
Auraptene and Other Prenyloxyphenylpropanoids Suppress Microglial Activation and Dopaminergic Neuronal Cell Death in a Lipopolysaccharide-Induced Model of Parkinson’s Disease

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
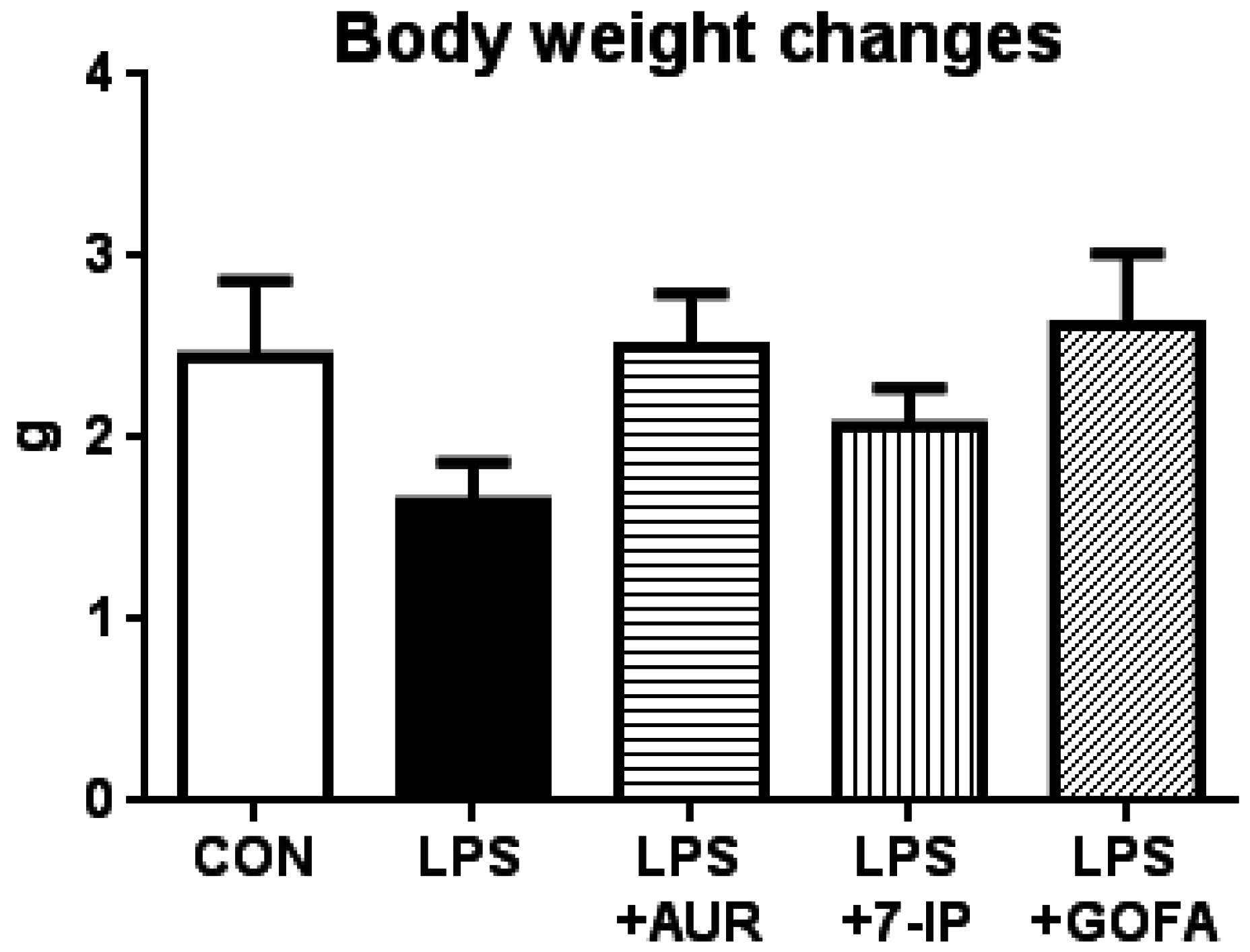
2.1. Effects of AUR, 7-IP, and GOFA on the LPS-Induced Sickness Response
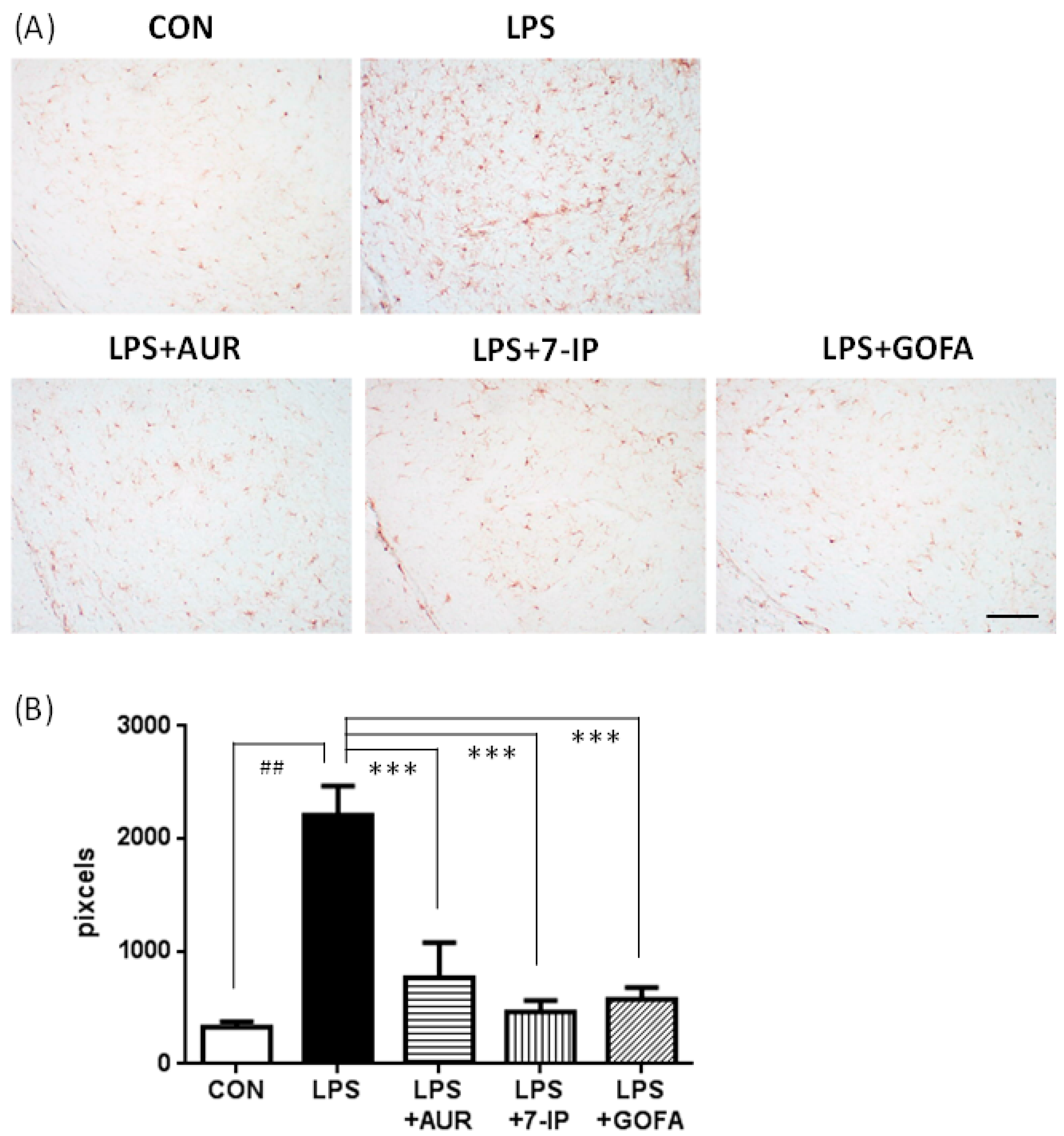
2.2. Effects of AUR, 7-IP, and GOFA on LPS-Induced Microglial Hyperactivation in the SN
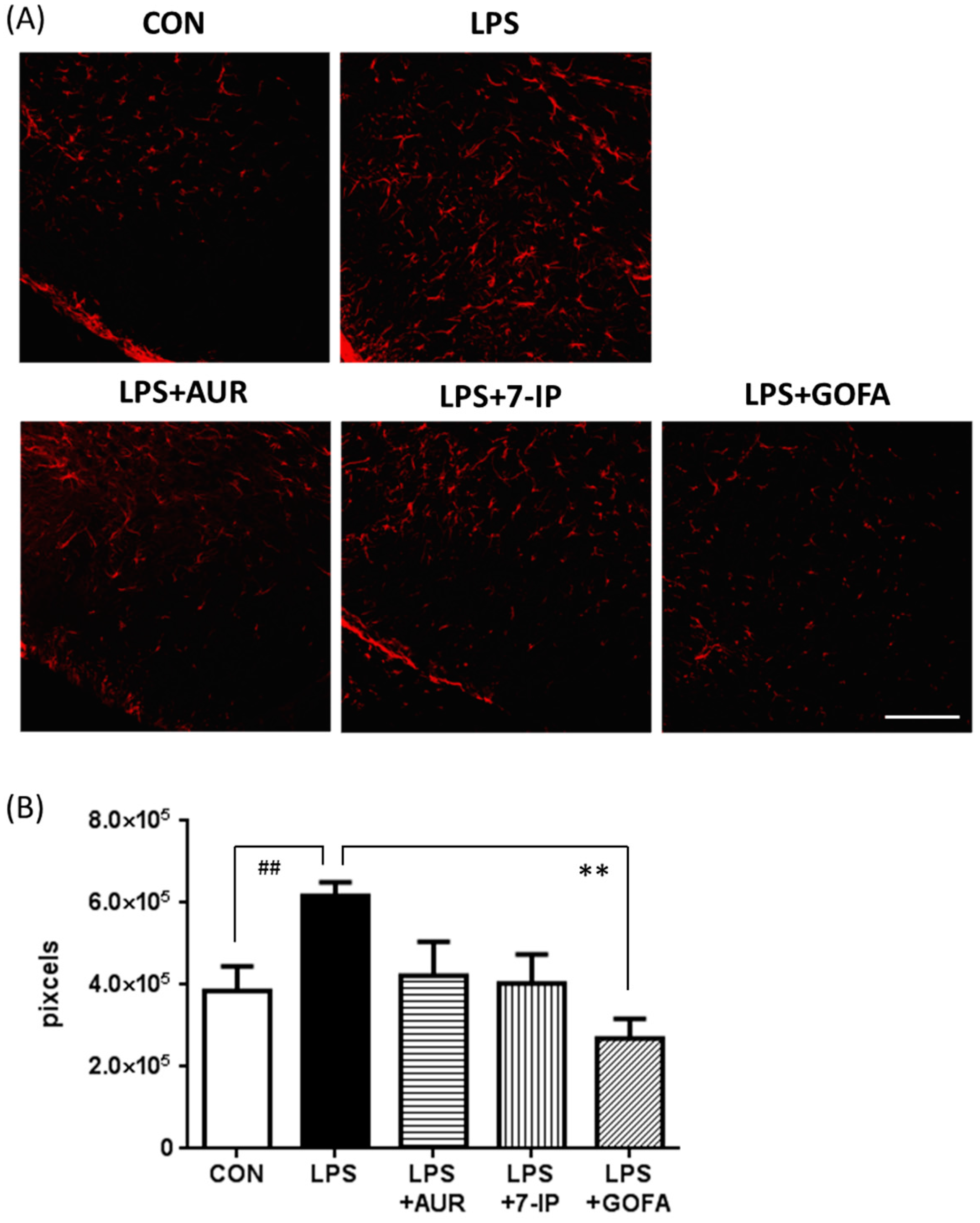
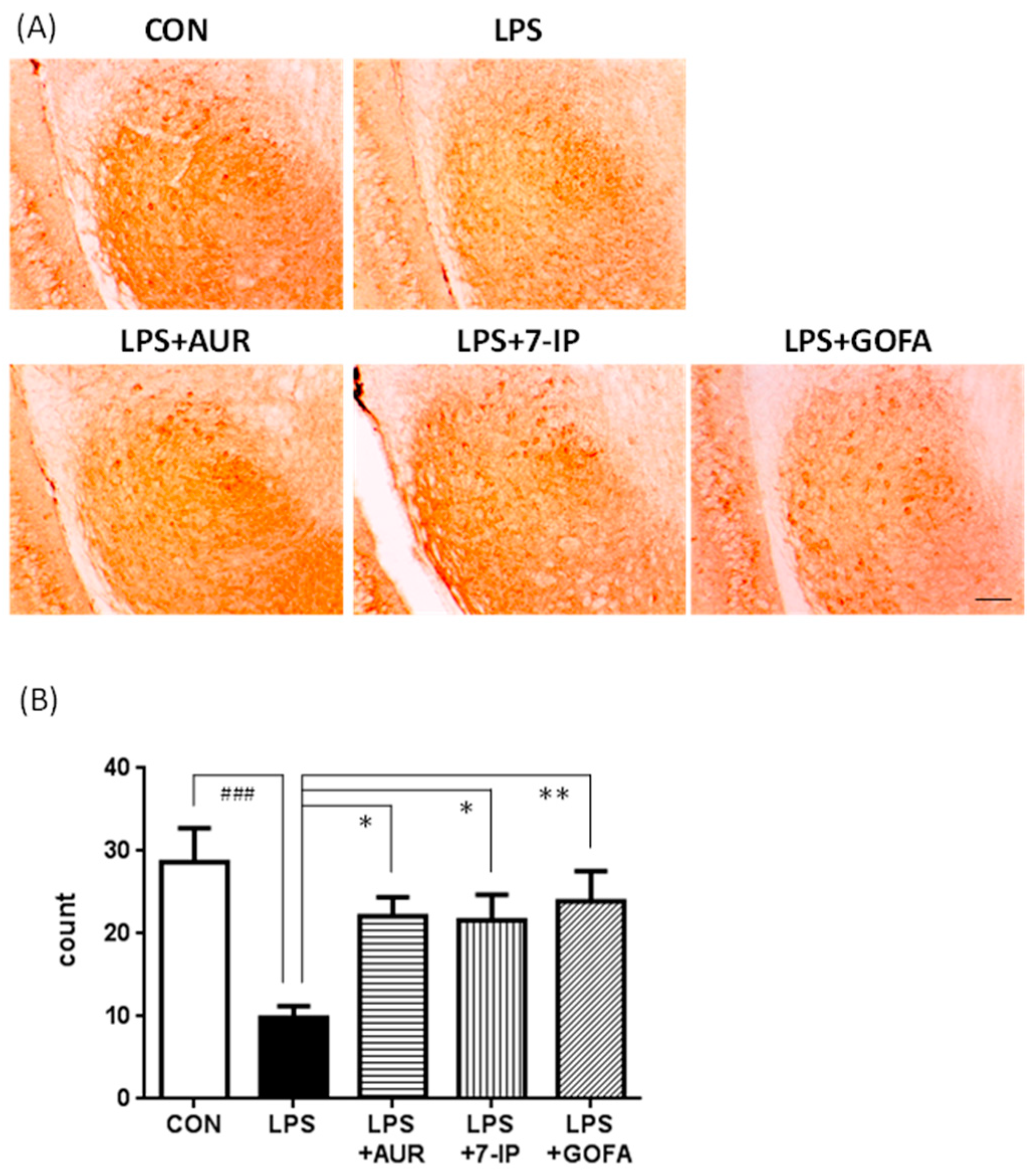
2.3. Effects of AUR, 7-IP, and GOFA on the LPS-Induced Activation of Astrocytes in the SN
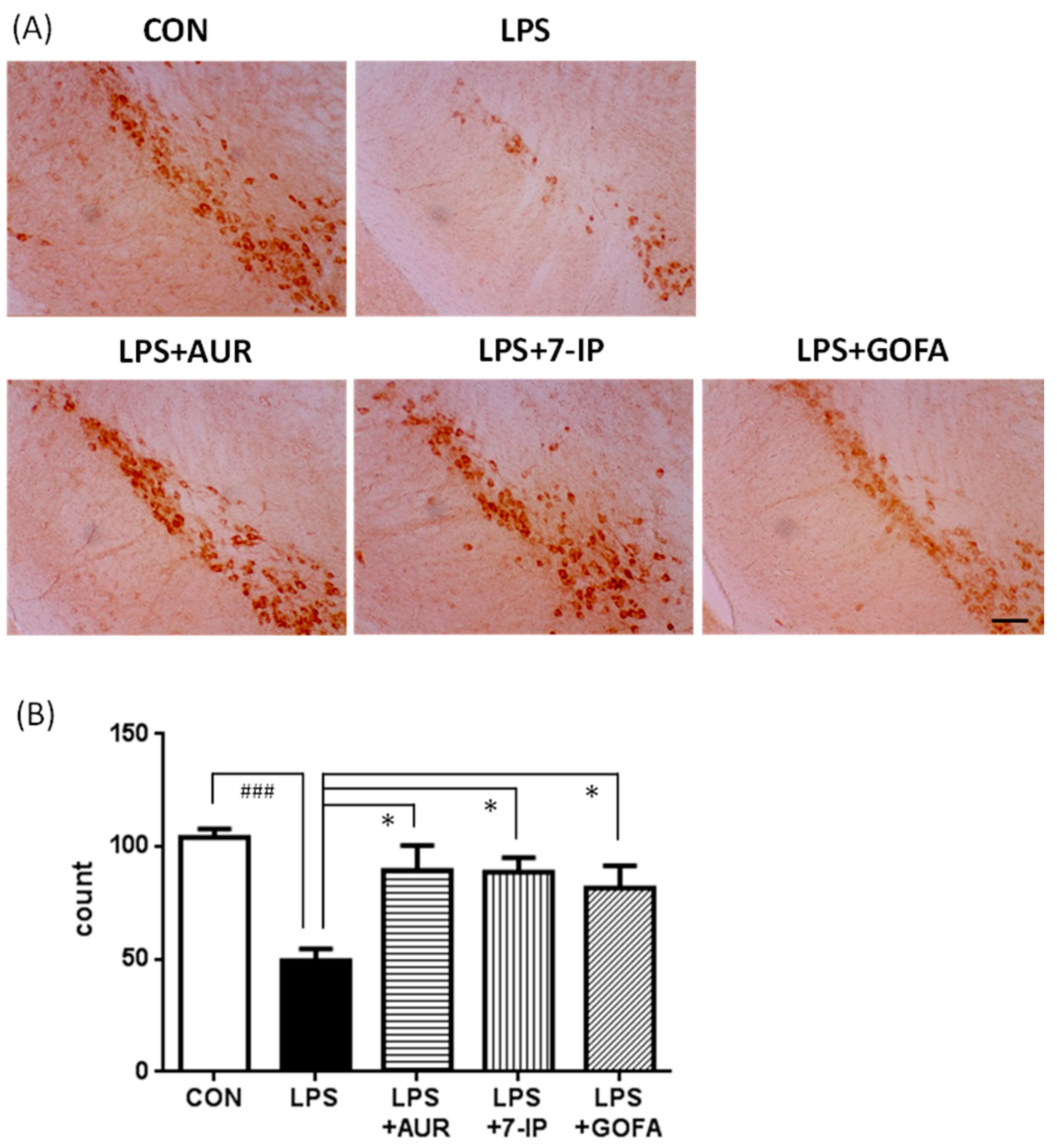
2.4. Effects of AUR, 7-IP, and GOFA on LPS-Induced Neuronal Cell Loss in the SN
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Intranigral Injection of LPS
4.4. Administration of AUR, 7-IP, or GOFA
4.5. Immunohistochemistry for Optical Microscopy
4.6. Immunohistochemistry for Confocal Fluorescence Microscopy
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Epifano, F.; Genovese, S.; Menghini, L.; Curini, M. Chemistry and pharmacology of oxyprenylated secondary plant metabolites. Phytochemistry 2007, 68, 939–953. [Google Scholar] [CrossRef] [PubMed]
- Curini, M.; Epifano, F.; Maltese, F.; Marcotullio, M.C.; Tubaro, A.; Altinier, G.; Gonzales, S.P.; Rodriguez, J.C. Synthesis and anti-inflammatory activity of natural and semisynthetic geranyloxycoumarins. Bioorg. Med. Chem. Lett. 2004, 14, 2241–2243. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yasui, Y.; Ishigamori-Suzuki, R.; Oyama, T. Citrus compounds inhibit inflammation- and obesity-related colon carcinogenesis in mice. Nutr. Cancer 2008, 60 (Suppl. S1), 70–80. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; de Azevedo, M.B.; Durán, N.; Alderete, J.B.; Epifano, F.; Genovese, S.; Tanaka, M.; Tanaka, T.; Curini, M. Colorectal cancer chemoprevention by 2β-cyclodextrin inclusion compounds of auraptene and 4′-geranyloxyferulic acid. Int. J. Cancer 2010, 126, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Shigemori, T.; Ohigashi, H. Zingiberaceous and citrus constituents, 1′-acetoxychavicol acetate, zerumbone, auraptene, and nobiletin, suppress lipopolysaccharide-induced cyclooxygenase-2 expression in RAW264.7 murine macrophages through different modes of action. J. Nutr. 2005, 135, 2987S–2992S. [Google Scholar] [PubMed]
- Ghanbarabadi, M.; Iranshahi, M.; Amoueian, S.; Mehri, S.; Motamedshariaty, V.S.; Mohajeri, S.A. Neuroprotective and memory enhancing effects of auraptene in a rat model of vascular dementia: Experimental study and histopathological evaluation. Neurosci. Lett. 2016, 623, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Epifano, F.; Molinaro, G.; Genovese, S.; Ngomba, R.T.; Nicoletti, F.; Curini, M. Neuroprotective effect of prenyloxycoumarins from edible vegetables. Neurosci. Lett. 2008, 443, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Genovese, S.; Epifano, F.; Curini, M.; Dudra-Jastrzebska, M.; Luszczki, J.J. Prenyloxyphenylpropanoids as a novel class of anticonvulsive agents. Bioorg. Med. Chem. Lett. 2009, 19, 5419–5422. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Minami, S.; Shimada, N.; Makihata, N.; Nakajima, M.; Furukawa, Y. Anti-inflammatory and neuroprotective effects of auraptene, a citrus coumarin, following cerebral global ischemia in mice. Eur. J. Pharmacol. 2013, 699, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Morita, M.; Kaji, M.; Amakura, Y.; Yoshimura, M.; Shimamoto, K.; Ookido, Y.; Nakajima, M.; Furukawa, Y. Auraptene acts as an anti-inflammatory agent in the mouse brain. Molecules 2015, 20, 20230–20239. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Yamamoto, K.; Mori, H.; Toyoda, N.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Sugawara, K.; Sudo, M.; Nakajima, M.; et al. Auraptene in the peels of Citrus kawachiensis (Kawachi Bankan) ameliorates lipopolysaccharide-induced inflammation in the mouse brain. Evid. Based Complement. Altern. Med. 2014, 2014, 408503. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zheng, Z.; Ruan, J.; Li, Z.; Tzeng, C.-M. Chronic inflammation links cancer and Parkinson’s disease. Front. Aging Neurosci. 2016, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s disease: Its role in neuronal deatu and implications for therapeutic intervention. Neurobiol. Dis. 2009, 37, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Datla, K.P.; Zbarsky, V.; Rai, D.; Parkar, S.; Osakabe, N.; Aruoma, O.I.; Dexter, D.T. Short-term supplementation with plant extracts rich in flavonoids protect nigrostriatal dopaminergic neurons in a rat model of Parkinson’s disease. J. Am. Coll. Nutr. 2007, 26, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Vila, M.; Przedborski, S. Targeting programmed cell death in neurodegenerative diseases. Nat. Rev. Neurosci. 2003, 4, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Furuya, T.; Yasuda, T.; Miura, M.; Mizuno, Y.; Mochizuki, H. Neurotoxic effects of lipopolysaccharide on nigral dopaminergic neurons are mediated by microglial activation, interleukin-1β, and expression of caspase-11 in mice. J. Biol. Chem. 2004, 279, 51647–51653. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Venderova, K.; Park, D.S.; Cai, H.; Schmidt, E. Animal models of Parkinson’s disease. Parkinsons Dis. 2011, 2011, 364328. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.; Herrera, A.J.; Venero, J.L.; Santiago, M.; de Pablos, R.M.; Villarán, R.F.; Espinosa-Oliva, A.M.; Argüelles, S.; Sarmiento, M.; Delgado-Cortés, M.J.; et al. Inflammatory animal model for Parkinson’s disease: The intranigral injection of LPS induced the inflammatory process along with the selective degeneration of nigrostriatal dopaminergic neurons. ISRN Neurol. 2011, 2011, 476158. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.D.; Ksiazek, K.; Menezes, R. Oxidative stress in neurodegenerative disease and ageing. Oxid. Med. Cell. Longev. 2012, 2012, 796360. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Nehru, B. Characterization of the lipopolysaccharide induced model of Parkinson’s disease: Role of oxidative stress and neuroinflammation. Neurochem. Int. 2015, 87, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.; Jin, Y.; Mizuno, A.; Suzuki, H.; Okada, Y.; Takasuka, N.; Tokuda, H.; Nishino, H.; Okuyama, T. Studies on cancer chemoprevention by traditional folk medicines XXIV. Inhibitory effect of a coumarin derivative, 7-isopentenyloxycoumarin, against tumor-promotion. Biol. Pharm. Bull. 2002, 25, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Epifano, F.; Pelucchini, C.; Curini, M.; Genovese, S. Insights on novel biologically active natural products: 7-Isopentenyloxycoumarin. Nat. Prod. Commun. 2009, 4, 1755–1760. [Google Scholar] [PubMed]
- Curini, M.; Epifano, F.; Genovese, S.; Marcotullio, M.C.; Menghini, L. 3-(4′-geranyloxy-3′-methoxyphenyl)-2-trans propenoic acid: A novel promising cancer chemopreventive agent. Anticancer Agents Med. Chem. 2006, 6, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Genovese, S.; Curini, M.; Gresele, P.; Corazzi, T.; Epifano, F. Inhibition of COX-1 activity and COX-2 expression by 3-(4′-geranyloxy-3′-methoxyphenyl)-2-trans propenoic acid and its semi-synthetic derivatives. Bioorg. Med. Chem. Lett. 2011, 21, 5995–5998. [Google Scholar] [CrossRef] [PubMed]
- Genovese, S.; Epifano, F.; Carlucci, G.; Marcotullio, M.C.; Curini, M.; Locatelli, M. Quantification of 4′-geranyloxyferulic acid, a new natural colon cancer chemopreventive agent, by HPLC-DAD in grapefruit skin extract. J. Pharm. Biomed. Anal. 2010, 53, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Kohno, H.; Suzuki, R.; Curini, M.; Epifano, F.; Maltese, F.; Gonzales, S.P.; Tanaka, T. Dietary administration with prenyloxycoumarins, auraptene and collinin, inhibits colitis-related colon carcinogenesis in mice. Int. J. Cancer 2006, 118, 2936–2942. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Benveniste, E.N. Immune function of astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Nilsson, M. Astrocyte activation and reactive gliosis. Glia 2005, 50, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Whitton, P.S. Inflammation as a causative factor in the aetiology of Parkinson’s disease. Br. J. Pharmacol. 2007, 150, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Brundin, P.; Li, J.Y.; Holton, J.L.; Lindvall, O.; Revesz, T. Research in motion: The enigma of Parkinson’s disease pathology spread. Nat. Rev. Neurosci. 2008, 9, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Won, S.J.; Kim, D.Y.; Gwag, B.J. Cellular and molecular pathways of ischemic neuronal death. J. Biochem. Mol. Biol. 2002, 35, 67–86. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.E.; Gendelman, H.E. Immunomodulation as a neuroprotective and therapeutic strategy for Parkinson’s disease. Curr. Opin. Pharmacol. 2016, 26, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ludka, F.K.; Dal-Cim, T.; Binder, L.B.; Constantino, L.C.; Massari, C.; Tasca, C.I. Atorvastatin and fluoxetine prevent oxidative stress and mitochondrial dysfunction evoked by glutamate toxicity in hippocampal slices. Mol. Neurobiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Kobayashi, M. The role for oxidative stress in neurodegenerative disease. Brain Nerve 2008, 60, 157–170. [Google Scholar] [PubMed]
- Niranjan, R. The role of inflammatory and oxidative stress mechanisms in the pathogenesis of Parkinson’s disease: Focus on astrocytes. Mol. Neurobiol. 2014, 49, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Rickert, U.; Grampp, S.; Wilms, H.; Spreu, J.; Knerlich-Lukoschus, F.; Held-Feindt, J.; Lucius, R. Glial cell line-derived neurotrophic factor family members reduce microglial activation via inhibiting p38MAPKs-mediated inflammatory responses. J. Neurodegener. Dis. 2014, 2014, 369468. [Google Scholar] [CrossRef] [PubMed]
- Bruyere, C.; Genovese, S.; Lallemand, B.; Ionescu-Motatu, A.; Curini, M.; Kiss, R.; Epifano, F. Growth inhibitory activities of oxyprenylated and non-prenylated naturally occurring phenylpropanoids in cancer cell lines. Bioorg. Med. Chem. Lett. 2011, 21, 4173–4178. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okuyama, S.; Semba, T.; Toyoda, N.; Epifano, F.; Genovese, S.; Fiorito, S.; Taddeo, V.A.; Sawamoto, A.; Nakajima, M.; Furukawa, Y. Auraptene and Other Prenyloxyphenylpropanoids Suppress Microglial Activation and Dopaminergic Neuronal Cell Death in a Lipopolysaccharide-Induced Model of Parkinson’s Disease. Int. J. Mol. Sci. 2016, 17, 1716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101716
Okuyama S, Semba T, Toyoda N, Epifano F, Genovese S, Fiorito S, Taddeo VA, Sawamoto A, Nakajima M, Furukawa Y. Auraptene and Other Prenyloxyphenylpropanoids Suppress Microglial Activation and Dopaminergic Neuronal Cell Death in a Lipopolysaccharide-Induced Model of Parkinson’s Disease. International Journal of Molecular Sciences. 2016; 17(10):1716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101716
Chicago/Turabian StyleOkuyama, Satoshi, Tomoki Semba, Nobuki Toyoda, Francesco Epifano, Salvatore Genovese, Serena Fiorito, Vito Alessandro Taddeo, Atsushi Sawamoto, Mitsunari Nakajima, and Yoshiko Furukawa. 2016. "Auraptene and Other Prenyloxyphenylpropanoids Suppress Microglial Activation and Dopaminergic Neuronal Cell Death in a Lipopolysaccharide-Induced Model of Parkinson’s Disease" International Journal of Molecular Sciences 17, no. 10: 1716. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17101716