
BL-038, a Benzofuran Derivative, Induces Cell Apoptosis in Human Chondrosarcoma Cells through Reactive Oxygen Species/Mitochondrial Dysfunction and the Caspases Dependent Pathway


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
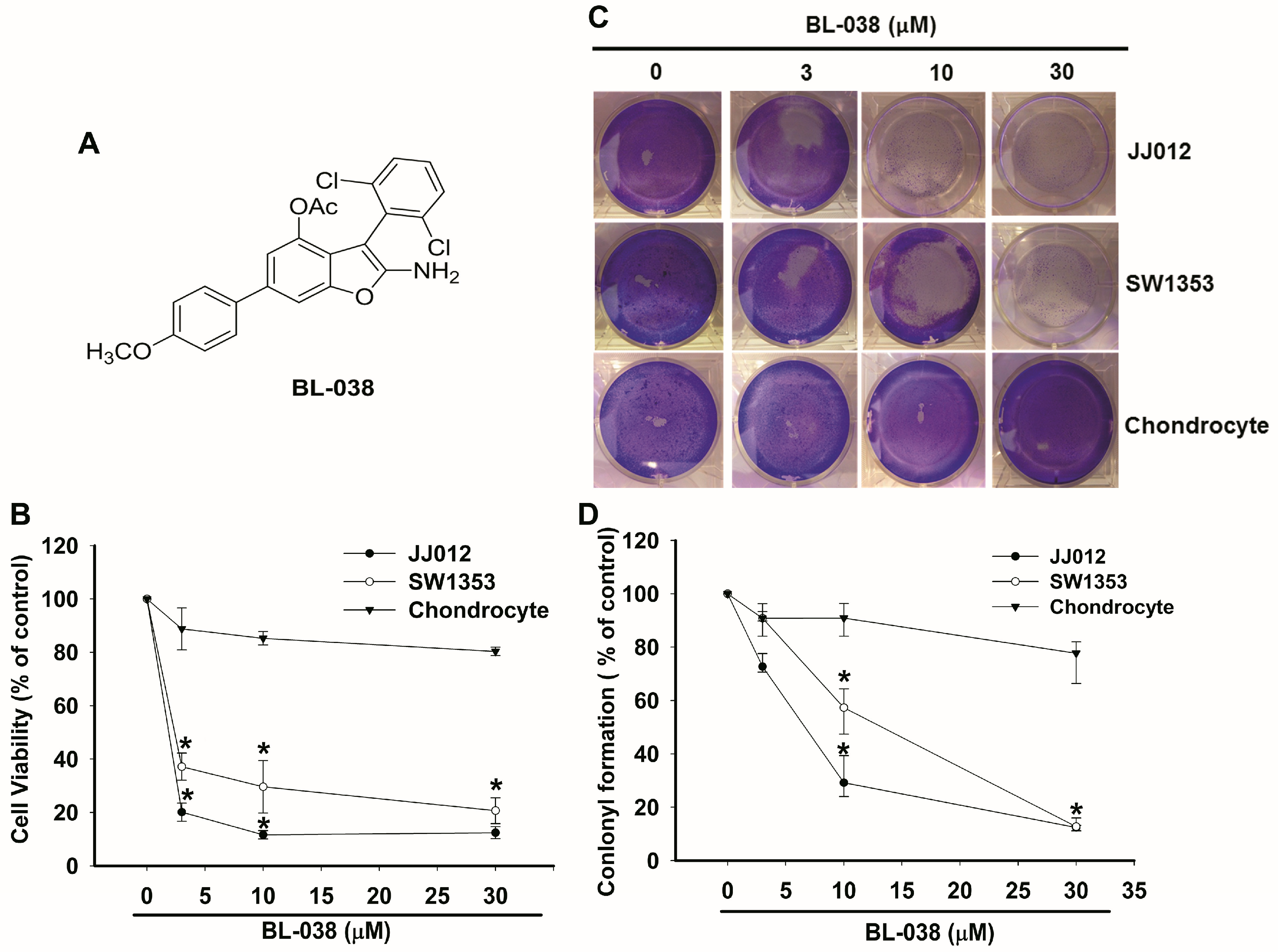
2.1. BL-038 Inhibits the Growth of Human Chondrosarcoma Cells
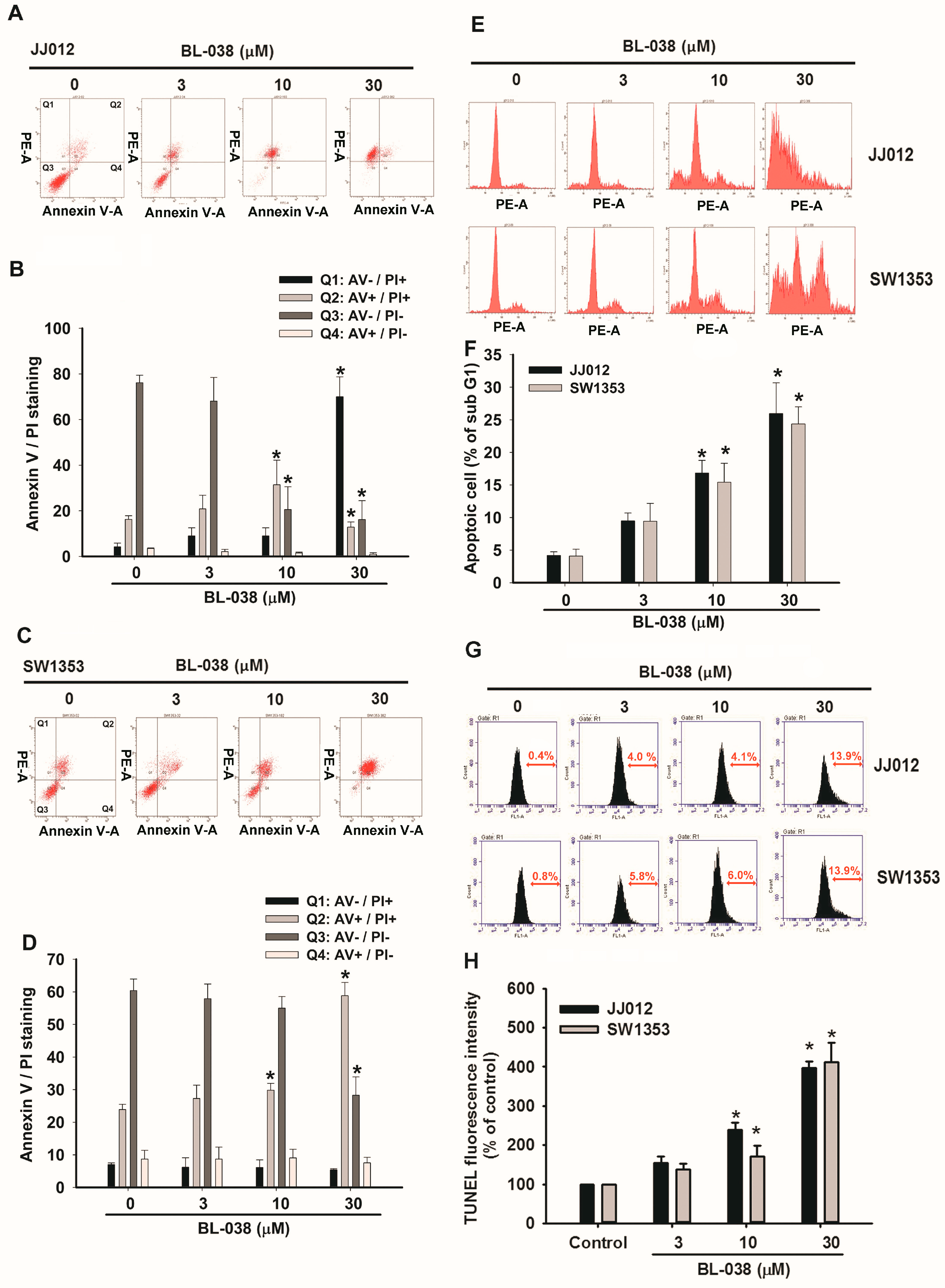
2.2. BL-038 Induces Apoptosis and Cell Migration in Human Chondrosarcoma Cells
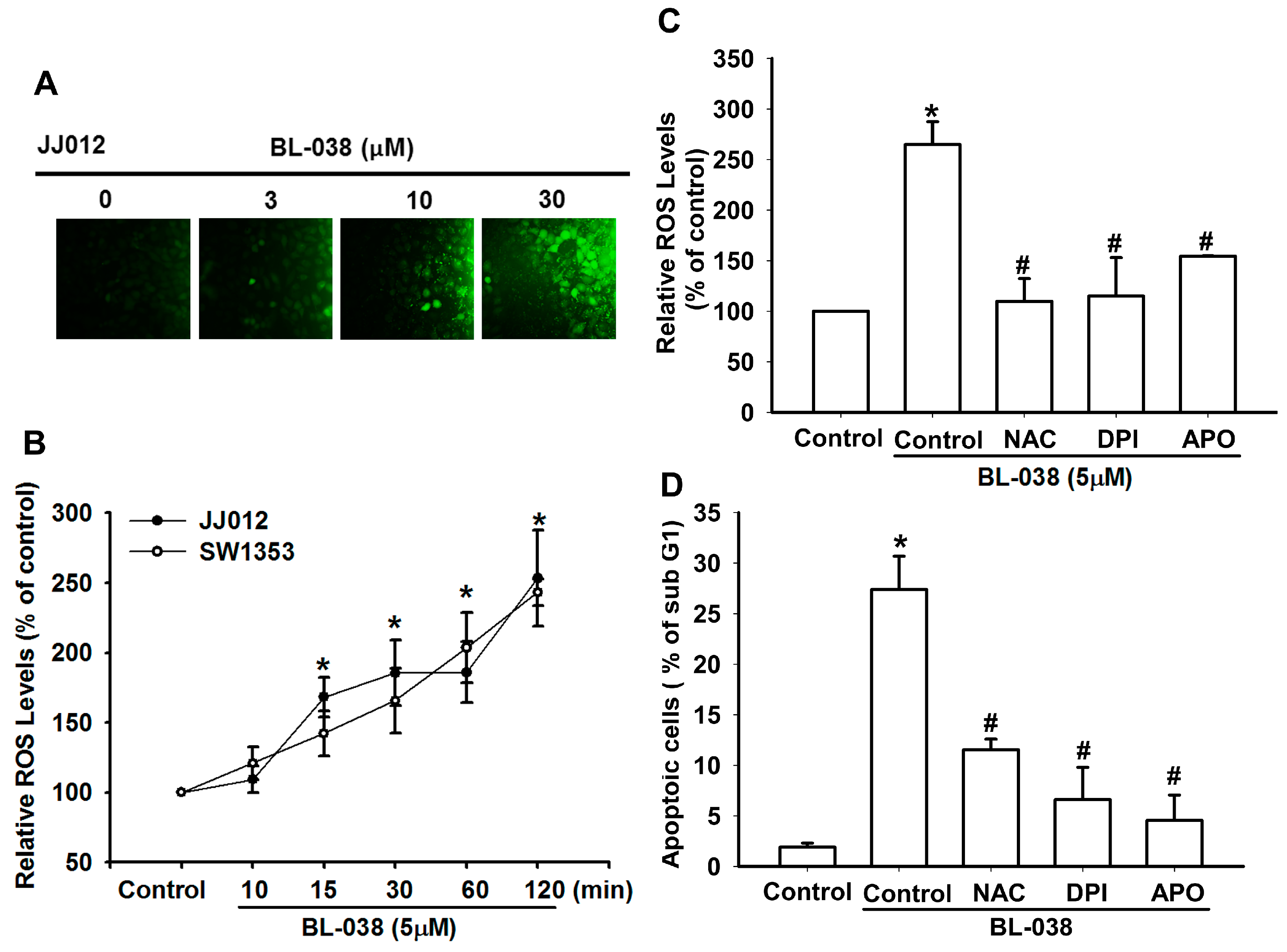
2.3. Reactive Oxygen Species (ROS) Are Involved in BL-038-Induced Apoptosis in Human Chondrosarcoma Cells
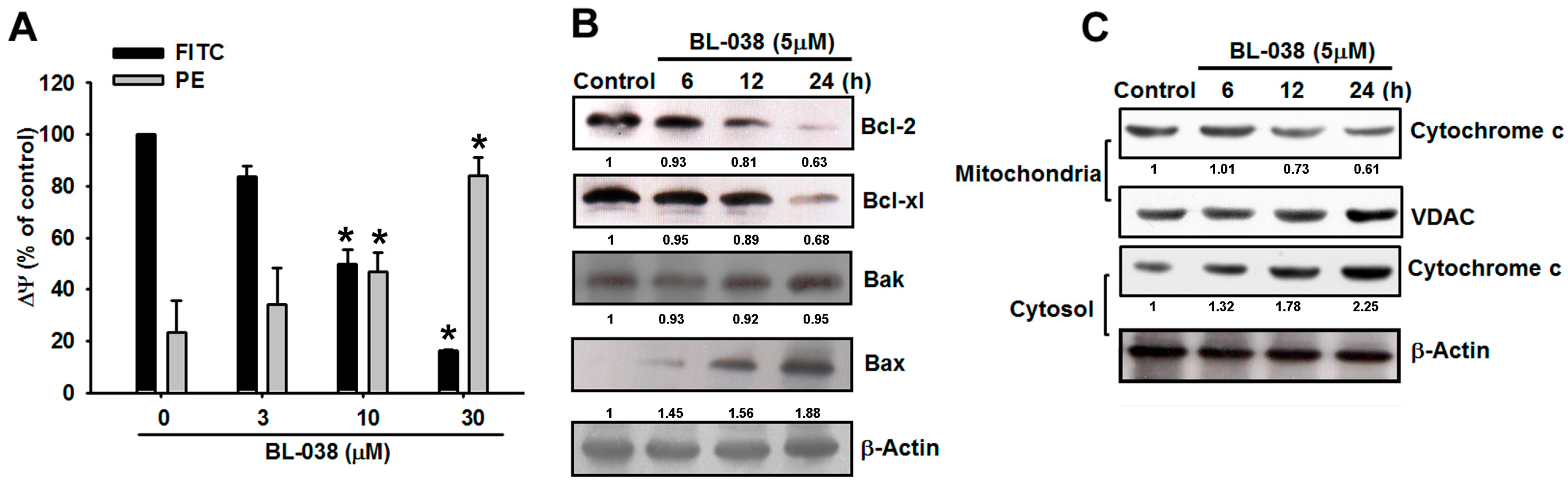
2.4. Involvement of Mitochondrial Dysfunction in BL-038-Induced Human Chondrosarcoma Cell Apoptosis
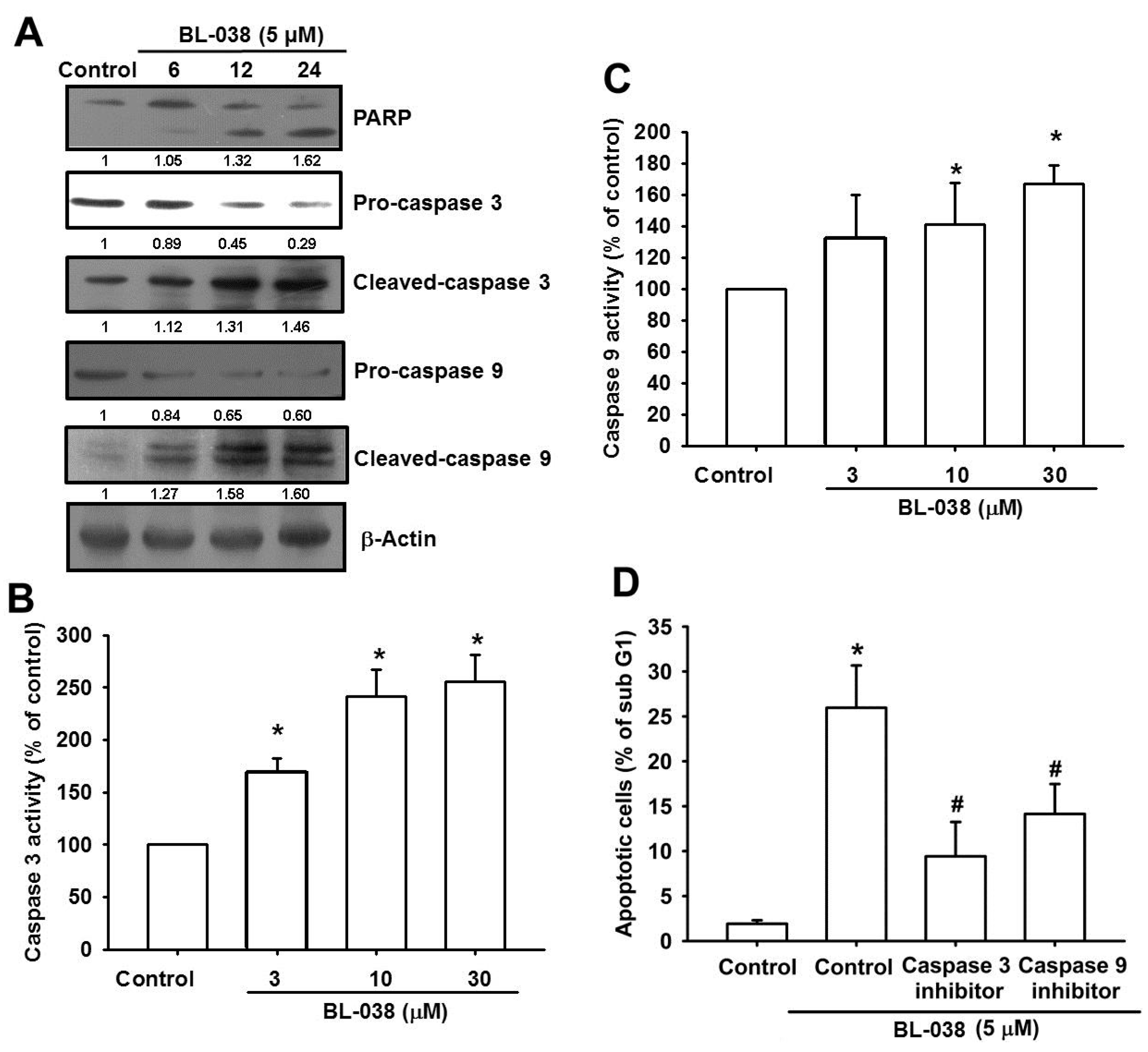
2.5. BL-038 Induces the Activation of Caspases in Human Chondrosarcoma Cells
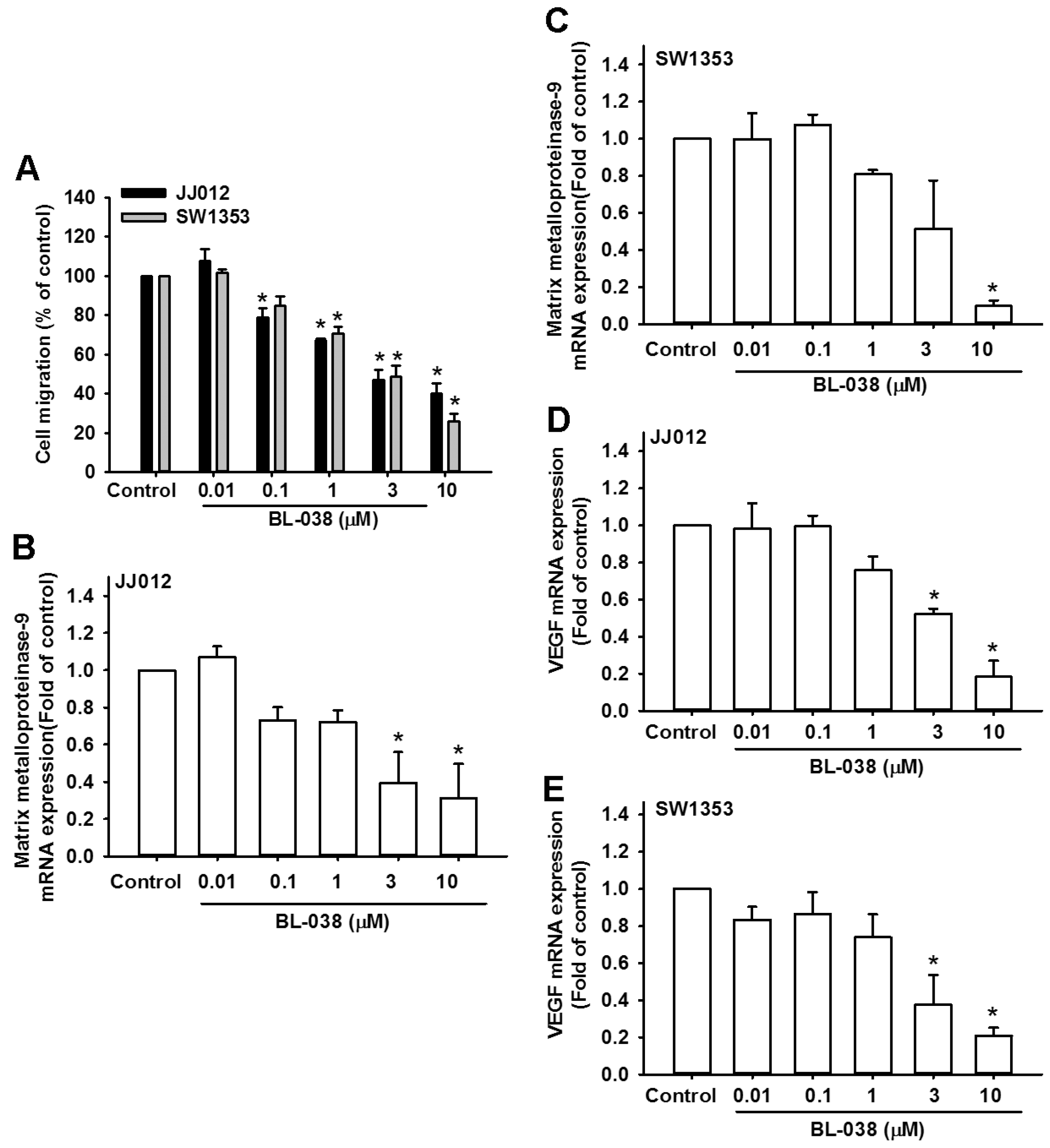
2.6. BL-038 Reduces Cell Migration and Angiogenesis by Decreasing the Expression of Matrix Metalloproteinase-9 and Vascular Endothelial Growth Factor in Human Chondrosarcoma Cells
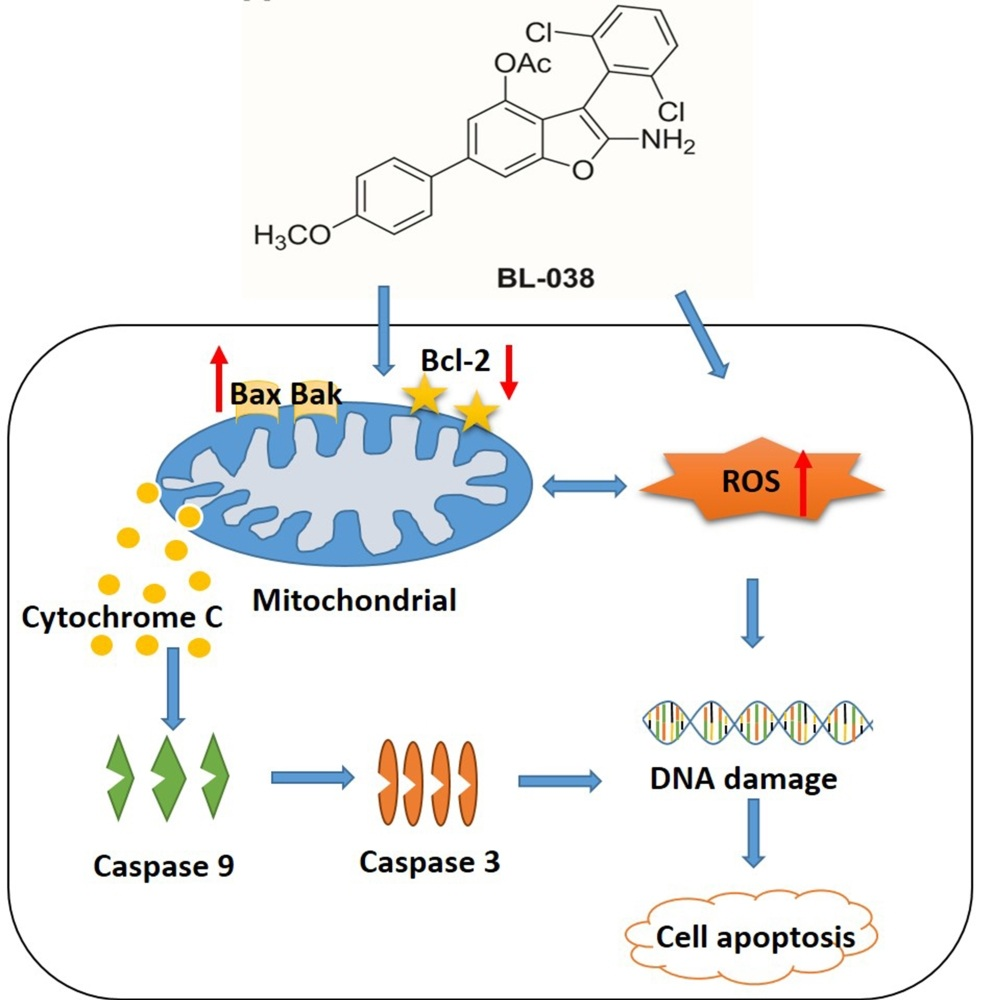
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesized Method of 2-Amino-3-(2,6-dichlorophenyl)-6-(4-methoxyphenyl)benzofuran-4-yl Acetate (BL-038)
4.3. Cell Culture
4.4. MTT Assay
4.5. Colony Formation Assay
4.6. Annexin V/PI Staining
4.7. Cell Cycle Analysis
4.8. TUNEL Staining
4.9. Determination of ROS Generation
4.10. Determination of Mitochondrial Membrane Potential
4.11. Western Blot Analysis
4.12. Caspases Activity Assay
4.13. Quantitative Real-Time PCR (qPCR)
4.14. Migration Assay
4.15. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Logie, C.I.; Walker, E.A.; Forsberg, J.A.; Potter, B.K.; Murphey, M.D. Chondrosarcoma: A diagnostic imager’s guide to decision making and patient management. Semin. Musculoskelet. Radiol. 2013, 17, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Mak, I.W.; Singh, S.; Turcotte, R.; Ghert, M. The epigenetic regulation of SOX9 by Mir-145 in human chondrosarcoma. J. Cell. Biochem. 2015, 116, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Fong, Y.C.; Tang, C.H. Novel strategies for the treatment of chondrosarcomas: Targeting integrins. BioMed Res. Int. 2013, 2013, 396839. [Google Scholar] [CrossRef] [PubMed]
- Boldogh, I.; Bacsi, A.; Choudhury, B.K.; Dharajiya, N.; Alam, R.; Hazra, T.K.; Mitra, S.; Goldblum, R.M.; Sur, S. ROS generated by pollen NADPH oxidase provide a signal that augments antigen-induced allergic airway inflammation. J. Clin. Investig. 2005, 115, 2169–2179. [Google Scholar] [CrossRef] [PubMed]
- Tickner, J.; Fan, L.M.; Du, J.; Meijles, D.; Li, J.M. Nox2-derived ROS in PPARγ signaling and cell-cycle progression of lung alveolar epithelial cells. Free Radic. Biol. Med. 2011, 51, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Hou, C.H.; Lin, F.L.; Tsao, Y.T.; Hou, S.M. Nimbolide induces ROS-regulated apoptosis and inhibits cell migration in osteosarcoma. Int. J. Mol. Sci. 2015, 16, 23405–23424. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Qiu, W.Y.; Pan, Q.; Yao, Y.F.; Xing, K.; Lou, M.F. Reactive oxygen species (ROS) are essential mediators in epidermal growth factor (EGF)-stimulated corneal epithelial cell proliferation, adhesion, migration, and wound healing. Exp. Eye Res. 2009, 89, 876–886. [Google Scholar] [CrossRef] [PubMed]
- You, B.R.; Shin, H.R.; Park, W.H. PX-12 inhibits the growth of A549 lung cancer cells via G2/M phase arrest and ROS-dependent apoptosis. Int. J. Oncol. 2014, 44, 301–308. [Google Scholar] [PubMed]
- Chang, Z.; Xing, J.; Yu, X. Curcumin induces osteosarcoma MG63 cells apoptosis via ROS/Cyto-C/Caspase-3 pathway. Tumour Biol. 2014, 35, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.W.; Kang, S.U.; Choi, J.W.; Shin, Y.S.; Baek, S.J.; Lee, S.H.; Kim, C.H. Tolfenamic acid induces apoptosis and growth inhibition in anaplastic thyroid cancer: Involvement of nonsteroidal anti-inflammatory drug-activated gene-1 expression and intracellular reactive oxygen species generation. Free Radic. Biol. Med. 2014, 67, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Jaattela, M. Cell death induced by TNF or serum starvation is independent of ErbB receptor signaling in MCF-7 breast carcinoma cells. Int. J. Cancer 2000, 86, 617–625. [Google Scholar] [CrossRef]
- Morita, A. Ultraviolet radiation-mediated cellular response. Protein Nucleic Acid Enzym. 1999, 44, 2517–2522. (In Japanese) [Google Scholar]
- Liu, B.; Yuan, B.; Zhang, L.; Mu, W.; Wang, C. ROS/p38/p53/puma signaling pathway is involved in emodin-induced apoptosis of human colorectal cancer cells. Int. J. Clin. Exp. Med. 2015, 8, 15413–15422. [Google Scholar] [PubMed]
- Chang, L.C.; Yu, Y.L. Dietary components as epigenetic-regulating agents against cancer. Biomedicine 2016, 6, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Padma, V.V. An overview of targeted cancer therapy. Biomedicine 2015, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Antiinflammatory activity of the antirheumatic herbal drug, gravel root (Eupatorium purpureum): Further biological activities and constituents. Phytother. Res. 2001, 15, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Pauletti, P.M.; Araujo, A.R.; Young, M.C.; Giesbrecht, A.M.; Bolzani, V.D. nor-Lignans from the leaves of Styrax ferrugineus (styracaceae) with antibacterial and antifungal activity. Phytochemistry 2000, 55, 597–601. [Google Scholar] [CrossRef]
- Masubuchi, M.; Kawasaki, K.; Ebiike, H.; Ikeda, Y.; Tsujii, S.; Sogabe, S.; Fujii, T.; Sakata, K.; Shiratori, Y.; Aoki, Y.; et al. Design and synthesis of novel benzofurans as a new class of antifungal agents targeting fungal N-myristoyltransferase. Part 1. Bioorg. Med. Chem. Lett. 2001, 11, 1833–1837. [Google Scholar] [CrossRef]
- Kayser, O.; Chen, M.; Kharazmi, A.; Kiderlen, A.F. Aurones interfere with leishmania major mitochondrial fumarate reductase. Z. Naturforsch. C Biol. Sci. 2002, 57, 717–720. [Google Scholar] [CrossRef]
- Li, X.Y.; He, B.F.; Luo, H.J.; Huang, N.Y.; Deng, W.Q. 3-Acyl-5-hydroxybenzofuran derivatives as potential anti-estrogen breast cancer agents: A combined experimental and theoretical investigation. Bioorg. Med. Chem. Lett. 2013, 23, 4617–4621. [Google Scholar] [CrossRef] [PubMed]
- Freedman, V.H.; Shin, S.I. Cellular tumorigenicity in nude mice: Correlation with cell growth in semi-solid medium. Cell 1974, 3, 355–359. [Google Scholar] [CrossRef]
- Zou, P.; Xia, Y.; Chen, T.; Zhang, J.; Wang, Z.; Chen, W.; Chen, M.; Kanchana, K.; Yang, S.; Liang, G. Selective killing of gastric cancer cells by a small molecule targeting ROS-mediated er stress activation. Mol. Carcinog. 2016, 55, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Llambi, F. Cell death signaling. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Li, X.; Li, Y.; Li, C.; Gao, B.; Gan, H.; Li, S.; Shen, J.; Kang, J.; Ding, S.; et al. Gallic acid induces apoptosis and inhibits cell migration by upregulating miR-518b in sw1353 human chondrosarcoma cells. Int. J. Oncol. 2014, 44, 91–98. [Google Scholar] [PubMed]
- Girard, N.; Bazille, C.; Lhuissier, E.; Benateau, H.; Llombart-Bosch, A.; Boumediene, K.; Bauge, C. 3-Deazaneplanocin a (DZNep), an inhibitor of the histone methyltransferase EZH2, induces apoptosis and reduces cell migration in chondrosarcoma cells. PLoS ONE 2014, 9, e98176. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Jiang, T.; Guo, F.; Gong, C.; Yang, K.; Wu, Y.; Huang, X.; Cheng, W.; Xu, K. Hedgehog pathway inhibitor-4 suppresses malignant properties of chondrosarcoma cells by disturbing tumor ciliogenesis. Oncol. Rep. 2014, 32, 1622–1630. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.P.; Lin, C.Y.; Shih, J.S.; Fong, Y.C.; Wang, S.W.; Li, T.M.; Tang, C.H. Adiponectin promotes VEGF-A-dependent angiogenesis in human chondrosarcoma through PI3K, Akt, mTOR, and HIF-α pathway. Oncotarget 2015, 6, 36746–36761. [Google Scholar] [PubMed]
- Wu, M.H.; Huang, C.Y.; Lin, J.A.; Wang, S.W.; Peng, C.Y.; Cheng, H.C.; Tang, C.H. Endothelin-1 promotes vascular endothelial growth factor-dependent angiogenesis in human chondrosarcoma cells. Oncogene 2014, 33, 1725–1735. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Charbonneau, C.; Wei, L.; Yang, W.; Chen, Q.; Terek, R.M. CXCR4-targeted therapy inhibits VEGF expression and chondrosarcoma angiogenesis and metastasis. Mol. Cancer Ther. 2013, 12, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zeng, G.; Li, X.; Wu, Z.; Wang, L. Cantharidin inhibits cell proliferation and promotes apoptosis in tongue squamous cell carcinoma through suppression of miR-214 and regulation of p53 and Bcl-2/bax. Oncol. Rep. 2015, 33, 3061–3068. [Google Scholar] [CrossRef] [PubMed]
- Su, C.C.; Liu, S.H.; Lee, K.I.; Huang, K.T.; Lu, T.H.; Fang, K.M.; Wu, C.C.; Yen, C.C.; Lai, C.H.; Su, Y.C.; et al. Cantharidin induces apoptosis through the calcium/PKC-regulated endoplasmic reticulum stress pathway in human bladder cancer cells. Am. J. Chin. Med. 2015, 43, 581–600. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, A.; Asadi, J.; Rostami-Charati, F.; Gharaei, R. High cytotoxicity and apoptotic effects of natural bioactive benzofuran derivative on the MCF-7 breast cancer cell line. Comb. Chem. High Throughput Screen. 2015, 18, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.; Ghert, M.A.; Harrelson, J.M.; Scully, S.P. Chondrosarcoma of bone: Analysis of 108 cases and evaluation for predictors of outcome. Clin. Orthop. Relat. Res. 2001, 391, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Gitelis, S.; Bertoni, F.; Picci, P.; Campanacci, M. Chondrosarcoma of bone. The experience at the istituto ortopedico rizzoli. J. Bone Jt. Surg. Am. 1981, 63, 1248–1257. [Google Scholar]
- Chen, M.C.; Hsu, S.L.; Lin, H.; Yang, T.Y. Retinoic acid and cancer treatment. Biomedicine 2014, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Silva, A.M.; Matos, M.; Monteiro, S.M.; Alvaro, A.R. Copper induced apoptosis in Caco-2 and Hep-G2 cells: Expression of caspases 3, 8 and 9, AIF and p53. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2016, 185, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Indran, I.R.; Tufo, G.; Pervaiz, S.; Brenner, C. Recent advances in apoptosis, mitochondria and drug resistance in cancer cells. Biochim. Biophys. Acta 2011, 1807, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. Bcl-2 family members and the mitochondria in apoptosis. Genes Dev. 1999, 13, 1899–1911. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.W.; Puthalakath, H. Bcl-2 family proteins: The sentinels of the mitochondrial apoptosis pathway. IUBMB Life 2008, 60, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhou, T.; Tao, Y.; Wang, Q.; Liu, P.; Liu, C. Salvianolic acid b attenuates hepatocyte apoptosis by regulating mediators in death receptor and mitochondrial pathways. Exp. Biol. Med. 2010, 235, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hwang, B.Y.; Su, B.N.; Chai, H.; Mi, Q.; Kinghorn, A.D.; Wild, R.; Swanson, S.M. Silvestrol, a potential anticancer rocaglate derivative from aglaia foveolata, induces apoptosis in lncap cells through the mitochondrial/apoptosome pathway without activation of executioner caspase-3 or -7. Anticancer Res. 2007, 27, 2175–2183. [Google Scholar] [PubMed]
- Felser, A.; Lindinger, P.W.; Schnell, D.; Kratschmar, D.V.; Odermatt, A.; Mies, S.; Jeno, P.; Krahenbuhl, S. Hepatocellular toxicity of benzbromarone: Effects on mitochondrial function and structure. Toxicology 2014, 324, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Oyinloye, B.E.; Adenowo, A.F.; Kappo, A.P. Reactive oxygen species, apoptosis, antimicrobial peptides and human inflammatory diseases. Pharmaceuticals 2015, 8, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Lee, Y.H.; Wang, Y.J. ROS-triggered signaling pathways involved in the cytotoxicity and tumor promotion effects of pentachlorophenol and tetrachlorohydroquinone. Chem. Res. Toxicol. 2015, 28, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.M.; Shen, M.; Zhou, Z.Y.; Pan, P.; Hoi, P.M.; Li, S.; Liang, W.; Ai, N.; Zhang, L.Q.; Li, C.W.; et al. Discovery of a benzofuran derivative (MBPTA) as a novel ROCK inhibitor that protects against MPP+-induced oxidative stress and cell death in SH-SY5Y cells. Free Radic. Biol. Med. 2014, 74, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, P.K.; Srivastava, S.; Gupta, J.; Thakur, I.S. Dibenzofuran induces oxidative stress, disruption of trans-mitochondrial membrane potential (Δψm) and G1 arrest in human hepatoma cell line. Toxicol. Lett. 2012, 214, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef]
- Merdad, A.; Karim, S.; Schulten, H.J.; Dallol, A.; Buhmeida, A.; Al-Thubaity, F.; Gari, M.A.; Chaudhary, A.G.; Abuzenadah, A.M.; Al-Qahtani, M.H. Expression of matrix metalloproteinases (MMPs) in primary human breast cancer: MMP-9 as a potential biomarker for cancer invasion and metastasis. Anticancer Res. 2014, 34, 1355–1366. [Google Scholar] [PubMed]
- Herrlinger, U.; Wiendl, H.; Renninger, M.; Forschler, H.; Dichgans, J.; Weller, M. Vascular endothelial growth factor (VEGF) in leptomeningeal metastasis: Diagnostic and prognostic value. Br. J. Cancer 2004, 91, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, S.I.; Arii, S.; Furutani, M.; Niwano, M.; Harada, T.; Mizumoto, M.; Mori, A.; Onodera, H.; Imamura, M. Predictive value of vascular endothelial growth factor (VEGF) in metastasis and prognosis of human colorectal cancer. Br. J. Cancer 1998, 78, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Arii, S.; Ishigami, S.; Mori, A.; Onodera, H.; Imamura, M. Implication of VEGF and mmps in hepatic metastasis of human colon cancer. Nihon Geka Gakkai Zasshi 1998, 99, 436–440. (In Japanese) [Google Scholar] [PubMed]
- Sung, L.Y.; Lo, W.H.; Chiu, H.Y.; Chen, H.C.; Chung, C.K.; Lee, H.P.; Hu, Y.C. Modulation of chondrocyte phenotype via baculovirus-mediated growth factor expression. Biomaterials 2007, 28, 3437–3447. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lv, L.; Ling, Z.; Wang, Y.; Liu, Y.; Li, L.; Liu, G.; Shen, L.; Yan, J.; Wang, Y. Long-circulating iodinated albumin-gadolinium nanoparticles as enhanced magnetic resonance and computed tomography imaging probes for osteosarcoma visualization. Anal. Chem. 2015, 87, 4299–4304. [Google Scholar] [CrossRef] [PubMed]
- Dijkers, P.F.; Birkenkamp, K.U.; Lam, E.W.; Thomas, N.S.; Lammers, J.W.; Koenderman, L.; Coffer, P.J. Fkhr-L1 can act as a critical effector of cell death induced by cytokine withdrawal: Protein kinase B-enhanced cell survival through maintenance of mitochondrial integrity. J. Cell Biol. 2002, 156, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.T.; Fong, Y.C.; Li, T.M.; Liu, J.F.; Hsu, C.W.; Chang, C.S.; Tang, C.H. DDTD, an isoflavone derivative, induces cell apoptosis through the reactive oxygen species/apoptosis signal-regulating kinase 1 pathway in human osteosarcoma cells. Eur. J. Pharmacol. 2008, 597, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Yang, W.H.; Fong, Y.C.; Kuo, S.C.; Chang, C.S.; Tang, C.H. BFPP, a phloroglucinol derivative, induces cell apoptosis in human chondrosarcoma cells through endoplasmic reticulum stress. Biochem. Pharmacol. 2010, 79, 1410–1417. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Fong, Y.C.; Chang, K.W.; Kuo, S.C.; Chang, C.S.; Tang, C.H. FPTB, a novel CA-4 derivative, induces cell apoptosis of human chondrosarcoma cells through mitochondrial dysfunction and endoplasmic reticulum stress pathways. J. Cell. Biochem. 2011, 112, 453–462. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.-F.; Chen, C.-Y.; Chen, H.-T.; Chang, C.-S.; Tang, C.-H. BL-038, a Benzofuran Derivative, Induces Cell Apoptosis in Human Chondrosarcoma Cells through Reactive Oxygen Species/Mitochondrial Dysfunction and the Caspases Dependent Pathway. Int. J. Mol. Sci. 2016, 17, 1491. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091491
Liu J-F, Chen C-Y, Chen H-T, Chang C-S, Tang C-H. BL-038, a Benzofuran Derivative, Induces Cell Apoptosis in Human Chondrosarcoma Cells through Reactive Oxygen Species/Mitochondrial Dysfunction and the Caspases Dependent Pathway. International Journal of Molecular Sciences. 2016; 17(9):1491. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091491
Chicago/Turabian StyleLiu, Ju-Fang, Chien-Yu Chen, Hsien-Te Chen, Chih-Shiang Chang, and Chih-Hsin Tang. 2016. "BL-038, a Benzofuran Derivative, Induces Cell Apoptosis in Human Chondrosarcoma Cells through Reactive Oxygen Species/Mitochondrial Dysfunction and the Caspases Dependent Pathway" International Journal of Molecular Sciences 17, no. 9: 1491. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091491