
Role of n-3 Polyunsaturated Fatty Acids in Ameliorating the Obesity-Induced Metabolic Syndrome in Animal Models and Humans

 ,
,
Abstract
:1. Introduction
2. Methodology of Reference Selection
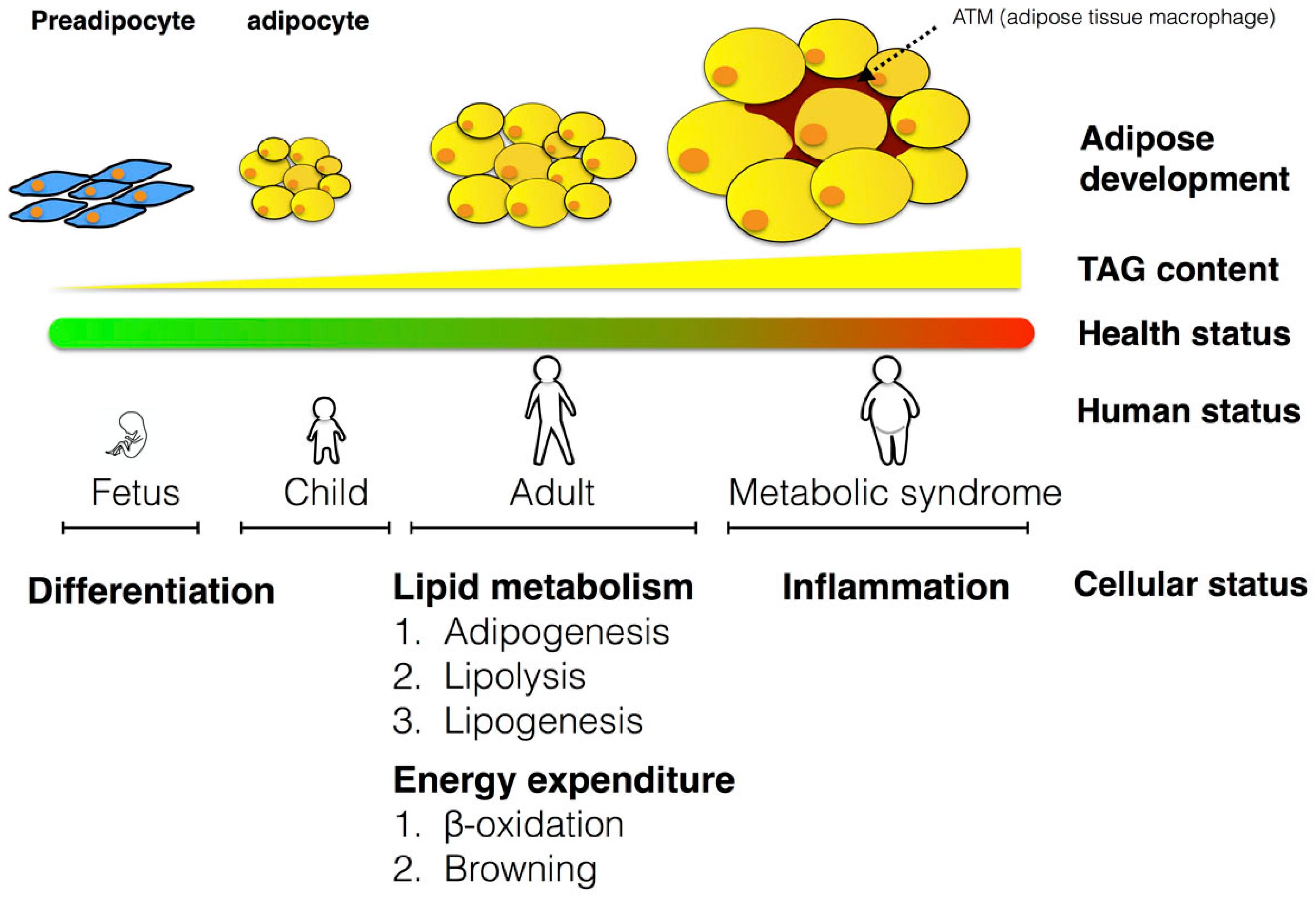
3. The Role of Adipose Tissues in Metabolism
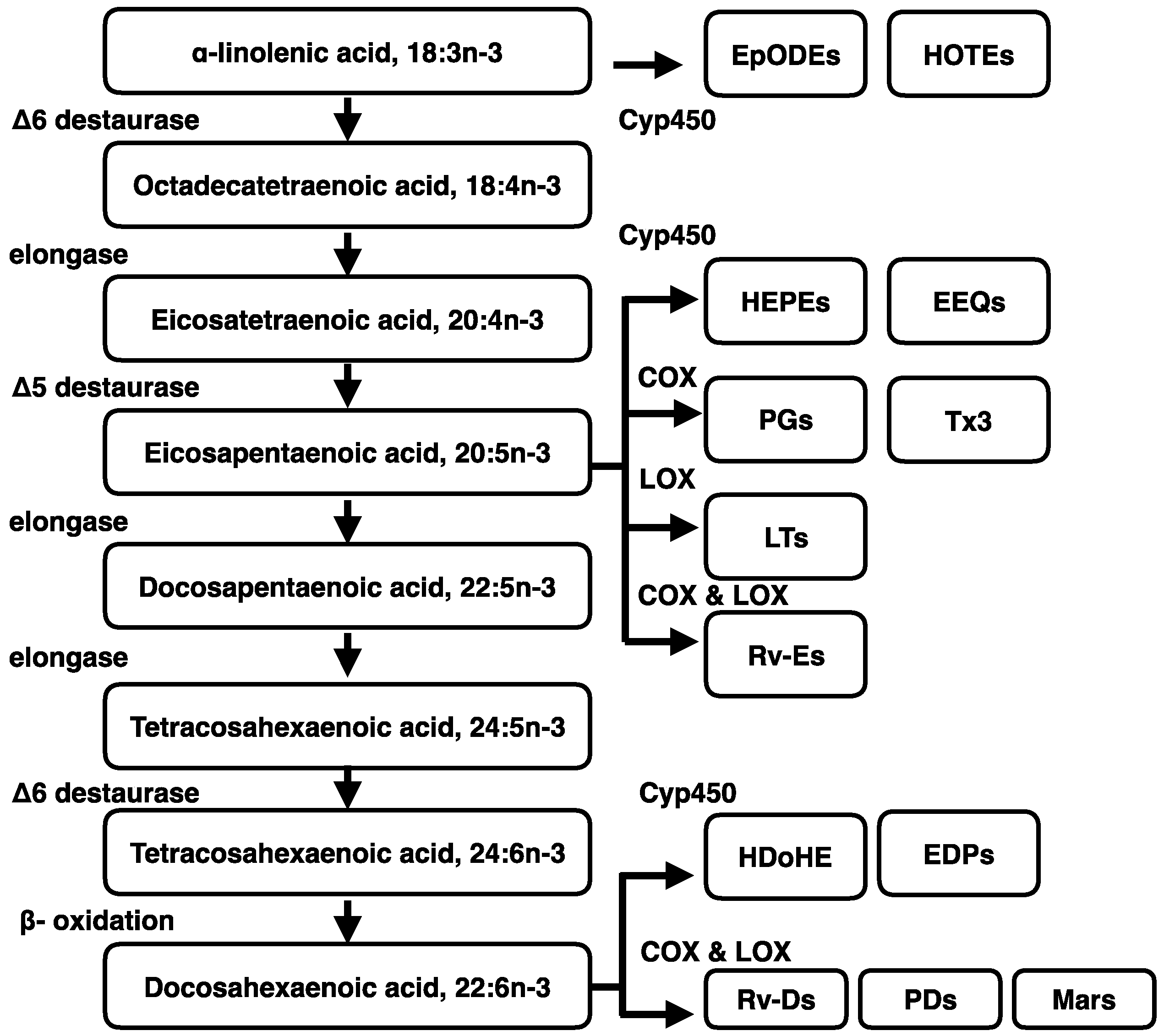
4. Metabolism of Dietary n-3 Polyunsaturated Fatty Acids (n-3 PUFA)
5. The Beneficial Physiological Effects of n-3 PUFA
5.1. The Beneficial Effect of n-3 PUFA in Human Studies
5.1.1. The Effects of n-3 PUFA in Different Periods
5.1.2. The Effects of n-3 PUFA on Inflammation Factors in Humans
5.1.3. The Receptor for n-3 PUFA
5.1.4. Confounding Factors May Mask the Effect of n-3 PUFA in Human Studies
5.2. The Benefits of n-3 PUFA in Animal Studies
5.2.1. The Effect of Endogenous n-3 PUFA in Animals
5.2.2. The Effect of Exogenous n-3 PUFA in Animals
5.2.3. Potential Mechanisms Mediate n-3 PUFA Effects
5.2.4. Suitable Animal Models for Dietary Intervention
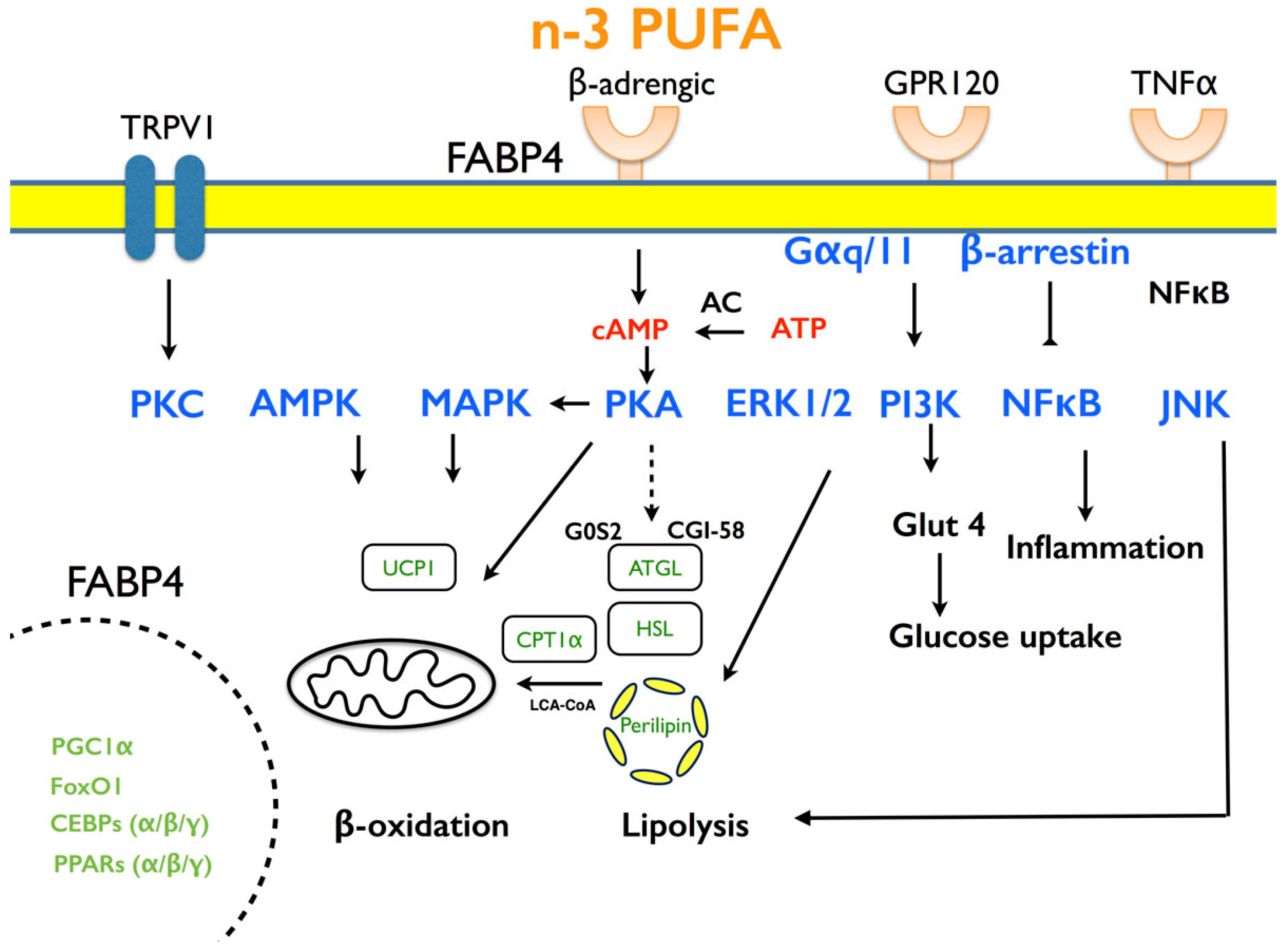
6. The Molecular Mechanisms by which PUFA Affects Lipid Metabolism
6.1. Adipogenesis
6.2. Lipid Accumulation
6.3. Energy Expenditure
6.4. Inflammation
7. The Interplay between Liver and Adipose Tissues
7.1. Lipotoxicity in Livers Regulated by n-3 PUFA
7.2. Hepatokines Regulated by n-3 PUFA
7.2.1. Serum Amyloid A
7.2.2. Fibroblast Growth Factor 21
7.2.3. Angiopoietin-Like 4
8. Conclusions and Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grundy, S.M. Obesity, metabolic syndrome, and cardiovascular disease. J. Clin. Endocrinol. Metab. 2004, 89, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Spite, M.; Clària, J.; Serhan, C.N. Resolvins, specialized proresolving lipid mediators, and their potential roles in metabolic diseases. Cell Metab. 2014, 19, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.-C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the metabolic syndrome a joint interim statement of the international diabetes federation task force on epidemiology and prevention; national heart, lung, and blood institute; American heart association; world heart federation; international atherosclerosis society; and international association for the study of obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [PubMed]
- Clarke, S.D. Polyunsaturated fatty acid regulation of gene transcription: A mechanism to improve energy balance and insulin resistance. Br. J. Nutr. 2000, 83, S59–S66. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. An increase in the ω-6/ω-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Evolutionary aspects of diet and essential fatty acids. World Rev. Nutr. Diet. 2001, 88, 18–27. [Google Scholar] [PubMed]
- Donahue, S.M.; Rifas-Shiman, S.L.; Gold, D.R.; Jouni, Z.E.; Gillman, M.W.; Oken, E. Prenatal fatty acid status and child adiposity at age 3 y: Results from a US pregnancy cohort. Am. J. Clin. Nutr. 2011, 93, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Conner, W.; Lin, D.; Colvis, C. Differential mobilization of fatty acids from adipose tissue. J. Lipid Res. 1996, 37, 290–298. [Google Scholar] [PubMed]
- Herzberg, G.R.; Skinner, C. Differential accumulation and release of long-chain n-3 fatty acids from liver, muscle, and adipose tissue triacylglycerols. Can. J. Physiol. Pharmacol. 1997, 75, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Strawford, A.; Antelo, F.; Christiansen, M.; Hellerstein, M. Adipose tissue triglyceride turnover, de novo lipogenesis, and cell proliferation in humans measured with 2H2O. Am. J. Physiol.-Endocrinol. Metab. 2004, 286, E577–E588. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef] [PubMed]
- Després, J.-P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Lafontan, M.; Girard, J. Impact of visceral adipose tissue on liver metabolism: Part I: Heterogeneity of adipose tissue and functional properties of visceral adipose tissue. Diabetes Metab. 2008, 34, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Cusi, K. The role of adipose tissue and lipotoxicity in the pathogenesis of type 2 diabetes. Curr. Diabetes Rep. 2010, 10, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Wree, A.; Kahraman, A.; Gerken, G.; Canbay, A. Obesity affects the liver—The link between adipocytes and hepatocytes. Digestion 2011, 83, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.; Dyerberg, J.; Hjørne, N. The composition of food consumed by Greenland Eskimos. Acta Med. Scand. 1976, 200, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Connor, W.E. Importance of n-3 fatty acids in health and disease. Am. J. Clin. Nutr. 2000, 71, 171S–175S. [Google Scholar] [PubMed]
- Lorente-Cebrián, S.; Costa, A.G.; Navas-Carretero, S.; Zabala, M.; Martínez, J.A.; Moreno-Aliaga, M.J. Role of ω-3 fatty acids in obesity, metabolic syndrome, and cardiovascular diseases: A review of the evidence. J. Physiol. Biochem. 2013, 69, 633–651. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T. Efficiency of conversion of α-linolenic acid to long chain n-3 fatty acids in man. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Conversion of α-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, M.; Ma, K.; Chang, L.; Bell, J.M.; Rapoport, S.I.; DeMar, J.C. Low liver conversion rate of α-linolenic to docosahexaenoic acid in awake rats on a high-docosahexaenoate-containing diet. J. Lipid Res. 2006, 47, 1812–1822. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, M.; DeMar, J.C.; Ma, K.; Chang, L.; Bell, J.M.; Rapoport, S.I. Docosahexaenoic acid synthesis from α-linolenic acid by rat brain is unaffected by dietary n-3 PUFA deprivation. J. Lipid Res. 2007, 48, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Panchal, S.K.; Diwan, V.; Brown, L. ω-3 fatty acids and metabolic syndrome: Effects and emerging mechanisms of action. Prog. Lipid Res. 2011, 50, 372–387. [Google Scholar] [CrossRef] [PubMed]
- Doughman, S.; Ryan, A.; Krupanidhi, S.; Sanjeevi, C.; Mohan, V. High DHA dosage from algae oil improves postprandial hypertriglyceridemia and is safe for type-2 diabetics. Int. J. Diabetes Dev. Ctries 2013, 33, 75–82. [Google Scholar] [CrossRef]
- Jump, D.B. n-3 polyunsaturated fatty acid regulation of hepatic gene transcription. Curr. Opin. Lipidol. 2008, 19, 242. [Google Scholar] [CrossRef] [PubMed]
- Polozova, A.; Salem, N., Jr. Role of liver and plasma lipoproteins in selective transport of n-3 fatty acids to tissues: A comparative study of 14C-DHA and 3H-oleic acid tracers. J. Mol. Neurosci. 2007, 33, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Al-Hasani, H.; Joost, H.-G. Nutrition-/diet-induced changes in gene expression in white adipose tissue. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, L.; Laiglesia, L.M.; Huerta, A.E.; Martínez, J.A.; Moreno-Aliaga, M.J. ω-3 fatty acids and adipose tissue function in obesity and metabolic syndrome. Prostaglandins Lipid Mediat. 2015, 121, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Todorčević, M.; Hodson, L. The Effect of Marine Derived n-3 Fatty Acids on Adipose Tissue Metabolism and Function. J. Clin. Med. 2016, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.D.; Howe, P.R. Long-chain ω-3 polyunsaturated fatty acids may be beneficial for reducing obesity—A review. Nutrients 2010, 2, 1212–1230. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Kim, W.K.; Bae, K.-H. Distinction of white, beige and brown adipocytes derived from mesenchymal stem cells. World J. Stem Cells 2014, 6, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Berry, R.; Jeffery, E.; Rodeheffer, M.S. Weighing in on adipocyte precursors. Cell Metab. 2014, 19, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, P.R.; Gnudi, L.; Tozzo, E.; Yang, H.; Leach, F.; Kahn, B.B. Adipose cell hyperplasia and enhanced glucose disposal in transgenic mice overexpressing GLUT4 selectively in adipose tissue. J. Biol. Chem. 1993, 268, 22243–22246. [Google Scholar] [PubMed]
- Rydén, M.; Andersson, D.P.; Bernard, S.; Spalding, K.; Arner, P. Adipocyte triglyceride turnover and lipolysis in lean and overweight subjects. J. Lipid Res. 2013, 54, 2909–2913. [Google Scholar] [CrossRef] [PubMed]
- Magkos, F.; Fraterrigo, G.; Yoshino, J.; Luecking, C.; Kirbach, K.; Kelly, S.C.; de las Fuentes, L.; He, S.; Okunade, A.L.; Patterson, B.W.; et al. Effects of Moderate and Subsequent Progressive Weight Loss on Metabolic Function and Adipose Tissue Biology in Humans with Obesity. Cell Metab. 2016. [Google Scholar] [CrossRef] [PubMed]
- Van der Lans, A.A.; Hoeks, J.; Brans, B.; Vijgen, G.H.; Visser, M.G.; Vosselman, M.J.; Hansen, J.; Jorgensen, J.A.; Wu, J.; Mottaghy, F.M.; et al. Cold acclimation recruits human brown fat and increases nonshivering thermogenesis. J. Clin. Investig. 2013, 123, 3395–3403. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bostrom, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Palou, A.; Picó, C.; Bonet, M.L.; Oliver, P. The uncoupling protein, thermogenin. Int. J. Biochem. Cell Biol. 1998, 30, 7–11. [Google Scholar] [CrossRef]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.-H.; Doria, A. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Li, L. Browning of white adipose tissue: Role of hypothalamic signaling. Ann. N. Y. Acad. Sci. 2013, 1302, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Baboota, R.K.; Singh, D.P.; Sarma, S.M.; Kaur, J.; Sandhir, R.; Boparai, R.K.; Kondepudi, K.K.; Bishnoi, M. Capsaicin induces “brite” phenotype in differentiating 3T3-L1 preadipocytes. PLoS ONE 2014, 9, e103093. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Goto, T.; Yu, R.; Uchida, K.; Tominaga, M.; Kano, Y.; Takahashi, N.; Kawada, T. Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Choi, J.H.; Mukherjee, R.; Hwang, K.-C.; Yun, J.W. Proteomic identification of fat-browning markers in cultured white adipocytes treated with curcumin. Mol. Cell. Biochem. 2016. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic peroxisome proliferator-activated receptor γ (PPARγ) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, W.N.; Galli, C. Fat and fatty acid terminology, methods of analysis and fat digestion and metabolism: A background review paper. Ann. Nutr. Metab. 2009, 55, 8–43. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B. The biochemistry of n-3 polyunsaturated fatty acids. J. Biol. Chem. 2002, 277, 8755–8758. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, R.P.; Layé, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.L.; Bazan, N.G. Membrane docosahexaenoate is supplied to the developing brain and retina by the liver. Proc. Natl. Acad. Sci. USA 1989, 86, 2903–2907. [Google Scholar] [CrossRef] [PubMed]
- Domenichiello, A.F.; Kitson, A.P.; Bazinet, R.P. Is docosahexaenoic acid synthesis from α-linolenic acid sufficient to supply the adult brain? Prog. Lipid Res. 2015, 59, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Anderson, M.J. The majority of dietary linoleate in growing rats is β-oxidized or stored in visceral fat. J. Nutr. 1997, 127, 146–152. [Google Scholar] [PubMed]
- Goyens, P.L.; Spilker, M.E.; Zock, P.L.; Katan, M.B.; Mensink, R.P. Compartmental modeling to quantify α-linolenic acid conversion after longer term intake of multiple tracer boluses. J. Lipid Res. 2005, 46, 1474–1483. [Google Scholar] [CrossRef] [PubMed]
- Hussein, N.; Ah-Sing, E.; Wilkinson, P.; Leach, C.; Griffin, B.A.; Millward, D.J. Long-chain conversion of [13C] linoleic acid and α-linolenic acid in response to marked changes in their dietary intake in men. J. Lipid Res. 2005, 46, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Salem, N.; Sinclair, A.J.; Cunnane, S.C. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467S–1476S. [Google Scholar] [PubMed]
- Qin, X.; Park, H.G.; Zhang, J.Y.; Lawrence, P.; Liu, G.; Subramanian, N.; Kothapalli, K.S.; Brenna, J.T. Brown but not white adipose cells synthesize ω-3 docosahexaenoic acid in culture. Prostaglandins Leukot. Essent. Fat. Acids 2016, 104, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, P.; Sierra-Johnson, J.; Gertow, K.; Rosell, M.; Vessby, B.; de Faire, U.; Hamsten, A.; Hellenius, M.-L.; Fisher, R. Fatty acid desaturases in human adipose tissue: Relationships between gene expression, desaturation indexes and insulin resistance. Diabetologia 2008, 51, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Truong, H.; DiBello, J.R.; Ruiz-Narvaez, E.; Kraft, P.; Campos, H.; Baylin, A. Does genetic variation in the Δ6-desaturase promoter modify the association between α-linolenic acid and the prevalence of metabolic syndrome? Am. J. Clin. Nutr. 2009, 89, 920–925. [Google Scholar] [CrossRef] [PubMed]
- Levy, B.D. Resolvins and protectins: Natural pharmacophores for resolution biology. Prostaglandins Leukot. Essent. Fat. Acids 2010, 82, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Brain, S.D.; Buckley, C.D.; Gilroy, D.W.; Haslett, C.; O’Neill, L.A.; Perretti, M.; Rossi, A.G.; Wallace, J.L. Resolution of inflammation: State of the art, definitions and terms. FASEB J. 2007, 21, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Mas, E.; Croft, K.D.; Zahra, P.; Barden, A.; Mori, T.A. Resolvins D1, D2, and other mediators of self-limited resolution of inflammation in human blood following n-3 fatty acid supplementation. Clin. Chem. 2012, 58, 1476–1484. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Mitchell, P.L.; Schwab, M.; Trottier, J.; Kang, J.X.; Barbier, O.; Marette, A. Transgenic ω-3 PUFA enrichment alters morphology and gene expression profile in adipose tissue of obese mice: Potential role for protectins. Metabolism 2015, 64, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.; Tang, Y.; Kosuri, M.; Bhatnagar, A.; Spite, M. Resolvin D1 decreases adipose tissue macrophage accumulation and improves insulin sensitivity in obese-diabetic mice. FASEB J. 2011, 25, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Richardson, K.; Louie-Gao, Q.; Arnett, D.K.; Parnell, L.D.; Lai, C.-Q.; Davalos, A.; Fox, C.S.; Demissie, S.; Cupples, L.A.; Fernandez-Hernando, C.; et al. The PLIN4 variant rs8887 modulates obesity related phenotypes in humans through creation of a novel miR-522 seed site. PLoS ONE 2011, 6, e17944. [Google Scholar] [CrossRef] [PubMed]
- Donahue, S.; Rifas-Shiman, S.; Olsen, S.; Gold, D.; Gillman, M.; Oken, E. Associations of maternal prenatal dietary intake of n-3 and n-6 fatty acids with maternal and umbilical cord blood levels. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Scaglioni, S.; Verduci, E.; Salvioni, M.; Bruzzese, M.G.; Radaelli, G.; Zetterström, R.; Riva, E.; Agostoni, C. Plasma long-chain fatty acids and the degree of obesity in Italian children. Acta Paediatr. 2006, 95, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Verduci, E.; Scaglioni, S.; Agostoni, C.; Radaelli, G.; Biondi, M.; Manso, A.S.; Riva, E.; Giovannini, M. The relationship of insulin resistance with SNP 276G>T at adiponectin gene and plasma long-chain polyunsaturated fatty acids in obese children. Pediatr. Res. 2009, 66, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Micallef, M.; Munro, I.; Phang, M.; Garg, M. Plasma n-3 polyunsaturated fatty acids are negatively associated with obesity. Br. J. Nutr. 2009, 102, 1370–1374. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.R.; Edwards, M.J.; DeFilipis, A.P.; Jacobson, T.A. Flaxseed oil increases the plasma concentrations of cardioprotective (n-3) fatty acids in humans. J. Nutr. 2006, 136, 83–87. [Google Scholar] [PubMed]
- Kunesova, M.; Braunerova, R.; Hlavatý, P.; Tvrzicka, E. The influence of n-3 polyunsaturated fatty acids and very low calorie diet during a short-term weight reducing regimen on weight loss and serum fatty acid composition in severely obese women. Physiol. Res. 2006, 55, 63. [Google Scholar] [PubMed]
- Agren, J.; Hänninen, O.; Julkunen, A.; Fogelholm, L.; Vidgren, H.; Schwab, U.; Pynnönen, O.; Uusitupa, M. Fish diet, fish oil and docosahexaenoic acid rich oil lower fasting and postprandial plasma lipid levels. Eur. J. Clin. Nutr. 1996, 50, 765–771. [Google Scholar] [PubMed]
- Blonk, M.C.; Bilo, H.; Nauta, J.; Popp-Snijders, C.; Mulder, C.; Donker, A. Dose-response effects of fish-oil supplementation in healthy volunteers. Am. J. Clin. Nutr. 1990, 52, 120–127. [Google Scholar] [PubMed]
- Kelley, D.S.; Siegel, D.; Fedor, D.M.; Adkins, Y.; Mackey, B.E. DHA supplementation decreases serum C-reactive protein and other markers of inflammation in hypertriglyceridemic men. J. Nutr. 2009, 139, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Neff, L.M.; Culiner, J.; Cunningham-Rundles, S.; Seidman, C.; Meehan, D.; Maturi, J.; Wittkowski, K.M.; Levine, B.; Breslow, J.L. Algal docosahexaenoic acid affects plasma lipoprotein particle size distribution in overweight and obese adults. J. Nutr. 2011, 141, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Ghayour-Mobarhan, M.; Rezaiean, S.; Hoseini, M.; Parizade, S.M.R.; Farhoudi, F.; Hosseininezhad, S.J.; Tavallaei, S.; Vejdani, A.; Azimi-Nezhad, M. ω-3 fatty acid supplements improve the cardiovascular risk profile of subjects with metabolic syndrome, including markers of inflammation and auto-immunity. Acta Cardiol. 2009, 64, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Dewell, A.; Marvasti, F.F.; Harris, W.S.; Tsao, P.; Gardner, C.D. Low-and high-dose plant and marine (n-3) fatty acids do not affect plasma inflammatory markers in adults with metabolic syndrome. J. Νutr. 2011, 141, 2166–2171. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.; Browning, L.; McLean, N.; Rothwell, J.; Mishra, G.; Moore, C.; Jebb, S. Additive benefits of long-chain n-3 polyunsaturated fatty acids and weight-loss in the management of cardiovascular disease risk in overweight hyperinsulinaemic women. Int. J. Obes. 2006, 30, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Poissonnet, C.M.; Burdi, A.R.; Bookstein, F.L. Growth and development of human adipose tissue during early gestation. Early Hum. Dev. 1983, 8, 1–11. [Google Scholar] [CrossRef]
- Muhlhausler, B.S.; Duffield, J.A.; Mcmillen, I.C. Increased maternal nutrition stimulates peroxisome proliferator activated receptor-γ, adiponectin, and leptin messenger ribonucleic acid expression in adipose tissue before birth. Endocrinology 2007, 148, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Ogden, C.L.; Carroll, M.D.; Kit, B.K.; Flegal, K.M. Prevalence of obesity and trends in body mass index among US children and adolescents, 1999–2010. J. Am. Med. Assoc. 2012, 307, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Ailhaud, G.; Massiera, F.; Weill, P.; Legrand, P.; Alessandri, J.-M.; Guesnet, P. Temporal changes in dietary fats: role of n-6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity. Prog. Lipid Res. 2006, 45, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Knittle, J.; Timmers, K.; Ginsberg-Fellner, F.; Brown, R.; Katz, D. The growth of adipose tissue in children and adolescents. Cross-sectional and longitudinal studies of adipose cell number and size. J. Clin. Investig. 1979, 63, 239. [Google Scholar] [CrossRef] [PubMed]
- Calabuig-Navarro, V.; Puchowicz, M.; Glazebrook, P.; Haghiac, M.; Minium, J.; Catalano, P.; Hauguel deMouzon, S.; O’Tierney-Ginn, P. Effect of ω-3 supplementation on placental lipid metabolism in overweight and obese women. Am. J. Clin. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Leslie, M.A.; Cohen, D.J.; Liddle, D.M.; Robinson, L.E.; Ma, D.W. A review of the effect of ω-3 polyunsaturated fatty acids on blood triacylglycerol levels in normolipidemic and borderline hyperlipidemic individuals. Lipids Health Dis. 2015, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Link, J.J.; Rohatgi, A.; de Lemos, J.A. HDL cholesterol: Physiology, pathophysiology, and management. Curr. Probl. Cardiol. 2007, 32, 268–314. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Miller, M.; Tighe, A.P.; Davidson, M.H.; Schaefer, E.J. ω-3 fatty acids and coronary heart disease risk: Clinical and mechanistic perspectives. Atherosclerosis 2008, 197, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Makhoul, Z.; Kristal, A.R.; Gulati, R.; Luick, B.; Bersamin, A.; O’Brien, D.; Hopkins, S.E.; Stephensen, C.B.; Stanhope, K.L.; Havel, P.J. Associations of obesity with triglycerides and C-reactive protein are attenuated in adults with high red blood cell eicosapentaenoic and docosahexaenoic acids. Eur. J. Clin. Nutr. 2011, 65, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Couet, C.; Delarue, J.; Ritz, P.; Antoine, J.; Lamisse, F. Effect of dietary fish oil on body fat mass and basal fat oxidation in healthy adults. Int. J. Obes. Relat. Metab. Disord. 1997, 21, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Clària, J.; Nguyen, B.T.; Madenci, A.L.; Ozaki, C.K.; Serhan, C.N. Diversity of lipid mediators in human adipose tissue depots. Am. J. Physiol.-Cell Physiol. 2013, 304, C1141–C1149. [Google Scholar] [CrossRef] [PubMed]
- Baik, I.; Abbott, R.D.; Curb, J.D.; Shin, C. Intake of fish and n-3 fatty acids and future risk of metabolic syndrome. J. Am. Diet. Assoc. 2010, 110, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Jellema, A.; Ramakers, J.; Mensink, R.P. Weight loss, but not fish oil consumption, improves fasting and postprandial serum lipids, markers of endothelial function, and inflammatory signatures in moderately obese men. J. Nutr. 2007, 137, 2635–2640. [Google Scholar] [PubMed]
- Ferguson, J.F.; Mulvey, C.K.; Patel, P.N.; Shah, R.Y.; Doveikis, J.; Zhang, W.; Tabita-Martinez, J.; Terembula, K.; Eiden, M.; Koulman, A.; et al. Ω-3 PUFA supplementation and the response to evoked endotoxemia in healthy volunteers. Mol. Nutr. Food Res. 2014, 58, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Barden, A.; Mas, E.; Croft, K.D.; Phillips, M.; Mori, T.A. Short-term n-3 fatty acid supplementation but not aspirin increases plasma proresolving mediators of inflammation. J. Lipid Res. 2014, 55, 2401–2407. [Google Scholar] [CrossRef] [PubMed]
- Dawczynski, C.; Massey, K.A.; Ness, C.; Kiehntopf, M.; Stepanow, S.; Platzer, M.; Grün, M.; Nicolaou, A.; Jahreis, G. Randomized placebo-controlled intervention with n-3 LC-PUFA-supplemented yoghurt: Effects on circulating eicosanoids and cardiovascular risk factors. Clin. Nutr. 2013, 32, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.C. Specialized pro-resolving mediators: Do they circulate in plasma? J. Lipid Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an ω-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar]
- Ichimura, A.; Hirasawa, A.; Poulain-Godefroy, O.; Bonnefond, A.; Hara, T.; Yengo, L.; Kimura, I.; Leloire, A.; Liu, N.; Iida, K.; et al. Dysfunction of lipid sensor GPR120 leads to obesity in both mouse and human. Nature 2012, 483, 350–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [PubMed]
- Storch, J.; McDermott, L. Structural and functional analysis of fatty acid-binding proteins. J. Lipid Res. 2009, 50, S126–S131. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Kantartzis, K.; Machann, J.; Schick, F.; Thamer, C.; Rittig, K.; Balletshofer, B.; Machicao, F.; Fritsche, A.; Häring, H.-U. Identification and characterization of metabolically benign obesity in humans. Arch. Intern. Med. 2008, 168, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, J.; Lynch, L.; Cawood, T.J.; Kwasnik, A.; Nolan, N.; Geoghegan, J.; McCormick, A.; O’Farrelly, C.; O’Shea, D. The relationship of omental and subcutaneous adipocyte size to metabolic disease in severe obesity. PLoS ONE 2010, 5, e9997. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Wang, J.; Wu, L.; Kang, Z.B. Transgenic mice: Fat-1 mice convert n-6 to n-3 fatty acids. Nature 2004, 427, 504. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Arita, M.; Taguchi, R.; Kang, J.X.; Marette, A. Transgenic restoration of long-chain n-3 fatty acids in insulin target tissues improves resolution capacity and alleviates obesity-linked inflammation and insulin resistance in high-fat–fed mice. Diabetes 2010, 59, 3066–3073. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Horakova, O.; Brauner, P.; Rossmeisl, M.; Pecina, P.; Franssen-van Hal, N.; Ruzickova, J.; Sponarova, J.; Drahota, Z.; Vlcek, C.; et al. Polyunsaturated fatty acids of marine origin upregulate mitochondrial biogenesis and induce β-oxidation in white fat. Diabetologia 2005, 48, 2365–2375. [Google Scholar] [CrossRef] [PubMed]
- Sardi, L.; Martelli, G.; Lambertini, L.; Parisini, P.; Mordenti, A. Effects of a dietary supplement of DHA-rich marine algae on Italian heavy pig production parameters. Livest. Sci. 2006, 103, 95–103. [Google Scholar] [CrossRef]
- Skiba, G.; Poławska, E.; Raj, S.; Weremko, D.; Czauderna, M.; Wojtasik, M. The influence of dietary fatty acids on their metabolism in liver and subcutaneous fat in growing pigs. J. Anim. Feed Sci. 2011, 487, 47. [Google Scholar]
- Murali, G.; Milne, G.L.; Webb, C.D.; Stewart, A.B.; McMillan, R.P.; Lyle, B.C.; Hulver, M.W.; Saraswathi, V. Fish oil and indomethacin in combination potently reduce dyslipidemia and hepatic steatosis in LDLR−/− mice. J. Lipid Res. 2012, 53, 2186–2197. [Google Scholar] [CrossRef] [PubMed]
- Muhlhausler, B.; Miljkovic, D.; Fong, L.; Xian, C.J.; Duthoit, E.; Gibson, R.A. Maternal ω-3 supplementation increases fat mass in male and female rat offspring. Front. Genet. 2011. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, Y.B.; Hein, G.; Chicco, A. Metabolic syndrome: Effects of n-3 PUFAs on a model of dyslipidemia, insulin resistance and adiposity. Lipids 2007, 42, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Sumida, H.; Suzuki, M. Beef tallow diet decreases β-adrenergic receptor binding and lipolytic activities in different adipose tissues of rat. Metabolism 1995, 44, 1271–1277. [Google Scholar] [CrossRef]
- Awad, A.B.; Chattopadhyay, J.P. Effect of dietary saturated fatty acids on hormone-sensitive lipolysis in rat adipocytes. J. Nutr. 1986, 116, 1088–1094. [Google Scholar] [PubMed]
- Bargut, T.C.L.; Souza-Mello, V.; Mandarim-de-Lacerda, C.A.; Aguila, M.B. Fish oil diet modulates epididymal and inguinal adipocyte metabolism in mice. Food Funct. 2016, 7, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Spurlock, M.E.; Gabler, N.K. The development of porcine models of obesity and the metabolic syndrome. J. Nutr. 2008, 138, 397–402. [Google Scholar] [PubMed]
- Belzung, F.; Raclot, T.; Groscolas, R. Fish oil n-3 fatty acids selectively limit the hypertrophy of abdominal fat depots in growing rats fed high-fat diets. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1993, 264, R1111–R1118. [Google Scholar]
- Parrish, C.; Pathy, D.; Angel, A. Dietary fish oils limit adipose tissue hypertrophy in rats. Metabolism 1990, 39, 217–219. [Google Scholar] [CrossRef]
- Madsen, L.; Petersen, R.K.; Kristiansen, K. Regulation of adipocyte differentiation and function by polyunsaturated fatty acids. Biochim. Biophys. Acta 2005, 1740, 266–286. [Google Scholar] [CrossRef] [PubMed]
- González-Périz, A.; Horrillo, R.; Ferré, N.; Gronert, K.; Dong, B.; Morán-Salvador, E.; Titos, E.; Martínez-Clemente, M.; López-Parra, M.; Arroyo, V.; et al. Obesity-induced insulin resistance and hepatic steatosis are alleviated by ω-3 fatty acids: A role for resolvins and protectins. FASEB J. 2009, 23, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Neuhofer, A.; Zeyda, M.; Mascher, D.; Itariu, B.K.; Murano, I.; Leitner, L.; Hochbrugger, E.E.; Fraisl, P.; Cinti, S.; Serhan, C.N.; et al. Impaired local production of proresolving lipid mediators in obesity and 17-HDHA as a potential treatment for obesity-associated inflammation. Diabetes 2013, 62, 1945–1956. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, K.; Malick, M.; Madingou, N.; Bourque-Riel, V.; Touchette, C.; Rousseau, G. Linoleic acid attenuates cardioprotection induced by resolvin D1. J. Nutr. Biochem. 2016, 31, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Shearer, G.C.; Savinova, O.V.; Harris, W.S. Fish oil—How does it reduce plasma triglycerides? Biochim. Biophys. Acta (BBA) 2012, 1821, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Demetrius, L. Of mice and men. When it comes to studying ageing and the means to slow it down, mice are not just small humans. EMBO Rep. 2005. [Google Scholar] [CrossRef]
- Terpstra, A.H. Differences between humans and mice in efficacy of the body fat lowering effect of conjugated linoleic acid: Role of metabolic rate. J. Nutr. 2001, 131, 2067–2068. [Google Scholar] [PubMed]
- Houpt, K.A.; Houpt, T.R.; Pond, W.G. The pig as a model for the study of obesity and of control of food intake: A review. Yale J. Biol. Med. 1979, 52, 307–329. [Google Scholar] [PubMed]
- Li, S.-J.; Ding, S.-T.; Mersmann, H.J.; Chu, C.-H.; Hsu, C.-D.; Chen, C.-Y. A nutritional nonalcoholic steatohepatitis minipig model. J. Nutr. Biochem. 2016, 28, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Larsen, L.F.; Olsen, A.K.; Hansen, A.K.; Bukhave, K.; Marckmann, P. Feeding minipigs fish oil for four weeks lowers postprandial triacylglycerolemia. J. Nutr. 2003, 133, 2273–2276. [Google Scholar] [PubMed]
- Castellano, C.-A.; Audet, I.; Laforest, J.-P.; Chouinard, Y.; Matte, J.J. Fish oil diets do not improve insulin sensitivity and secretion in healthy adult male pigs. Br. J. Nutr. 2010, 103, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Behme, M.T. Dietary fish oil enhances insulin sensitivity in miniature pigs. J. Nutr. 1996, 126, 1549–1553. [Google Scholar] [PubMed]
- Yu, Y.-H.; Wu, S.-C.; Cheng, W.T.-K.; Mersmann, H.J.; Shen, T.-L.; Ding, S.-T. The function of porcine PPARγ and dietary fish oil effect on the expression of lipid and glucose metabolism related genes. J. Nutr. Biochem. 2011, 22, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Chen, C.-C.; Chen, Y.-J.; Lin, Y.-Y.; Mersmann, H.J.; Ding, S.-T. Enhanced amelioration of high-fat diet-induced fatty liver by Docosahexaenoic acid and Lysine supplementations. BioMed Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.M.; Ding, S.T. Effect of polyunsaturated fatty acids on the expression of transcription factor adipocyte determination and differentiation-dependent factor 1 and of lipogenic and fatty acid oxidation enzymes in porcine differentiating adipocytes. Br. J. Nutr. 2003, 90, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Liu, H.-Y.; Chang, Y.-T.; Cheng, Y.-H.; Mersmann, H.J.; Kuo, W.-H.; Ding, S.-T. Isolation and Differentiation of Adipose-Derived Stem Cells from Porcine Subcutaneous Adipose Tissues. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed]
- Meadus, W.J.; Duff, P.; Rolland, D.; Aalhus, J.L.; Uttaro, B.; Dugan, M.E.R. Feeding docosahexaenoic acid to pigs reduces blood triglycerides and induces gene expression for fat oxidation. Can. J. Anim. Sci. 2011, 91, 601–612. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [PubMed]
- Moseti, D.; Regassa, A.; Kim, W.-K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [PubMed]
- Forman, B.M.; Chen, J.; Evans, R.M. Hypolipidemic drugs, polyunsaturated fatty acids, and eicosanoids are ligands for peroxisome proliferator-activated receptors α and δ. Proc. Natl. Acad. Sci. USA 1997, 94, 4312–4317. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Martin, S.; Parton, R.G. Lipid droplet-organelle interactions; sharing the fats. Biochim. Biophys. Acta (BBA) 2009, 1791, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Manickam, E.; Sinclair, A.J.; Cameron-Smith, D. Suppressive actions of eicosapentaenoic acid on lipid droplet formation in 3T3-L1 adipocytes. Lipids Health Dis. 2010, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Wang, Y.; Sul, H.S. Lipolysis in adipocytes. Int. J. Biochem. Cell Biol. 2010, 42, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Brasaemle, D.L.; Rubin, B.; Harten, I.A.; Gruia-Gray, J.; Kimmel, A.R.; Londos, C. Perilipin A increases triacylglycerol storage by decreasing the rate of triacylglycerol hydrolysis. J. Biol. Chem. 2000, 275, 38486–38493. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, A.S.; Egan, J.J.; Wek, S.A.; Garty, N.B.; Blanchette-Mackie, E.; Londos, C. Perilipin, a major hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storage droplets. J. Biol. Chem. 1991, 266, 11341–11346. [Google Scholar] [PubMed]
- Barber, E.; Sinclair, A.J.; Cameron-Smith, D. Comparative actions of ω-3 fatty acids on in vitro lipid droplet formation. Prostaglandins Leukot. Essent. Fat. Acids (PLEFA) 2013, 89, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Murali, G.; Desouza, C.V.; Clevenger, M.E.; Ramalingam, R.; Saraswathi, V. Differential effects of eicosapentaenoic acid and docosahexaenoic acid in promoting the differentiation of 3T3-L1 preadipocytes. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-K.; Della-Fera, M.; Lin, J.; Baile, C.A. Docosahexaenoic acid inhibits adipocyte differentiation and induces apoptosis in 3T3-L1 preadipocytes. J. Nutr. 2006, 136, 2965–2969. [Google Scholar] [PubMed]
- Jiang, H.-P.; Serrero, G. Isolation and characterization of a full-length cDNA coding for an adipose differentiation-related protein. Proc. Natl. Acad. Sci. USA 1992, 89, 7856–7860. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Takahashi, K.; Nishimaki-Mogami, T.; Kagechika, H.; Yamamoto, M.; Itabe, H. Docosahexaenoic acid induces adipose differentiation-related protein through activation of retinoid x receptor in human choriocarcinoma BeWo cells. Biol. Pharm. Bull. 2009, 32, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Hengbo, S.; Jun, L.; Jun, L.; Wangsheng, Z.; Huibin, T.; Huaiping, S. PPARG modulated lipid accumulation in dairy gmec via regulation of ADRP gene. J. Cell. Biochem. 2015, 116, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Tobin, K.A.R.; Harsem, N.K.; Dalen, K.T.; Staff, A.C.; Nebb, H.I.; Duttaroy, A.K. Regulation of ADRP expression by long-chain polyunsaturated fatty acids in BeWo cells, a human placental choriocarcinoma cell line. J. Lipid Res. 2006, 47, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, C.; Hong, Y.-H.; Iga, T.; Hishikawa, D.; Suzuki, Y.; Song, S.-H.; Choi, K.-C.; Adachi, T.; Hirasawa, A.; Tsujimoto, G. The regulation of adipogenesis through GPR120. Biochem. Biophys. Res. Commun. 2007, 354, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, A.; Hara, T.; Hirasawa, A. Regulation of energy homeostasis via GPR120. Front. Endocrinol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Reddy, J.K.; Hashimoto, T. Peroxisomal β-oxidation and peroxisome proliferator-activated receptor α: An adaptive metabolic system. Annu. Rev. Nutr. 2001, 21, 193–230. [Google Scholar] [CrossRef] [PubMed]
- Van Veldhoven, P.P.; Mannaerts, G.P. Role and organization of peroxisomal β-oxidation. In Current Views of Fatty Acid Oxidation and Ketogenesis; Springer: Berlin/Heidelberg, Germany, 2002; pp. 261–272. [Google Scholar]
- Gao, X.; Li, K.; Hui, X.; Kong, X.; Sweeney, G.; Wang, Y.; Xu, A.; Teng, M.; Liu, P.; Wu, D. Carnitine palmitoyltransferase 1A prevents fatty acid-induced adipocyte dysfunction through suppression of c-Jun N-terminal kinase. Biochem. J. 2011, 435, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Xie, W.; Lei, T.; Hamilton, J.A. Eicosapentaenoic acid, but not oleic acid, stimulates β-oxidation in adipocytes. Lipids 2005, 40, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Vluggens, A.; Andreoletti, P.; Viswakarma, N.; Jia, Y.; Matsumoto, K.; Kulik, W.; Khan, M.; Huang, J.; Guo, D.; Yu, S.; et al. Functional significance of the two ACOX1 isoforms and their crosstalks with PPARα and RXRα. Lab. Investig. 2010, 90, 696–708. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Rossmeisl, M.; Kuda, O.; Kopecky, J. Stimulation of mitochondrial oxidative capacity in white fat independent of UCP1: A key to lean phenotype. Biochim. Biophys. Acta (BBA) 2013, 1831, 986–1003. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Liu, D.Y.; Ma, L.Q.; Luo, Z.D.; Cao, T.B.; Zhong, J.; Yan, Z.C.; Wang, L.J.; Zhao, Z.G.; Zhu, S.J.; et al. Activation of transient receptor potential vanilloid type-1 channel prevents adipogenesis and obesity. Circ. Res. 2007, 100, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, F.; Inoue, N.; Masamoto, Y.; Matsumura, S.; Kimura, W.; Kadowaki, M.; Higashi, T.; Tominaga, M.; Inoue, K.; Fushiki, T. Non-pungent capsaicin analogs (capsinoids) increase metabolic rate and enhance thermogenesis via gastrointestinal TRPV1 in mice. Biosci. Biotechnol. Biochem. 2009, 73, 2690–2697. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Tsukamoto-Yasui, M.; Hara-Kimura, Y.; Inoue, N.; Nogusa, Y.; Okabe, Y.; Nagashima, K.; Kato, F. Intragastric administration of capsiate, a transient receptor potential channel agonist, triggers thermogenic sympathetic responses. J. Appl. Physiol. 2011, 110, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Matta, J.A.; Miyares, R.L.; Ahern, G.P. TRPV1 is a novel target for ω-3 polyunsaturated fatty acids. J. Physiol. 2007, 578, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Periz, A.; Planaguma, A.; Gronert, K.; Miquel, R.; Lopez-Parra, M.; Titos, E.; Horrillo, R.; Ferre, N.; Deulofeu, R.; Arroyo, V.; et al. Docosahexaenoic acid (DHA) blunts liver injury by conversion to protective lipid mediators: Protectin D1 and 17S-hydroxy-DHA. FASEB J. 2006, 20, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Uhlar, C.M.; Whitehead, A.S. Serum amyloid A, the major vertebrate acute-phase reactant. Eur. J. Biochem. 1999, 265, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Björk, C.; Wilhelm, U.; Mandrup, S.; Larsen, B.D.; Bordoni, A.; Hedén, P.; Rydén, M.; Arner, P.; Laurencikiene, J. Effects of selected bioactive food compounds on human white adipocyte function. Nutr. Metab. 2016, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odegaard, J.I.; Chawla, A. The immune system as a sensor of the metabolic state. Immunity 2013, 38, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Marks, K.A.; Marvyn, P.M.; Henao, J.J.; Bradley, R.M.; Stark, K.D.; Duncan, R.E. Fasting enriches liver triacylglycerol with n-3 polyunsaturated fatty acids: Implications for understanding the adipose-liver axis in serum docosahexaenoic acid regulation. Genes Nutr. 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Bhaswant, M.; Poudyal, H.; Brown, L. Mechanisms of enhanced insulin secretion and sensitivity with n-3 unsaturated fatty acids. J. Nutr. Biochem. 2015, 26, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Sugden, M.C.; Bulmer, K.; Gibbons, G.F.; Knight, B.L.; Holness, M.J. Peroxisome-proliferator-activated receptor-α (PPARα) deficiency leads to dysregulation of hepatic lipid and carbohydrate metabolism by fatty acids and insulin. Biochem. J. 2002, 364, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Caron-Dorval, D.; Verreault, M.; Couture, P.; Deshaies, Y.; Barbier, O.; Vohl, M.C. PPARα L162V polymorphism alters the potential of n-3 fatty acids to increase lipoprotein lipase activity. Mol. Nutr. Food Res. 2010, 54, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Ide, T.; Murata, M.; Sugano, M. Stimulation of the activities of hepatic fatty acid oxidation enzymes by dietary fat rich in alpha-linolenic acid in rats. J. Lipid Res. 1996, 37, 448–463. [Google Scholar] [PubMed]
- Ide, T.; Kobayashi, H.; Ashakumary, L.; Rouyer, I.A.; Takahashi, Y.; Aoyama, T.; Hashimoto, T.; Mizugaki, M. Comparative effects of perilla and fish oils on the activity and gene expression of fatty acid oxidation enzymes in rat liver. Biochim. Biophys. Acta 2000, 1485, 23–35. [Google Scholar] [CrossRef]
- Gondret, F.; Ferré, P.; Dugail, I. ADD-1/SREBP-1 is a major determinant of tissue differential lipogenic capacity in mammalian and avian species. J. Lipid Res. 2001, 42, 106–113. [Google Scholar] [PubMed]
- Nakatani, T.; Kim, H.-J.; Kaburagi, Y.; Yasuda, K.; Ezaki, O. A low fish oil inhibits SREBP-1 proteolytic cascade, while a high-fish-oil feeding decreases SREBP-1 mRNA in mice liver relationship to anti-obesity. J. Lipid Res. 2003, 44, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Chen, C.-C.; Li, T.-K.; Wang, P.-H.; Liu, L.-R.; Chang, F.-Y.; Wang, Y.-C.; Yu, Y.-H.; Lin, S.-P.; Mersmann, H.J.; et al. Docosahexaenoic acid suppresses the expression of FoxO and its target genes. J. Nutr. Biochem. 2012, 23, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Neuschwander-Tetri, B.A. Hepatic Lipotoxicity and the Pathogenesis of Nonalcoholic Steatohepatitis: The Central Role of Nontriglyceride Fatty Acid Metabolites. Hepatology 2010, 52, 774–788. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A.; Jokelainen, K.; Tipoe, G.L.; Rahemtulla, A.; Dannenberg, A.J. Dietary saturated fatty acids reverse inflammatory and fibrotic changes in rat liver despite continued ethanol administration. J. Pharmacol. Exp. Ther. 2001, 299, 638–644. [Google Scholar] [PubMed]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.M.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the toll-like receptor 4/NF-κB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Dannenberg, A.J.; Nanji, A.A. Dietary saturated fatty acids: A novel treatment for alcoholic liver disease. Alcohol. Clin. Exp. Res. 1998, 22, 750–752. [Google Scholar] [CrossRef]
- Mei, S.; Ni, H.M.; Manley, S.; Bockus, A.; Kassel, K.M.; Luyendyk, J.P.; Copple, B.L.; Ding, W.X. Differential roles of unsaturated and saturated fatty acids on autophagy and apoptosis in hepatocytes. J. Pharmacol. Exp. Ther. 2011, 339, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, L.; Magliocco, O.; Spampinato, D.; Piro, S.; Oliveri, C.; Alagona, C.; Papa, G.; Rabuazzo, A.M.; Purrello, F. Effects of n-3 polyunsaturated fatty acids in subjects with nonalcoholic fatty liver disease. Dig. Liver Dis. 2008, 40, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Leinonen, E.; Hurt-Camejo, E.; Wiklund, O.; Hultén, L.M.; Hiukka, A.; Taskinen, M.-R. Insulin resistance and adiposity correlate with acute-phase reaction and soluble cell adhesion molecules in type 2 diabetes. Atherosclerosis 2003, 166, 387–394. [Google Scholar] [CrossRef]
- Wang, Y.C.; Kuo, W.H.; Chen, C.Y.; Lin, H.Y.; Wu, H.T.; Liu, B.H.; Chen, C.H.; Mersmann, H.J.; Chang, K.J.; Ding, S.T. Docosahexaenoic acid regulates serum amyloid A protein to promote lipolysis through down regulation of perilipin. J. Nutr. Biochem. 2010, 21, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.C.; Chen, C.Y.; Lee, H.S.; Wang, Y.C.; Li, T.K.; Mersamm, H.J.; Ding, S.T.; Wang, P.H. Docosahexaenoic acid enhances hepatic serum amyloid A expression via protein kinase A-dependent mechanism. J. Biol. Chem. 2009, 284, 32239–32247. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Chen, C.; Cheng, W.; Ding, S. The effect of dietary docosahexaenoic acid enrichment on the expression of porcine hepatic genes. Asian Australas. J. Anim. Sci. 2007, 20, 768–774. [Google Scholar] [CrossRef]
- Chen, C.H.; Wang, P.H.; Liu, B.H.; Hsu, H.H.; Mersmann, H.J.; Ding, S.T. Serum amyloid A protein regulates the expression of porcine genes related to lipid metabolism. J. Nutr. 2008, 138, 674–679. [Google Scholar] [PubMed]
- Liu, L.R.; Lin, S.P.; Chen, C.C.; Chen, Y.J.; Tai, C.C.; Chang, S.C.; Juang, R.H.; Tseng, Y.W.; Liu, B.H.; Mersmann, H.J. Serum Amyloid A induces lipolysis by downregulating perilipin through ERK1/2 and PKA signaling pathways. Obesity 2011, 19, 2301–2309. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y.; Eren, F.; Yonal, O.; Kurt, R.; Aktas, B.; Celikel, C.A.; Ozdogan, O.; Imeryuz, N.; Kalayci, C.; Avsar, E. Increased serum FGF21 levels in patients with nonalcoholic fatty liver disease. Eur. J. Clin. Investig. 2010, 40, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Mangelsdorf, D.J. Fibroblast growth factor 21: From pharmacology to physiology. Am. J. Clin. Nutr. 2010, 91, 254S–257S. [Google Scholar] [CrossRef] [PubMed]
- Giannini, C.; Feldstein, A.E.; Santoro, N.; Kim, G.; Kursawe, R.; Pierpont, B.; Caprio, S. Circulating levels of FGF-21 in obese youth: Associations with liver fat content and markers of liver damage. J. Clin. Endocrinol. Metab. 2013, 98, 2993–3000. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zhou, Y.; Chen, S.-H.; Zhao, X.-L.; Ran, L.; Zeng, X.-L.; Wu, Y.; Chen, J.-L.; Kang, C.; Shu, F.-R.; et al. Fish oil supplements lower serum lipids and glucose in correlation with a reduction in plasma fibroblast growth factor 21 and prostaglandin E2 in nonalcoholic fatty liver disease associated with hyperlipidemia: A randomized clinical trial. PLoS ONE 2015, 10, e0133496. [Google Scholar] [CrossRef] [PubMed]
- Mandard, S.; Zandbergen, F.; van Straten, E.; Wahli, W.; Kuipers, F.; Müller, M.; Kersten, S. The fasting-induced adipose factor/angiopoietin-like protein 4 is physically associated with lipoproteins and governs plasma lipid levels and adiposity. J. Biol. Chem. 2006, 281, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Puthanveetil, P.; Wang, F.; Kim, M.S.; Abrahani, A.; Rodrigues, B. Severity of diabetes governs vascular lipoprotein lipase by affecting enzyme dimerization and disassembly. Diabetes 2011, 60, 2041–2050. [Google Scholar] [CrossRef] [PubMed]
- Brands, M.; Sauerwein, H.P.; Ackermans, M.T.; Kersten, S.; Serlie, M.J. ω-3 long-chain fatty acids strongly induce angiopoietin-like 4 in humans. J. Lipid Res. 2013, 54, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S.; Lichtenstein, L.; Steenbergen, E.; Mudde, K.; Hendriks, H.F.; Hesselink, M.K.; Schrauwen, P.; Müller, M. Caloric restriction and exercise increase plasma ANGPTL4 levels in humans via elevated free fatty acids. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Ruge, T.; Sukonina, V.; Kroupa, O.; Makoveichuk, E.; Lundgren, M.; Svensson, M.K.; Olivecrona, G.; Eriksson, J.W. Effects of hyperinsulinemia on lipoprotein lipase, angiopoietin-like protein 4, and glycosylphosphatidylinositol-anchored high-density lipoprotein binding protein 1 in subjects with and without type 2 diabetes mellitus. Metabolism 2012, 61, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Aranceta, J.; Pérez-Rodrigo, C. Recommended dietary reference intakes, nutritional goals and dietary guidelines for fat and fatty acids: A systematic review. Br. J. Nutr. 2012, 107, S8–S22. [Google Scholar] [CrossRef] [PubMed]
- Gorjão, R.; Azevedo-Martins, A.K.; Rodrigues, H.G.; Abdulkader, F.; Arcisio-Miranda, M.; Procopio, J.; Curi, R. Comparative effects of DHA and EPA on cell function. Pharmacol. Ther. 2009, 122, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Moleres, A.; Ochoa, M.C.; Rendo-Urteaga, T.; Martínez-González, M.A.; San Julián, M.C.A.; Martínez, J.A.; Marti, A. Dietary fatty acid distribution modifies obesity risk linked to the rs9939609 polymorphism of the fat mass and obesity-associated gene in a Spanish case-control study of children. Br. J. Nutr. 2012, 107, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Surma, M.A.; Herzog, R.; Vasilj, A.; Klose, C.; Christinat, N.; Morin-Rivron, D.; Simons, K.; Masoodi, M.; Sampaio, J.L. An automated shotgun lipidomics platform for high throughput, comprehensive, and quantitative analysis of blood plasma intact lipids. Eur. J. Lipid Sci. Technol. 2015, 117, 1540–1549. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 99, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Walenta, E.; Akiyama, T.E.; Lagakos, W.S.; Lackey, D.; Pessentheiner, A.R.; Sasik, R.; Hah, N.; Chi, T.J.; Cox, J.M.; Powels, M.A.; et al. A GPR120-selective agonist improves insulin resistance and chronic inflammation in obese mice. Nat. Med. 2014, 20, 942–947. [Google Scholar]
- Titos, E.; Rius, B.; González-Périz, A.; López-Vicario, C.; Morán-Salvador, E.; Martínez-Clemente, M.; Arroyo, V.; Clària, J. Resolvin D1 and its precursor docosahexaenoic acid promote resolution of adipose tissue inflammation by eliciting macrophage polarization toward an M2-like phenotype. J. Immunol. 2011, 187, 5408–5418. [Google Scholar] [CrossRef] [PubMed]
- Darabi, M.; Byagowi, S.; Fayezi, S.; Darabi, M.; Mirshahvaladi, S.; Sahmani, M. Transcriptional regulation of Δ6-desaturase by peroxisome proliferative-activated receptor δ agonist in human pancreatic cancer cells: Role of MEK/ERK1/2 pathway. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Kitson, A.P.; Stroud, C.K.; Stark, K.D. Elevated production of docosahexaenoic acid in females: Potential molecular mechanisms. Lipids 2010, 45, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Shida, T.; Kamei, N.; Takeda-Morishita, M.; Isowa, K.; Takayama, K. Colonic delivery of docosahexaenoic acid improves impaired glucose tolerance via GLP-1 secretion and suppresses pancreatic islet hyperplasia in diabetic KK-A y mice. Int. J. Pharm. 2013, 450, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Capel, F.; Acquaviva, C.; Pitois, E.; Laillet, B.; Rigaudière, J.-P.; Jouve, C.; Pouyet, C.; Gladine, C.; Comte, B.; Saban, C.V. DHA at nutritional doses restores insulin sensitivity in skeletal muscle by preventing lipotoxicity and inflammation. J. Nutr. Biochem. 2015, 26, 949–959. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Treatment or Dosage | Duration | Observation | Reference |
|---|---|---|---|---|
| Mother-child pair | 1. At 29 weeks gestation, the food-frequency questionnaire (FFQ) quantified the average frequency of consumption of >140 specified foods and beverage. 2. 0.15 ± 0.14 g DHA + EPA/day. | 3 months | 1. Maternal plasma DHA + EPA was 1.9% ± 0.6%, and umbilical plasma concentration was 4.6% ± 1.2%. 2. DHA + EPA = lower skin fold thickness and odds of obesity. 3. Maternal plasma DHA + EPA not associated with child adiposity. | [7] |
| Healthy men | 1. Fish diet group: 4.3 ± 0.5 fish meals (provided 0.38 ± 0.04 g EPA and 0.67 ± 0.09 g) per week. 2. Fish oil group: oil 1.33 g EPA and 0.95 g DHA per day. 3. DHA-oil group: DHA-oil (EPA-free) 1.68 g DHA per day. | 15 weeks | Fish, fish oil or DHA = lower plasma TAG concentration and total chylomicron+increased HDL2/HDL3 cholesterol. | [72] |
| Healthy men | 0, 3, 6, or 12 capsules/day, (Each capsule provided 300 mg EPA and 200 mg DHA) | 12 weeks | 3 and 6 g n-3 fatty acids were similar, but VLDL, LDL and total HDL-cholesterol subtractions were no significantly different. | [73] |
| Moderately hyperlipidemic but otherwise healthy men | 1. Placebo (7.5 g olive oil/day) 2. DHA capsules (7.5 g DHA oil/day) | 90 days | 1. The inflammatory markers was no difference within 45 days. 2. The CRP and IL-6 decreased, and anti-inflammatory matrix metalloproteinase-2 was increased at 90 days. 3. SAA positively associated with sum of saturated fatty acids. | [74] |
| Healthy weight, overweight and obese adults | 2 g/day of algal DHA | 4.5 months | 1. DHA-supplemented group, the decrease in mean VLDL particle size and increase in LDL and HDL. 2. DHA supplementation reduced VLDL and total TAG. | [75] |
| Healthy weight, overweight and obese adults | 1. >2 fatty fish meals/week 2. Restricted diet | Not mentioned | 1. Plasma n-3 PUFA lower in obese participants. 2. Inversely correlated with EPA and DHA intakes. | [69] |
| Metabolic syndrome | 1. Control 2. 1 g fish oil containing 180 mg EPA + 120 mg DHA | 6 months | Reduced body weight and serum concentration of LDL-cholesterol, TAG in fish oil group. | [76] |
| Metabolic syndrome | 1. Placebo = 4 or 6 g, soybean oil/day 2. Low flaxseed oil = 2.2 g ALA/day 3. High flaxseed oil = 6.6 g ALA/day 4. Low fish oil = 700 mg EPA + 500 mg DHA/day 5. High fish oil = 2.1 g EPA + 1.5 g DHA/day | 8 weeks | 1. The inflammatory marker were no significantly different. 2. Decreased plasma TAG and blood pressure decreased significantly+ increased LDL cholesterol. | [77] |
| Overweight hyperinsulinaemic women | 1. Control = no weight loss + placebo oil 2. Weight loss program 10% weight loss + 5 g n-3 PUFA containing 1.3 g EPA + 2.9 DHA/day 3. Weight loss + placebo oil containing 2.8 g linoleic acid and 1.4 g oleic acid/day | 24 weeks | 1. Weight loss with both diets. 2. Diet n-3 PUFA increased adipose tissue n-3 PUFA. 3. Weight loss group improved insulin sensitivity. 4. n-3 PUFA increased plasma TAG and adipoinectin. | [78] |
| Species | Breed | Treatment | Duration | Observation | Reference |
|---|---|---|---|---|---|
| Mice | C57BL/6 | 1. Standard chow group 2. High-fat lard group 3. High-fat lard plus fish oil (FO) group (40 g soybean oil + 119 lard + 119 FO) 4. High fat fish oil group (40 g soybean oil + 238 g FO) | 8 weeks | Fish oil decreased adipose tissue, body mass gain and insulin resistance even with lard. | [115] |
| Mice | Male C57BL/6J | 1. 20% flax-seed oil 2. sHFf-F2 diet and 44% of lipids were replaced by n-3 PUFA concentrate (6% EPA and 51% DHA) | 5 weeks | 1. Fatty acid oxidation genes, Ppargc 1α, Nrf1 and Cpt1a elevated in epididymal fat by EPA + DHA. 2. Mitochondrial protein and induce β-oxidation in edidymal fat but not dorsolumbar fat in EPA + DHA. | [107] |
| Mice | Muscle specific-PPARγ transgenic mice | 1. Control, 58% carbohydrate, 13.5% fat and 28.5% protein 2. Control + 4 mg Rosiglitazone/kg 3. High fish oil group containing (36% carbohydrate, 35.5% fat and 28.5% protein) 4. High-beef tallow diet (36% carbohydrate, 35.5% fat and 28.5% protein) | 4 months | 1. The PPARγ transgenic mice increased the expression of muscle Glut 4. 2. Fish oil group increased adipogenic and glucose uptake genes and lower blood glucose in transgenic mice. 3. Adiponectin elevated by fish oil. | [131] |
| Mice | Male C57BL/6 mice | Treatment: High fat diet induction for 23 weeks, then daily gavage with supplement. Supplement: 1. 0.5 or 1% DHA (based on the average daily dietary intake) 2. 1% DHA 3. 0.5% lysine + 1% DHA 4. 1% lysine + 1% DHA | 4 weeks | 1. The mRNA expression of hepatic pro inflammatory cytokines were suppressed by DHA and combinations of DHA + Lysine. 2. The lipogenic gene, ACC1 was suppressed by DHA. 3. Combination of DHA and lysine inhibited ACC1, fatty acid synthase, lipoprotein lease in gonadal adipose tissue. 4. The symptoms of nonalcoholic fatty liver disease were decreased by DHA and lysine. | [132] |
| Mice | 1. Male C57BL/6J wild-type (WT) mice 2. Male BKS.Cg-Dock 7m+/+Leprdb/J (db/db) mice 3. Lean nondiabetic littermates (db/+) | Treatment: 1. 18 weeks of the HF diet (60% kcal from fat) or 2. low-fat (LF) control diet (10% kcal from fat) Dose: 1. DHA (4 µg/g body weight),17-HDHA (50 ng/g body weight) 2. Control (0.9% NaCl containing 3% delipidated fatty acid-free BSA and 2% ethanol) | Intraperitoneal injection every 12 h for 8 days or continuous application with osmotic (Alzet) pumps (∼120) for 15 days | 1. Genetic and diet-induced obesity decreases adipose tissue n-3 PUFA–derived lipid mediators 17-HDHA and PD1. 2. Adipose tissue 17-HDHA and PD1 decreased after only 4 days of HF diet. 3. Greater adipose tissue 17-HDHA, which reduced adipose tissue inflammation. 4. Attenuated inflammation and improved insulin sensitivity induced by n-3 PUFA linked to increase SPMs and their precursors in adipose tissue. 5. Treatment with 17-HDHA reduces obesity-induced adipose tissue inflammation. 6. Treatment with 17-HDHA improves metabolic regulation in obesity. | [121] |
| Rat | Wistar rats | 1. Control (sucrose replaced by starch) 2. Sucrose-rich (SRD) diet (corn oil, 8/100 g) 3. SRD + fish oil, fish oil 7/100 g + 1/100g corn oil | 6 + 2 months | 1. Fish oil decreased plasma TAG, VLDL and adipocyte size in sucrose diet. 2. Fish oil reversed dyslipidemia and improved insulin action and glucose sensitivity in muscle. | [112] |
| Rat | Female wistar Rat | Treatment: 1. Control (5% fat, 0.22% n-3 PUFA of total fatty acids) 2. 5% fat, 1.29% n-3 PUFA of total fatty acids Dose: 1. 6.5~9.0 mg n-3 PUFA/day | Dams = 3 weeks gestation + 3 weeks suckling; Offspring = 3 weeks post-weaning | 1. n-3 PUFA increased body fat in males and females at 6 weeks of age. 2. Fat mostly subcutaneous depots, but no up-regulation of adipogenic/lipogenic gene transcription. | [111] |
| Rat | Sprague-dailey male rats | Treatment: 1. 14% coconut oil, beef fat or safflower oil 2. 900 mg/g 3. 10% n-3 PUFA | 4 weeks | 1. Norepinephrine-stimulated lipolysis was 50% lower in saturate diet. 2. The activities of 3’-5’-cyclic nucleotide (cAMP) phosphodiesterase and hormone sensitive lipase were lower in saturated fatty acids compared with polyunsaturated fatty acids. | [114] |
| Pig | Male castrated minipigs | Treatment: 1. Diet enriched with cod liver oil 2. Diet enriched with a mixture of sunflower palm and olive oil Dose: 1. ∼2.5 g/day (n-3) PUFA | 4 weeks | 1. The fish oil-enriched diet was associated with lower TAG, glycerol and nonesterified fatty acid concentrations in the hours after the gastric fat load than the control diet. 2. No significant effect of fish oil supplementation on plasma triacylglycerol clearance in minipigs. | [128] |
| Pig | Healthy duroc boars | Dose: 1. 62 g hydrogenated animals fat 2. 60 g menhaden oil containing 10.8 g DHA and 9.0 g EPA 3. 60 g tuna oil containing 19.8 g DHA and 3.9 g EPA | 7 months | Long term supplementation of dietary n-3 PUFA did not affect insulin metabolism, but n-3 PUFA increase the fat accumulation. | [129] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-W.; Chien, Y.-S.; Chen, Y.-J.; Ajuwon, K.M.; Mersmann, H.M.; Ding, S.-T. Role of n-3 Polyunsaturated Fatty Acids in Ameliorating the Obesity-Induced Metabolic Syndrome in Animal Models and Humans. Int. J. Mol. Sci. 2016, 17, 1689. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101689
Huang C-W, Chien Y-S, Chen Y-J, Ajuwon KM, Mersmann HM, Ding S-T. Role of n-3 Polyunsaturated Fatty Acids in Ameliorating the Obesity-Induced Metabolic Syndrome in Animal Models and Humans. International Journal of Molecular Sciences. 2016; 17(10):1689. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101689
Chicago/Turabian StyleHuang, Chao-Wei, Yi-Shan Chien, Yu-Jen Chen, Kolapo M. Ajuwon, Harry M. Mersmann, and Shih-Torng Ding. 2016. "Role of n-3 Polyunsaturated Fatty Acids in Ameliorating the Obesity-Induced Metabolic Syndrome in Animal Models and Humans" International Journal of Molecular Sciences 17, no. 10: 1689. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101689