
Methodological Challenges in Protein Microarray and Immunohistochemistry for the Discovery of Novel Autoantibodies in Paediatric Acute Disseminated Encephalomyelitis

Abstract
:1. Introduction
2. Results
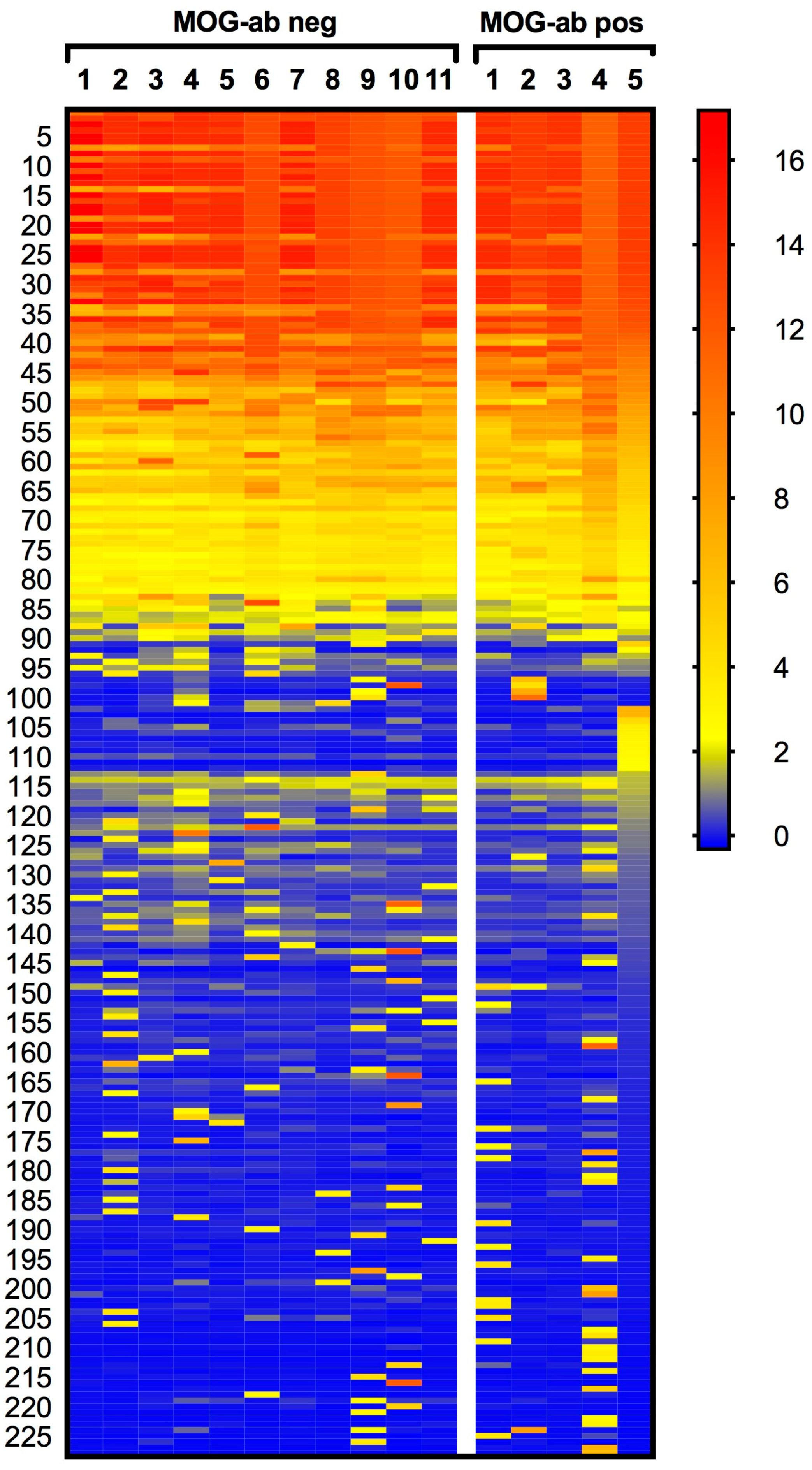
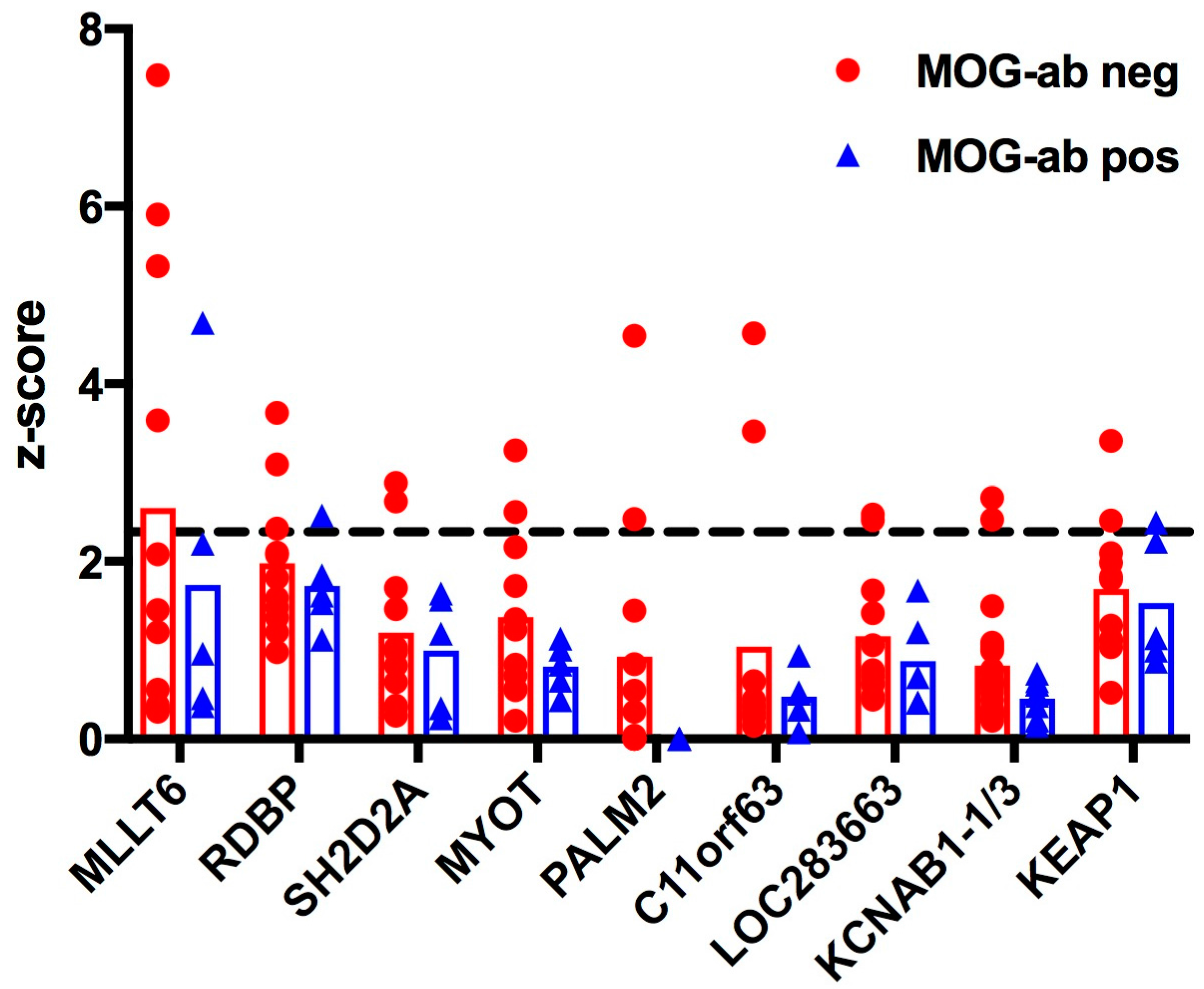
2.1. Protein Microarray Did Not Discriminate between Paediatric ADEM Patients with and without MOG Antibodies
2.2. Rat Brain Immunohistochemistry Revealed No Potential Novel Target in Paediatric ADEM
2.3. Sera of Paediatric MOG Antibody Negative ADEM Patients Did Not Contain Antibodies against Leucine-Rich Glioma-Inactivated 1 (LGI1) and Contactin-Associated Protein-2 (CASPR2)
2.4. Antibodies against Recombinant Myelin Proteins/Peptides Did Not Reveal Significant Differences between the Two Groups
3. Discussion
4. Materials and Methods
4.1. Patients and Samples
4.2. Protein Microarray
4.3. Rat Brain Immunohistochemistry
4.4. Immunocytochemistry
4.5. Enzyme-Linked Immunosorbent Assay (ELISA) to Myelin Antigens
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ADEM | acute disseminated encephalomyelitis |
| AMPAR | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor |
| AQP4 | aquaporin-4 |
| CASPR2 | contactin-associated protein-2 |
| CI | confidence interval |
| CI-P | Chebyshev’s inequality principle |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| ELISA | enzyme-linked immunosorbent assay |
| FITC | fluorescein isothiocyanate |
| GAD | glutamic acid decarboxylase |
| Ig | Immunoglobulin |
| IHC | Immunohistochemistry |
| IPMSSG | International Paediatric Multiple Sclerosis Study Group |
| KCNAB1 | voltage-gated potassium channel subunit beta-1 |
| KCNAB2 | voltage-gated potassium channel subunit beta member-2 |
| KCTD6 | potassium channel tetramerisation domain containing 6 |
| KCTD18 | potassium channel tetramerisation domain containing 18 |
| LGI1 | leucine-rich glioma-inactivated 1 |
| MBP | myelin basic protein |
| MOG | myelin oligodendrocyte glycoprotein |
| MS | multiple sclerosis |
| NMO | neuromyelitis optica |
| NMOSD | NMO spectrum disorders |
| OD | optical density |
| PFA | paraformaldehyde |
| PLP | proteolipid protein |
| rhMOG | recombinant human MOG |
| v | Variant |
| VGKC | voltage-gated potassium channel |
References
- Krupp, L.B.; Tardieu, M.; Amato, M.P.; Banwell, B.; Chitnis, T.; Dale, R.C.; Ghezzi, A.; Hintzen, R.; Kornberg, A.; Pohl, D.; et al. International pediatric multiple sclerosis study group criteria for pediatric multiple sclerosis and immune-mediated central nervous system demyelinating disorders: Revisions to the 2007 definitions. Mult. Scler. 2013, 19, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Tenembaum, S.N. Acute disseminated encephalomyelitis. Handb. Clin. Neurol. 2013, 112, 1253–1262. [Google Scholar] [PubMed]
- Brilot, F.; Dale, R.C.; Selter, R.C.; Grummel, V.; Kalluri, S.R.; Aslam, M.; Busch, V.; Zhou, D.; Cepok, S.; Hemmer, B. Antibodies to native myelin oligodendrocyte glycoprotein in children with inflammatory demyelinating central nervous system disease. Ann. Neurol. 2009, 66, 833–842. [Google Scholar] [CrossRef] [PubMed]
- O'Connor, K.C.; McLaughlin, K.A.; De Jager, P.L.; Chitnis, T.; Bettelli, E.; Xu, C.; Robinson, W.H.; Cherry, S.V.; Bar-Or, A.; Banwell, B.; et al. Self-antigen tetramers discriminate between myelin autoantibodies to native or denatured protein. Nat. Med. 2007, 13, 211–217. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, K.A.; Chitnis, T.; Newcombe, J.; Franz, B.; Kennedy, J.; McArdel, S.; Kuhle, J.; Kappos, L.; Rostasy, K.; Pohl, D.; et al. Age-dependent B cell autoimmunity to a myelin surface antigen in pediatric multiple sclerosis. J. Immunol. 2009, 183, 4067–4076. [Google Scholar] [CrossRef] [PubMed]
- Probstel, A.K.; Dornmair, K.; Bittner, R.; Sperl, P.; Jenne, D.; Magalhaes, S.; Villalobos, A.; Breithaupt, C.; Weissert, R.; Jacob, U.; et al. Antibodies to MOG are transient in childhood acute disseminated encephalomyelitis. Neurology 2011, 77, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Di Pauli, F.; Mader, S.; Rostasy, K.; Schanda, K.; Bajer-Kornek, B.; Ehling, R.; Deisenhammer, F.; Reindl, M.; Berger, T. Temporal dynamics of anti-MOG antibodies in cns demyelinating diseases. Clin. Immunol. 2011, 138, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Hacohen, Y.; Absoud, M.; Deiva, K.; Hemingway, C.; Nytrova, P.; Woodhall, M.; Palace, J.; Wassmer, E.; Tardieu, M.; Vincent, A.; et al. Myelin oligodendrocyte glycoprotein antibodies are associated with a non-MS course in children. Neurol. Neuroimmunol. Neuroinflamm. 2015, 2, e81. [Google Scholar] [CrossRef] [PubMed]
- Dale, R.C.; Tantsis, E.M.; Merheb, V.; Kumaran, R.Y.; Sinmaz, N.; Pathmanandavel, K.; Ramanathan, S.; Booth, D.R.; Wienholt, L.A.; Prelog, K.; et al. Antibodies to MOG have a demyelination phenotype and affect oligodendrocyte cytoskeleton. Neurol. Neuroimmunol. Neuroinflamm. 2014, 1, e12. [Google Scholar] [CrossRef] [PubMed]
- Ketelslegers, I.A.; Van Pelt, D.E.; Bryde, S.; Neuteboom, R.F.; Catsman-Berrevoets, C.E.; Hamann, D.; Hintzen, R.Q. Anti-MOG antibodies plead against ms diagnosis in an acquired demyelinating syndromes cohort. Mult. Scler. 2015, 21, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Carbonell, C.; Vargas-Lowy, D.; Musallam, A.; Healy, B.; McLaughlin, K.; Wucherpfennig, K.W.; Chitnis, T. Clinical and MRI phenotype of children with MOG antibodies. Mult. Scler. 2016, 22, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Baumann, M.; Sahin, K.; Lechner, C.; Hennes, E.M.; Schanda, K.; Mader, S.; Karenfort, M.; Selch, C.; Hausler, M.; Eisenkolbl, A.; et al. Clinical and neuroradiological differences of paediatric acute disseminating encephalomyelitis with and without antibodies to the myelin oligodendrocyte glycoprotein. J. Neurol. Neurosurg. Psychiatry 2015, 86, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.C.; Breithaupt, C.; Reindl, M.; Schanda, K.; Rostasy, K.; Berger, T.; Dale, R.C.; Brilot, F.; Olsson, T.; Jenne, D.; et al. Distinction and temporal stability of conformational epitopes on myelin oligodendrocyte glycoprotein recognized by patients with different inflammatory central nervous system diseases. J. Immunol. 2013, 191, 3594–3604. [Google Scholar] [CrossRef] [PubMed]
- Hacohen, Y.; Singh, R.; Rossi, M.; Lang, B.; Hemingway, C.; Lim, M.; Vincent, A. Clinical relevance of voltage-gated potassium channel-complex antibodies in children. Neurology 2015, 85, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Hacohen, Y.; Nishimoto, Y.; Fukami, Y.; Lang, B.; Waters, P.; Lim, M.J.; Yuki, N.; Vincent, A. Paediatric brainstem encephalitis associated with glial and neuronal autoantibodies. Dev. Med. Child Neurol. 2016, 58, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Peschl, P.; Reindl, M.; Schanda, K.; Sospedra, M.; Martin, R.; Lutterotti, A. Antibody responses following induction of antigen-specific tolerance with antigen-coupled cells. Mult. Scler. 2014, 21, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Wingerchuk, D.M.; Banwell, B.; Bennett, J.L.; Cabre, P.; Carroll, W.; Chitnis, T.; de Seze, J.; Fujihara, K.; Greenberg, B.; Jacob, A.; et al. International consensus diagnostic criteria for neuromyelitis optica spectrum disorders. Neurology 2015, 85, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Van Haren, K.; Tomooka, B.H.; Kidd, B.A.; Banwell, B.; Bar-Or, A.; Chitnis, T.; Tenembaum, S.N.; Pohl, D.; Rostasy, K.; Dale, R.C.; et al. Serum autoantibodies to myelin peptides distinguish acute disseminated encephalomyelitis from relapsing-remitting multiple sclerosis. Mult. Scler. 2013, 19, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Kaliakatsos, M.; Hacohen, Y.; Siddiqui, A.; Dlamini, N.; Vincent, A.; Lim, M. Acute disseminated encephalomyelitis associated with positive voltage gated potassium channel complex antibody. Mult. Scler. Relat. Disord. 2013, 2, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Querol, L.; Clark, P.L.; Bailey, M.A.; Cotsapas, C.; Cross, A.H.; Hafler, D.A.; Kleinstein, S.H.; Lee, J.Y.; Yaari, G.; Willis, S.N.; et al. Protein array-based profiling of CSF identifies RBPJ as an autoantigen in multiple sclerosis. Neurology 2013, 81, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Berger, T.; Rubner, P.; Schautzer, F.; Egg, R.; Ulmer, H.; Mayringer, I.; Dilitz, E.; Deisenhammer, F.; Reindl, M. Antimyelin antibodies as a predictor of clinically definite multiple sclerosis after a first demyelinating event. N. Engl. J. Med. 2003, 349, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Kuhle, J.; Pohl, C.; Mehling, M.; Edan, G.; Freedman, M.S.; Hartung, H.P.; Polman, C.H.; Miller, D.H.; Montalban, X.; Barkhof, F.; et al. Lack of association between antimyelin antibodies and progression to multiple sclerosis. N. Engl. J. Med. 2007, 356, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Lang, B.; Makuch, M.; Moloney, T.; Dettmann, I.; Mindorf, S.; Probst, C.; Stoecker, W.; Buckley, C.; Newton, C.R.; Leite, M.I.; et al. Intracellular and non-neuronal targets of voltage-gated potassium channel complex antibodies. J. Neurol. Neurosurg. Psychiatry 2017. [Google Scholar] [CrossRef] [PubMed]
- Graus, F.; Titulaer, M.J.; Balu, R.; Benseler, S.; Bien, C.G.; Cellucci, T.; Cortese, I.; Dale, R.C.; Gelfand, J.M.; Geschwind, M.; et al. A clinical approach to diagnosis of autoimmune encephalitis. Lancet Neurol. 2016, 15, 391–404. [Google Scholar] [CrossRef]
- Jarius, S.; Ruprecht, K.; Kleiter, I.; Borisow, N.; Asgari, N.; Pitarokoili, K.; Pache, F.; Stich, O.; Beume, L.A.; Hummert, M.W.; et al. MOG-IgG in NMO and related disorders: A multicenter study of 50 patients. Part 1: Frequency, syndrome specificity, influence of disease activity, long-term course, association with AQP4-IgG, and origin. J. Neuroinflamm. 2016, 13, 279. [Google Scholar] [CrossRef] [PubMed]
- Hoftberger, R.; Sabater, L.; Marignier, R.; Aboul-Enein, F.; Bernard-Valnet, R.; Rauschka, H.; Ruiz, A.; Blanco, Y.; Graus, F.; Dalmau, J.; et al. An optimized immunohistochemistry technique improves NMO-IgG detection: Study comparison with cell-based assays. PLoS ONE 2013, 8, e79083. [Google Scholar] [CrossRef] [PubMed]
- Reindl, M.; Linington, C.; Brehm, U.; Egg, R.; Dilitz, E.; Deisenhammer, F.; Poewe, W.; Berger, T. Antibodies against the myelin oligodendrocyte glycoprotein and the myelin basic protein in multiple sclerosis and other neurological diseases: A comparative study. Brain J. Neurol. 1999, 122 Pt 11, 2047–2056. [Google Scholar] [CrossRef]
- Rauer, S.; Euler, B.; Reindl, M.; Berger, T. Antimyelin antibodies and the risk of relapse in patients with a primary demyelinating event. J. Neurol. Neurosurg. Psychiatry 2006, 77, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.; Reindl, M.; Lutterotti, A.; Kuenz, B.; Ehling, R.; Gneiss, C.; Lackner, P.; Deisenhammer, F.; Berger, T. Epitope specificity of serum antibodies directed against the extracellular domain of myelin oligodendrocyte glycoprotein: Influence of relapses and immunomodulatory treatments. J. Neuroimmunol. 2006, 174, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Pittock, S.J.; Reindl, M.; Achenbach, S.; Berger, T.; Bruck, W.; Konig, F.; Morales, Y.; Lassmann, H.; Bryant, S.; Moore, S.B.; et al. Myelin oligodendrocyte glycoprotein antibodies in pathologically proven multiple sclerosis: Frequency, stability and clinicopathologic correlations. Mult. Scler. 2007, 13, 7–16. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Protein Symbol | Protein Name and Known Function | MOG Antibody Negative | MOG Antibody Positive |
|---|---|---|---|
| MLLT6 | Myeloid/lymphoid or mixed-lineage leukaemia, translocated to, 6 | 4 | 1 |
| RDBP | RNA binding protein, component of negative elongation factor E (NELF-E) of transcription | 3 | 1 |
| SH2D2A | SH2 domain containing 2A, adaptor protein thought to function in T cell signal transduction | 2 | 0 |
| MYOT | Myotilin, stability of thin filaments during muscle contraction | 2 | 0 |
| PALM2 | Paralemmin-2, implicated in plasma membrane dynamics in neurons and other cell types | 2 | 0 |
| C11orf63 | Chromosome 11 open reading frame 63, protein not characterized | 2 | 0 |
| LOC283663 | Predicted: Homo sapiens hypothetical protein LOC283663, protein not characterized | 2 | 0 |
| KCNAB1 v1/v3 | Voltage-gated potassium channel subunit beta-1, variants 1 and 3 | 2 | 0 |
| KEAP1 | Kelch-like ECH-associated protein 1, regulates antioxidant response and ubiquitination | 2 | 1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peschl, P.; Ramberger, M.; Höftberger, R.; Jöhrer, K.; Baumann, M.; Rostásy, K.; Reindl, M. Methodological Challenges in Protein Microarray and Immunohistochemistry for the Discovery of Novel Autoantibodies in Paediatric Acute Disseminated Encephalomyelitis. Int. J. Mol. Sci. 2017, 18, 679. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18030679
Peschl P, Ramberger M, Höftberger R, Jöhrer K, Baumann M, Rostásy K, Reindl M. Methodological Challenges in Protein Microarray and Immunohistochemistry for the Discovery of Novel Autoantibodies in Paediatric Acute Disseminated Encephalomyelitis. International Journal of Molecular Sciences. 2017; 18(3):679. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18030679
Chicago/Turabian StylePeschl, Patrick, Melanie Ramberger, Romana Höftberger, Karin Jöhrer, Matthias Baumann, Kevin Rostásy, and Markus Reindl. 2017. "Methodological Challenges in Protein Microarray and Immunohistochemistry for the Discovery of Novel Autoantibodies in Paediatric Acute Disseminated Encephalomyelitis" International Journal of Molecular Sciences 18, no. 3: 679. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18030679