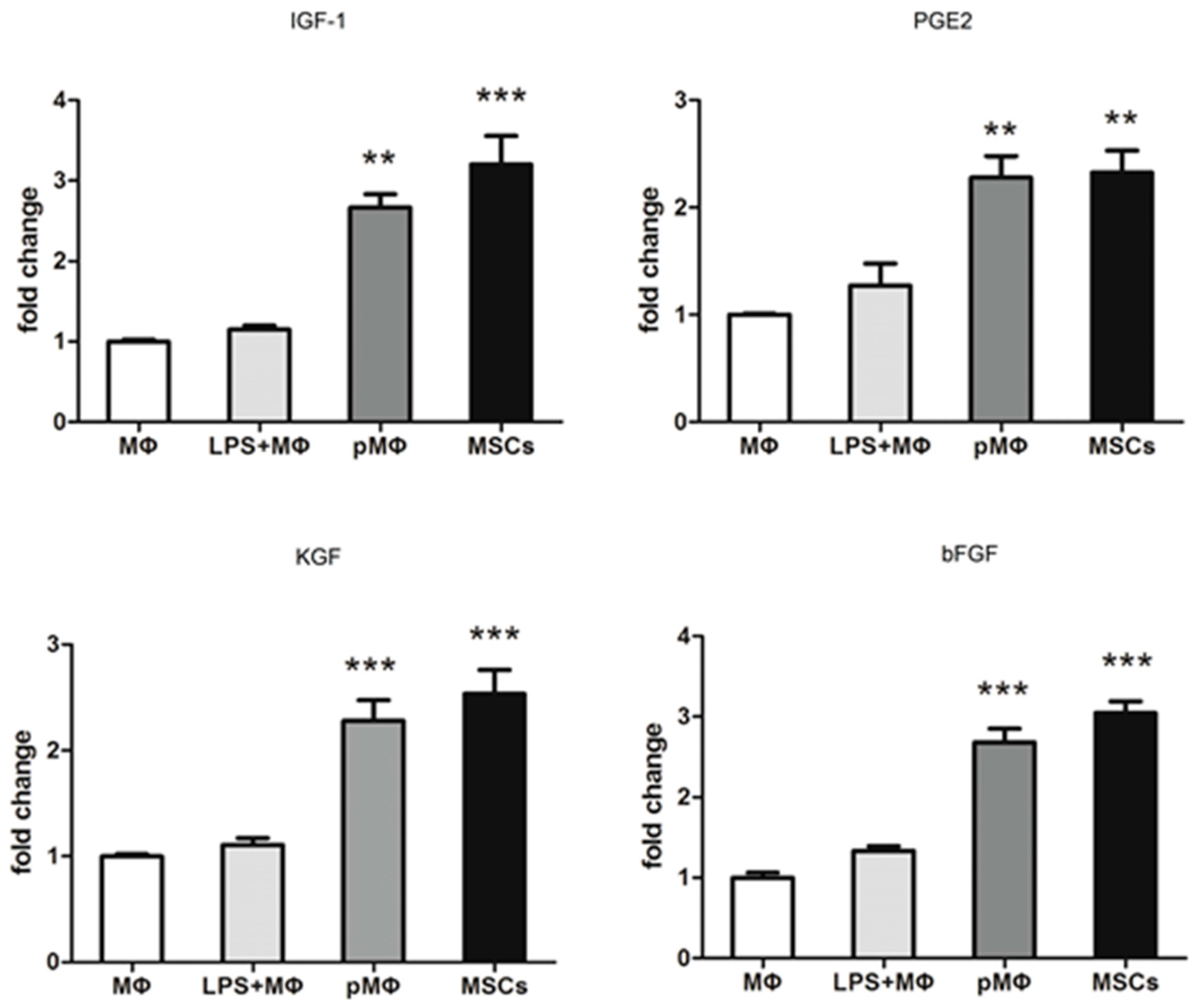
2.3. Growth Factors Released by pMΦ, MΦ and LPS + MΦ and Mesenchymal Stem Cells (MSCs)
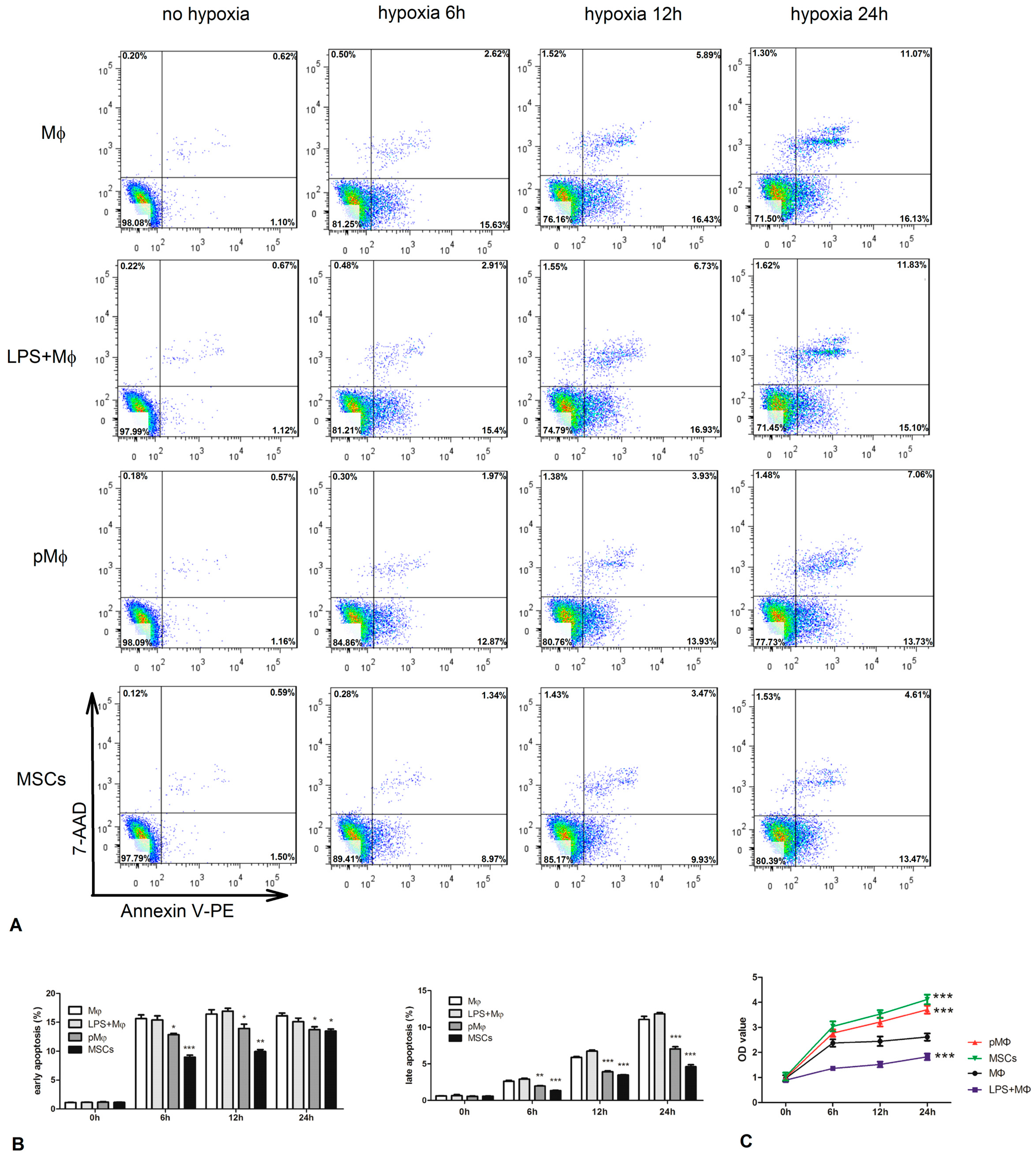
2.4. Supernatant Fluids of pMΦ Improved Anti-Anoxia and Proliferation of Cardiac Stem Cells (CSCs)
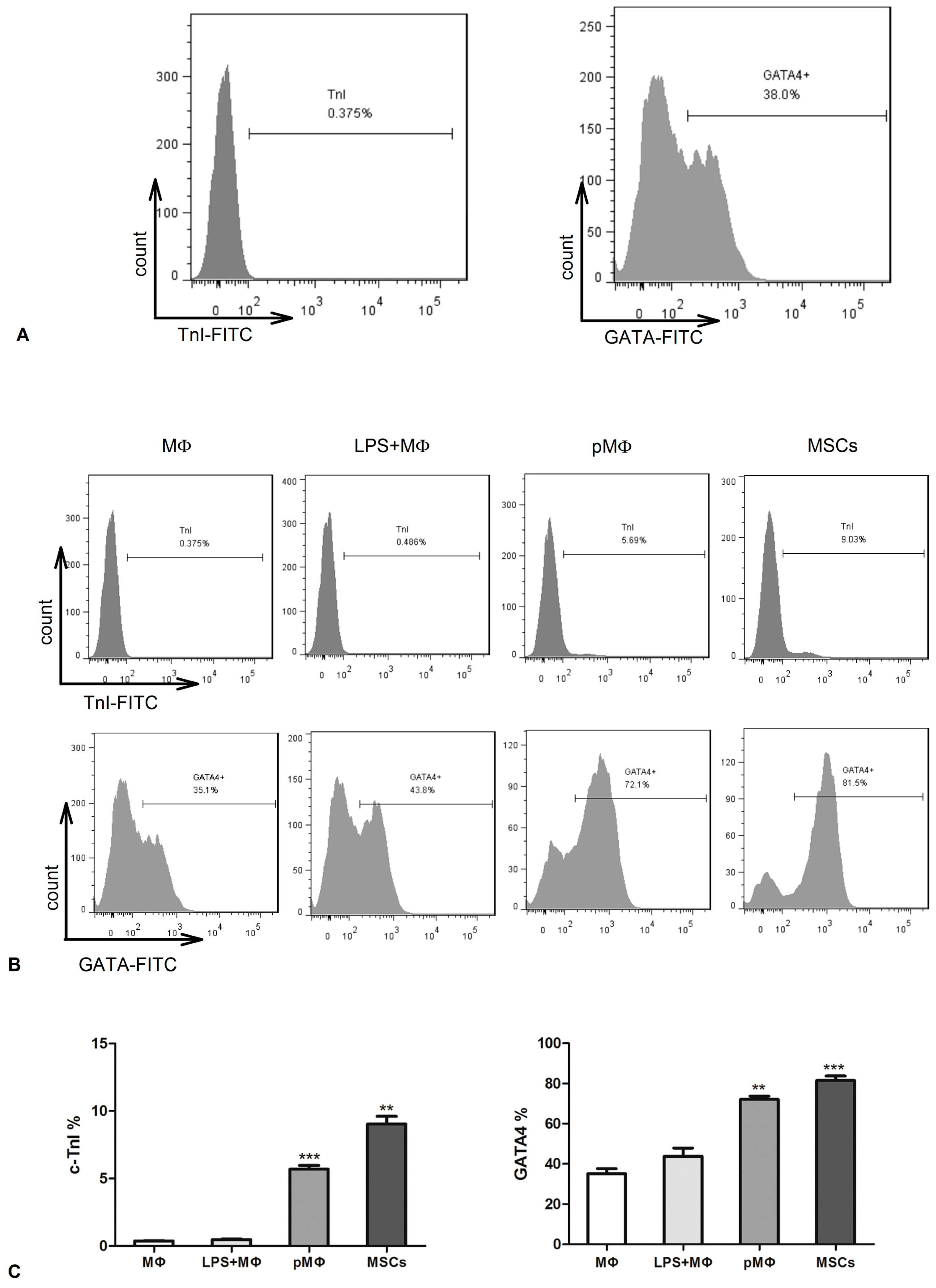
2.5. Supernatant Fluids of pMΦ Induced CSCs Differentiation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Isolation and Culture
4.3. GFP Transfection
4.4. Detection of MSC Apoptosis and Isolation of pMΦ
4.5. Cell Viability Assay
4.6. Real-Time PCR Analysis
4.7. Flow Cytometry (FCM) Analysis
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gaziano, T.A. Cardiovascular disease in the developing world and its cost-effective management. Circulation 2005, 112, 3547–3553. [Google Scholar] [CrossRef] [PubMed]
- Segers, V.F.; Lee, R.T. Stem-cell therapy for cardiac disease. Nature 2008, 451, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Schachinger, V.; Assmus, B.; Britten, M.B.; Honold, J.; Lehmann, R.; Teupe, C.; Abolmaali, N.D.; Vogl, T.J.; Hofmann, W.K.; Martin, H.; et al. Transplantation of progenitor cells and regeneration enhancement in acute myocardial infarction: Final one-year results of the topcare-ami trial. J. Am. Coll. Cardiol. 2004, 44, 1690–1699. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Martin, B.J. Mesenchymal stem cells and their potential as cardiac therapeutics. Circ. Res. 2004, 95, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Hass, R.; Kasper, C.; Bohm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyahara, Y.; Nagaya, N.; Kataoka, M.; Yanagawa, B.; Tanaka, K.; Hao, H.; Ishino, K.; Ishida, H.; Shimizu, T.; Kangawa, K.; et al. Monolayered mesenchymal stem cells repair scarred myocardium after myocardial infarction. Nat. Med. 2006, 12, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Price, M.J.; Chou, C.C.; Frantzen, M.; Miyamoto, T.; Kar, S.; Lee, S.; Shah, P.K.; Martin, B.J.; Lill, M.; Forrester, J.S.; et al. Intravenous mesenchymal stem cell therapy early after reperfused acute myocardial infarction improves left ventricular function and alters electrophysiologic properties. Int. J. Cardiol. 2006, 111, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, Y.; Deng, Z. Pre-conditioned mesenchymal stem cells: A better way for cell-based therapy. Stem Cell Res. Ther. 2013, 4, 63. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.B.; Chen, L.J.; Tang, Y.L.; Ma, G.S.; Shen, C.X.; Qi, C.M.; Zhu, Q.; Yao, Y.Y.; Liu, N.F. Ho-1 gene overexpression enhances the beneficial effects of superparamagnetic iron oxide labeled bone marrow stromal cells transplantation in swine hearts underwent ischemia/reperfusion: An MRI study. Basic Res. Cardiol. 2010, 105, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Fu, C.; Song, L.; Yao, Y.; Zhang, X.; Chen, Z.; Li, Y.; Ma, G.; Shen, C. Exposure to supernatants of macrophages that phagocytized dead mesenchymal stem cells improves hypoxic cardiomyocytes survival. Int. J. Cardiol. 2013, 165, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Laugwitz, K.L.; Moretti, A.; Lam, J.; Gruber, P.; Chen, Y.; Woodard, S.; Lin, L.Z.; Cai, C.L.; Lu, M.M.; Reth, M.; et al. Postnatal isl1 + cardioblasts enter fully differentiated cardiomyocyte lineages. Nature 2005, 433, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Bradfute, S.B.; Gallardo, T.D.; Nakamura, T.; Gaussin, V.; Mishina, Y.; Pocius, J.; Michael, L.H.; Behringer, R.R.; Garry, D.J.; et al. Cardiac progenitor cells from adult myocardium: Homing, differentiation, and fusion after infarction. Proc. Natl. Acad. Sci. USA 2003, 100, 12313–12318. [Google Scholar] [CrossRef] [PubMed]
- Urbich, C.; Aicher, A.; Heeschen, C.; Dernbach, E.; Hofmann, W.K.; Zeiher, A.M.; Dimmeler, S. Soluble factors released by endothelial progenitor cells promote migration of endothelial cells and cardiac resident progenitor cells. J. Mol. Cell. Cardiol. 2005, 39, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.L.; Rokosh, G.; Sanganalmath, S.K.; Yuan, F.; Sato, H.; Mu, J.; Dai, S.; Li, C.; Chen, N.; Peng, Y.; et al. Intracoronary administration of cardiac progenitor cells alleviates left ventricular dysfunction in rats with a 30-day-old infarction. Circulation 2010, 121, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Italiani, P.; Boraschi, D. From monocytes to M1/M2 macrophages: Phenotypical vs. Functional differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [PubMed]
- Janssens, S.; Dubois, C.; Bogaert, J.; Theunissen, K.; Deroose, C.; Desmet, W.; Kalantzi, M.; Herbots, L.; Sinnaeve, P.; Dens, J.; et al. Autologous bone marrow-derived stem-cell transfer in patients with ST-segment elevation myocardial infarction: Double-blind, randomised controlled trial. Lancet 2006, 367, 113–121. [Google Scholar] [CrossRef]
- Barbash, I.M.; Chouraqui, P.; Baron, J.; Feinberg, M.S.; Etzion, S.; Tessone, A.; Miller, L.; Guetta, E.; Zipori, D.; Kedes, L.H.; et al. Systemic delivery of bone marrow-derived mesenchymal stem cells to the infarcted myocardium: Feasibility, cell migration, and body distribution. Circulation 2003, 108, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Winkler, T.; von Roth, P.; Schuman, M.R.; Sieland, K.; Stoltenburg-Didinger, G.; Taupitz, M.; Perka, C.; Duda, G.N.; Matziolis, G. In vivo visualization of locally transplanted mesenchymal stem cells in the severely injured muscle in rats. Tissue Eng. A 2008, 14, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Lichtenauer, M.; Mildner, M.; Baumgartner, A.; Hasun, M.; Werba, G.; Beer, L.; Altmann, P.; Roth, G.; Gyongyosi, M.; Podesser, B.K.; et al. Intravenous and intramyocardial injection of apoptotic white blood cell suspensions prevents ventricular remodelling by increasing elastin expression in cardiac scar tissue after myocardial infarction. Basic Res. Cardiol. 2011, 106, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Paidassi, H.; Tacnet-Delorme, P.; Arlaud, G.J.; Frachet, P. How phagocytes track down and respond to apoptotic cells. Crit. Rev. Immunol. 2009, 29, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Zou, X.B.; Chai, Y.F.; Yao, Y.M. Macrophage polarization in inflammatory diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Labonte, A.C.; Tosello-Trampont, A.C.; Hahn, Y.S. The role of macrophage polarization in infectious and inflammatory diseases. Mol. Cells 2014, 37, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Tobacman, L.S. Thin filament-mediated regulation of cardiac contraction. Annu. Rev. Physiol. 1996, 58, 447–481. [Google Scholar] [CrossRef] [PubMed]
- Bodor, G.S.; Survant, L.; Voss, E.M.; Smith, S.; Porterfield, D.; Apple, F.S. Cardiac troponin t composition in normal and regenerating human skeletal muscle. Clin. Chem. 1997, 43, 476–484. [Google Scholar] [PubMed]
- Skeik, N.; Patel, D.C. A review of troponins in ischemic heart disease and other conditions. Int. J. Angiol. 2007, 16, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Clause, K.C.; Tchao, J.; Powell, M.C.; Liu, L.J.; Huard, J.; Keller, B.B.; Tobita, K. Developing cardiac and skeletal muscle share fast-skeletal myosin heavy chain and cardiac troponin-I expression. PLoS ONE 2012, 7, e40725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, J.E., 3rd; Bodor, G.S.; Davila-Roman, V.G.; Delmez, J.A.; Apple, F.S.; Ladenson, J.H.; Jaffe, A.S. Cardiac troponin I. A marker with high specificity for cardiac injury. Circulation 1993, 88, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.J.; Evans, T. Regulation of cardiac myocyte apoptosis by the GATA-4 transcription factor. Life Sci. 2004, 74, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Peterkin, T.; Gibson, A.; Loose, M.; Patient, R. The roles of GATA-4, -5 and -6 in vertebrate heart development. Semin. Cell Dev. Biol. 2005, 16, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Pourrajab, F.; Babaei Zarch, M.; Baghi Yazdi, M.; Rahimi Zarchi, A.; Vakili Zarch, A. Application of stem cell/growth factor system, as a multimodal therapy approach in regenerative medicine to improve cell therapy yields. Int. J. Cardiol. 2014, 173, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.; Tilokee, E.L.; Latham, N.; Mount, S.; Rafatian, G.; Strydhorst, J.; Ye, B.; Boodhwani, M.; Chan, V.; Ruel, M.; et al. Paracrine engineering of human cardiac stem cells with insulin-like growth factor 1 enhances myocardial repair. J. Am. Heart Assoc. 2015, 4, e002104. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, S.; Greder, L.; Dutton, J.; Keirstead, S.A.; Lepley, M.; Zhang, L.; Kaufman, D.; Zhang, J. Effective cardiac myocyte differentiation of human induced pluripotent stem cells requires vegf. PLoS ONE 2013, 8, e53764. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Zhu, W.; Cheng, M.; Chen, L.; Zhang, J.; Sun, T.; Kishore, R.; Phillips, M.I.; Losordo, D.W.; Qin, G. Hypoxic preconditioning enhances the benefit of cardiac progenitor cell therapy for treatment of myocardial infarction by inducing CXCR4 expression. Circ. Res. 2009, 104, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Shen, L.; Qian, K.; Phillips, M.I. A novel two-step procedure to expand cardiac Sca-1 + cells clonally. Biochem. Biophys. Res. Commun. 2007, 359, 877–883. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward and Reverse of Prime |
|---|---|
| IL-10 | 5’-TGCACTACCAAAGCCACAAG-3’; 5’-TGATCCTCATGCCAGTCAGT-3’ |
| Il-6 | 5’-TCTGCAAGAGACTTCCATCCA-3’; 5’-AGTCTCCTCTCCGGACTTGT-3’ |
| TNF-α | 5’-GGTGCCTATGTCTCAGCCTC-3’; 5’-CCACTTGGTGGTTTGTGAGTG-3’ |
| TGF-β | 5’-ATTCCTGGCGTTACCTTGG-3’; 5’-AGCCCTGTATTCCGTCTCCT |
| IGF-1 | 5’-GGGCTGAGCTGGTGGATG-3’; 5’-CTCCAGTCTCCTCAGATCAC-3’ |
| PGE2 | 5’-GGAGACTCTTCGAGGAGCACTT-3’;5’-GGCGATTTAGCAGCAGATATAAGAA-3’ |
| KGF | 5’-CTGCCAACTCTGCTCTACAG-3’; 5’-TCCAACTGCCACGGTCCTGAT-3’ |
| bFGF | 5’-AAGGGAGTGTGTGCCAACC-3’; 5’-GCCCAGTTCGTTTCAGTGC-3’ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Jin, X.; Zhou, Z.; Shen, C. Enhancement of Anti-Hypoxic Activity and Differentiation of Cardiac Stem Cells by Supernatant Fluids from Cultured Macrophages that Phagocytized Dead Mesenchymal Stem Cells. Int. J. Mol. Sci. 2016, 17, 1175. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071175
Liu L, Jin X, Zhou Z, Shen C. Enhancement of Anti-Hypoxic Activity and Differentiation of Cardiac Stem Cells by Supernatant Fluids from Cultured Macrophages that Phagocytized Dead Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2016; 17(7):1175. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071175
Chicago/Turabian StyleLiu, Liang, Xian Jin, Zhong’e Zhou, and Chengxing Shen. 2016. "Enhancement of Anti-Hypoxic Activity and Differentiation of Cardiac Stem Cells by Supernatant Fluids from Cultured Macrophages that Phagocytized Dead Mesenchymal Stem Cells" International Journal of Molecular Sciences 17, no. 7: 1175. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17071175