
The Roles of Glutamine in the Intestine and Its Implication in Intestinal Diseases



1
Food Science and Human Nutrition Department, Center for Nutritional Sciences, College of Agricultural and Life Sciences, University of Florida, Gainesville, FL 32611, USA
2
Department of Food and Nutrition, Brain Korea 21 PLUS Project, College of Human Ecology, Yonsei University, Seoul 03722, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(5), 1051; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18051051
Submission received: 25 March 2017
/
Revised: 9 May 2017
/
Accepted: 10 May 2017
/
Published: 12 May 2017
(This article belongs to the Special Issue Nutrigenomics of Risk Factors for Disease)
Abstract
:Glutamine, the most abundant free amino acid in the human body, is a major substrate utilized by intestinal cells. The roles of glutamine in intestinal physiology and management of multiple intestinal diseases have been reported. In gut physiology, glutamine promotes enterocyte proliferation, regulates tight junction proteins, suppresses pro-inflammatory signaling pathways, and protects cells against apoptosis and cellular stresses during normal and pathologic conditions. As glutamine stores are depleted during severe metabolic stress including trauma, sepsis, and inflammatory bowel diseases, glutamine supplementation has been examined in patients to improve their clinical outcomes. In this review, we discuss the physiological roles of glutamine for intestinal health and its underlying mechanisms. In addition, we discuss the current evidence for the efficacy of glutamine supplementation in intestinal diseases.
{kind=link}
{kind=link}
1. Introduction
Glutamine is the most abundant amino acid in human blood, skeletal muscle, and the free amino acid pool [1]. It plays physiologically important roles in various metabolic processes: as an intermediary in energy metabolism, and as a substrate for the synthesis of peptides and non-peptides such as nucleotide bases, glutathione, and neurotransmitters [2,3,4]. Additionally, glutamine contributes to the detoxification of ammonia and systemic acid-base balance [5]. The involvement of glutamine metabolism in immune systems [1,6,7] and in cancer cells [8,9,10] has been documented in the past two decades. Moreover, glutamine metabolism has direct relevance to clinical medicine. This was initially highlighted when glutamine, classically a non-essential amino acid, was considered to be conditionally essential during certain catabolic states, such as trauma or sepsis [11]. The theory of “conditionally essential” glutamine during illness was based on observations that intestinal, renal, and immune cells utilize large amounts of glutamine, exceeding the endogenous glutamine production [12,13], and that plasma and muscle glutamine levels are markedly reduced in these conditions [14].
Among the various tissues using glutamine at high rates, the intestine utilizes about 30% of total glutamine [15], indicating that it is a key nutrient for the intestine. Studies in healthy adults have demonstrated that three quarters of enterally provided glutamine is absorbed into the splanchnic tissues, and most of the absorbed glutamine is metabolized within the intestine [16,17]. One-fourth of the plasma glutamine is taken up by the small intestine when it passes the organ [18]. It has been reported that the intestine competes with other tissues for glutamine from the body amino acid pool and dietary sources [19]. Glutamine metabolism in the intestine has been intensively studied. Its functions include maintaining nucleotide metabolism and intestinal barrier function, modulation of inflammation, and regulating stress responses and apoptosis [20,21,22]. Concomitantly, the efficacy of glutamine supplementation has been tested in humans and animal models with intestinal diseases [23,24,25]. This review aims to discuss the role of glutamine in the intestine, and to summarize current evidence for the clinical efficacy of glutamine supplementation in intestinal diseases, especially inflammatory bowel diseases (IBDs).
2. Roles of Glutamine in the Intestine
2.1. Tissue Integrity
The mammalian intestinal lumen is lined with a single layer of epithelial cells [26]. As these cells are renewed every four to five days, a continuously high level of cell proliferation is required to maintain homeostasis [27]. In general, cell proliferation is regulated by a number of signaling pathways and hormones such as growth factors. When proliferation is activated by these signals, crypt-residing intestinal stem cells differentiate into specialized epithelial cell types, including enterocytes, goblet cells, paneth cells, and enterocytes, which enables the maintenance of normal intestinal tissue integrity [28].
Glutamine influences a number of signaling pathways that regulate cell cycle regulation and proliferation. Mitogen-activated protein kinases (MAPKs) are protein kinases that orchestrate a number of cell functions, including cell proliferation and differentiation [29]. Rhoads et al. demonstrated that glutamine is required for intestinal cell proliferation by activating multiple MAPKs, including extracellular signal-regulated kinases (ERK1/2) and c-Jun N-terminal kinases (JNK1/2), in the rat intestinal mucosal cell line, IEC-6 [30] (Figure 1). Additionally, glutamine contributes to intestinal cell proliferation by augmenting the effects of growth factors such as epidermal growth factor (EGF), insulin-like growth factor-I (IGF-I), and transforming growth factor-α (TGF-α). Restriction of glutamine in cell culture media resulted in impaired EGF-stimulation of DNA, RNA, and protein synthesis and cell replication in IEC-6 cells [31]. Consumption of glutamine-enriched diets significantly enhanced IGF-I-mediated DNA and protein synthesis in a rat model of short bowel syndrome [32]. In a porcine model of ischemia, glutamine administration enhanced the action of TGF-α on mucosal cell proliferation [33].
Tight junctions, consisting of various proteins, seal adjacent epithelial cells to produce a physical barrier between epithelial and endothelial cells [34]. Tight junctions are dynamic rather than static structures. Indeed, tight junctions constantly remodel their structures with a relatively high rate of turnover to interact with external stimuli by which they control the entry of ions, nutrients, and water [35]. In addition, tight junctions maintain intestinal integrity, which prevents pathogens and toxins from entering the intestinal lumen [36]. There are four types of transmembrane components of tight junctions, including claudins, occludin, tricellulin, and junctional adhesion molecules [37]. In response to various physiological stimuli and signal pathways, tight junctions modulate the transport of luminal molecules into mucosal cells by adjusting their tightness [38]. These signaling molecules are protein kinase C, MAPKs, and myosin light chain kinases (MLCK). Activation of protein kinase C resulted in the upregulation of occludin, zonula occudens (ZO)-1, ZO-2, and claudin 1 in primary human epithelial cells, leading to enhancing transepithelial electrical resistance [39]. MAPKs could directly interact with the C-terminal tail of occludin, which mediates the prevention of hydrogen peroxide-induced disruption of tight junctions [40]. Furthermore, MLCK-induced phosphorylation of myosin light chain regulated tight junction permeability in Caco-2 cells [41,42].
Multiple lines of evidence indicate that glutamine modulates the expression of tight junction proteins. In the human colon carcinoma cell line Caco-2, glutamine deprivation markedly reduced the expression of multiple tight junction proteins, including claudin-1, occludin, and ZO-1 [43]. Restriction of glutamine in cell culture media significantly increased epithelial cell permeability in Caco-2 cells, as determined by multiple methods including transepithelial electrical resistance (TER), tracer models using 14C-mannitol, and fluorescein isothiocyanate-dextran [44]. The addition of glutamine in glutamine-deprived cells rescued the impaired barrier functions. In methotrexate-treated Caco-2 cells, the addition of glutamine improved permeability along with increased ZO-1 and occludin expressions [45]. By measuring TER and inulin flux, Seth et al. also reported that the addition of glutamine prevented acetaldehyde-induced disruption of tight junctions and impaired paracellular permeability [46]. Maintaining intestinal permeability by tight junction proteins has shown to be beneficial for treatment of multiple intestinal pathologic conditions such as inflammatory bowel disease and celiac disease [47]. These studies suggest that glutamine supplementation may be beneficial for individuals with an impaired gut permeability by enhancing the expression of tight junction proteins.
Mechanistically, glutamine has been shown to influence a number of signaling pathways that regulate the expression of tight junction components [48], although much of this process still remains unclear. In the Caco-2 cells, deprivation of glutamine activated the phosphatidylinositol 3-kinase (PI3K)/Akt pathway, which led to a reduction in claudin-1 expression and TER [49]. On the contrary, glutamine supplementation reduced the activation of the PI3K/Akt pathway, which reversed claudin-1 expression in the cells, suggesting that glutamine supplementation regulates phosphorylation states of tight junction proteins. Interestingly, normal formation of tight junction is controlled by phosphorylation of occludin and ZO-1 [50,51,52]. Therefore, glutamine-mediated tight junction maintenance is in part mediated by phosphorylation of tight junction proteins. Wang et al. showed that glutamine activated calcium/calmodulin-dependent kinase 2-AMP-activated protein kinase signaling in porcine jejunal enterocytes [53]. They also reported that glutamine induced the expression of ZO-1, ZO-2, and ZO-3 and caused greater distribution of claudin-1, claudin-4, and ZO-1 at the plasma membranes [53]. Dokladny et al. reported that heat shock-induced activation of heat shock factor-1 (HSF-1) induced nuclear translocation of HSF-1 and expression of occluding in Caco-2 cells [54].
2.2. Inflammatory Pathway
Inflammation has been shown to be a cause of intestinal diseases, such as ulcerative colitis, Crohn’s disease, and colorectal cancer [55]. Therefore, treatment of intestinal inflammation is important to target these diseases. Several lines of evidence indicate that glutamine has an anti-inflammatory property by influencing a number of inflammatory signaling pathways, including the nuclear factor κB (NF-κB) and signal transducer and activator of transcription (STAT) pathways [56].
The transcription factor NF-κB regulates a number of immune responses [57]. During the early phase of infection, it activates various genes to boost the inflammatory reaction. NF-κB is a multiprotein complex composed of five members of the Rel family: p50, p52, p65, RelB, and c-Rel. Under steady-state conditions, NF-κB resides in the cytoplasm and is maintained inactive by a family of inhibitors, designated inhibitor of κB (IκB). In response to various extracellular stimuli, IκB kinase phosphorylates IκB proteins, triggering their degradation and release from NF-κB, which thus becomes active. Active NF-κB complex is translocated into the nucleus where it induces the expression of genes harboring NF-κB-binding elements, such as interleukin-6 (Il-6) and tumor necrosis factor-α (Tnf-α). During inflammatory status, productions of inflammatory cytokines IL-6 and TNF-α are highly elevated, which stimulates immune response by activating multiple target cells such as antigen-presenting cells and T cells and by inducing acute-phase proteins [58]. Glutamine has been shown to suppress NF-κB pathway activation. Administration of glutamine via intraperitoneal injection or oral gavage suppressed NF-κB activation in rodent models of colitis [59,60] and in lipopolysaccharide (LPS)-treated piglet enterocytes [61]. Mechanistically, NF-κB suppression by glutamine was associated with increased expression of cellular heat shock proteins (HSPs) such as HSP25 and HSP70, which are induced by HSF-1 [60]. The HSF-1-mediated heat shock response has been shown to inhibit NF-κB activation and NF-κB-dependent gene expression. This hypothesis was supported by a marked increase in NF-κB activation in embryonic fibroblast from HSF-1-null mutant mice due to lack of HSPs expression [62]. These studies suggest that the anti-inflammatory effect of glutamine, in part, may be related to HSF-1 activation and suppression of NF-κB-mediated inflammatory cytokine expression. Additionally, glutamine influences IκB stability. In LPS-treated Caco-2 cells, deprivation of glutamine reduced the expression of IκBα, triggering elevated NF-κB binding to DNA as well as increased expression of inflammatory cytokine interleukin-8 (IL-8) [63]. In human ileocecal adenocarcinoma HCT-8 cells, pre-treatment with glutamine reduced the level of IκBα degradation and production of IL-8 during TNF-α-induced inflammation [64]. In support of these findings in vitro, Kretzmann et al. showed that glutamine supplementation for seven days significantly reduced IκBα degradation, leading to suppression of NF-κB activation in a rat colitis model [65]. Since production of IL-8, a cytokine that stimulates migration of neutrophils to inflammatory sites, was influenced by glutamine status [63,64], glutamine-mediated IL-8 regulation could be an important event for targeting intestinal inflammation.
STAT proteins are transcription factors that modulate the immune system, cellular proliferation, and development [66]. They have been extensively studied for their roles in regulating inflammation by inducing the expression of cytokines including IL-6 [67]. In rat colitis models, glutamine administration via the rectal route reduced the phosphorylation of STAT1 and STAT5, indicating that glutamine influences STAT signaling activation [65]. In LPS-treated Caco-2 cells, glutamine depletion upregulated STAT4, whereas glutamine supplementation downregulated STAT4 expression and IL-8 production [68]. Therefore, the anti-inflammatory effect of glutamine may be contributed to by inhibiting STAT activation and by inhibiting expression of inflammatory cytokines such as IL-6 and IL-8 in intestinal tissues.
Nitric oxide (NO) is synthesized by multiple cells and modulates a variety of cellular signaling pathways, including inflammatory responses [69]. During intestinal inflammation, NO may play a dichotomous role, as both beneficial and harmful effects of NO have been observed [70]. Glutamine is an important regulator of NO synthesis [71,72]. Houdijk et al. reported that the level of whole body plasma nitrate, the stable end-product of NO production, was reduced in rats fed a glutamine-enriched diet [73]. Similarly, in rats with intestinal ischemia-reperfusion injury, glutamine-enriched diet reduced mucosal expression of inducible NO synthase, an inflammatory enzyme, and decreased the plasma NO concentration [74].
Since sustained activation of inflammatory signaling pathways and prolonged production of pro-inflammatory cytokines are critical in the development and progress of intestinal inflammatory diseases, there has been efforts to suppress the production of inflammatory mediators to treat patients with intestinal inflammatory diseases including IBD [75]. Therefore, based on the in vitro and in vivo studies mentioned above, glutamine supplementation could be one promising candidate for treating intestinal inflammatory disorders by inhibiting activation of NF-κB and STAT, and suppressing expression of inflammatory cytokines such as IL-6, TNF-α, and IL-8, and inflammatory enzyme inducible NO synthase.
2.3. Apoptosis and Cellular Stresses
As intestinal epithelial cells have a turnover rate of four to five days, it is critical for these cells to maintain a fine balance between proliferation and apoptosis for normal function [76]. Spontaneous apoptosis in intestinal epithelia is essential for maintaining its normal architecture [77]. However, a number of cellular stresses induced by exogenous agents or by intracellular stimuli including endotoxemia, nutrient deprivation, and lack of growth factor can disturb the balance between proliferation and apoptosis. This imbalance between proliferation and apoptosis triggers intestinal pathologic conditions due to sustained apoptotic cell death [78,79,80,81]. Indeed, loss of epithelial cells in ulcerative colitis and bacterial infection mainly occurs by increased apoptosis in crypts [82,83]. Intestinal inflammatory disorders such as celiac disease and nematode infections are highly associated with increased apoptosis of intestinal epithelial cells [84,85]. Even though the differentiated enterocytes undergo apoptosis which maintains normal gut epithelial function, dysregulated apoptosis is seen in a number of pathological conditions in the gastrointestinal tract. Therefore, it is critical to inhibit apoptosis of intestinal epithelial cells to prevent the intestinal pathologic conditions [86]. Glutamine has been shown to display anti-apoptotic properties in intestine. In rat intestinal epithelial (RIE-1) cells, glutamine deprivation resulted in apoptosis [87]. Similarly, glutamine supplementation effectively reduced toxin-induced apoptosis in human intestinal epithelial T84 cells [88], and sodium laurate-induced apoptosis in RIE-1 cells [89], collectively suggesting that glutamine is critical to suppressing apoptosis.
Further studies have demonstrated the mechanisms underlying the anti-apoptotic capacity of glutamine. First, as a precursor for glutathione (GSH), glutamine prevents apoptosis by maintaining normal cellular redox status. Along with cysteine and glycine, glutamate converted from glutamine produces GSH, an important cellular antioxidant [90]. GSH is present in the cell in both reduced and oxidized forms (GSSG), and the ratio of GSH to GSSG determines the cellular redox potential. Because depletion of GSH induces apoptosis due to excessive oxidative stress [91], normal glutamine metabolism plays a critical role in preventing apoptosis by providing glutamate required for maintaining normal GSH/GSSG ratio.
Secondly, glutamine regulates caspase activation. Caspases are a family of protease enzymes that play important roles in inducing apoptosis [92]. Basically, they are present as a proenzyme, but various stimuli can activate the enzymes through cleavage. In RIE-1 cells, glutamine-deprived cells showed significantly higher caspase-3 activity along with a higher level of apoptosis [93]. Administration of glutamine reduced caspase-3 activity in neonatal piglet enterocytes [61] as well as caspase-8 activity in T84 cells [88].
In addition, glutamine enhances the expression of heat shock proteins (HSPs) [94]. HSPs have been reported to modulate apoptotic cell death by acting as a molecular chaperone, allowing the cells to adapt to stressful conditions [95]. In rats with sepsis, glutamine administration significantly increased the expression of HSP-70 and HSP-25, possibly via increased phosphorylation of heat shock factor (HSF)-1 [96]. The increased expression of HSPs via glutamine supplementation markedly improved the survival rate of rats with sepsis. Ropeleski et al. reported that glutamine enhanced HSF-1-mediated gene expression of Hsp-72 during heat shock in IEC-18 and H4 intestinal cells [97]. On the contrary, glutamine-deficient conditions reduced gene expression of Hsp-72, which resulted in higher caspase-3 activity and apoptotic cell death.
The protective effect of glutamine is also attributed to its role in modulating cellular stress responses such as endoplasmic reticulum (ER) stress and autophagy. The ER is an organelle responsible for protein synthesis, folding, and modification. A number of pathologic conditions, including IBD, disrupt ER function, resulting in ER stress [98]. As extensive ER stress triggers sustained apoptosis and further insults, attenuating ER stress is critical for cell protection and survival. In rats with colitis, administration of glutamine markedly reduced the activation of ER stress markers, such as glucose responsive protein 78, CCAAT/enhancer binding protein (C/EBP) homologous protein, and caspase-12 [99]. These results show that glutamine supplementation reduces ER stress and apoptosis. Supporting these observations, glutamine treatment reduced the activation of ER stress in Caco-2 cells treated with pharmacological ER stress inducers.
Autophagy is a catabolic process activated during a number of metabolic stress conditions, such as nutrient deprivation [100]. Upon activation, autophagy breaks down cellular organelles and proteins to supply them as an energy source. It has been shown that autophagy provides a protective effect against intestinal pathologic conditions. Autophagy-related 16-like 1 (Atg16L1), a gene essential for functional autophagosome, has been implicated in Crohn’s disease [101,102]. Saitoh et al. showed that mice lacking Atg16L1 were more susceptible to induced acute colitis, supporting the importance of autophagy to inhibit intestinal inflammation [103]. Furthermore, mutation of Atg5 and Atg7, autophagy-related genes, in mouse intestinal epithelium resulted in increased production of TNF-α and IL-1β following LPS administration [104]. Additionally, Paneth cells lacking Atg16L1, Atg5, or Atg7 showed impaired secretion of antimicrobial proteins, which confer intestinal protection against pathogens [105]. Since IBD pathogenesis is associated with dysfunction of Paneth cells [106], functional autophagy is essential for intestinal homeostasis and preventing intestinal inflammation. Glutamine has been reported to increase autophagy in intestinal epithelial cells. In Caco-2 and IEC-18 cells, Sakiyama et al. reported that glutamine treatment augmented the number of autophagosomes as well as the level of microtubule-associated protein light chain 3-phospholipid conjugates, which are markers for autophagy activation [107]. As a result, the enhanced autophagy via glutamine treatment suppressed intestinal apoptosis under stress conditions. The authors demonstrated that glutamine regulates autophagy by influencing mechanistic target of rapamycin (mTOR) signaling. The mTOR pathway integrates signals from nutrients status and growth factors to modulate multiple cellular processes, including autophagy [108]. In agreement with this finding, Van Der Vos et al. demonstrated that glutamine synthetase is a target of forkhead box O3 (FOXO3), which is a transcription factor activated during autophagy [109]. FOXO3-mediated glutamine synthesis resulted in mTOR inhibition, which promoted autophagy activation and cellular survival.
3. Clinical Implications for Intestinal Diseases
Given the importance of glutamine in maintaining normal cellular functions, as discussed above, it is not surprising that glutamine supplementation has been considered and examined in the clinical setting, particularly in diseases implying impaired glutamine metabolism. A conditional glutamine deficient status has been postulated in patients with acute critical illness. In these patients, marked reduction in plasma glutamine concentration is possibly due to the consequence of muscle wasting [14]. Regarding intestinal diseases, patients with Crohn’s disease display low plasma and cellular glutamine concentrations, and reduced mucosal glutaminase activity [110]. These observations led to the hypothesis that glutamine supplementation would improve clinical outcomes.
A number of experiments in animals with IBD demonstrated that glutamine supplementation is able to protect the intestinal mucosa, raising the possibility of use of glutamine support in human patients. In mice with dextran sulfate sodium-induced colitis, oral glutamine supplementation (41.7 g/kg diet; 10 days) resulted in mitigated colonic inflammatory reactions [111] as well as increased expression of small-intestinal intraepithelial γδ-T cells [112]. In dextran sulfate sodium-induced rats, administration of glutamine (0.75 g/kg body weight (BW)/d; 7 days) by oral gavage increased HSP25 and HSP70, and reduced bleeding and diarrhea [60]. Rats with trinitrobenzene sulfonic acid-induced colitis that received dietary glutamine supplementation (20 g/kg or 40 g/kg; 2 weeks) showed reduced production of pro-inflammatory cytokines, including TNF-α and IL-8, bacterial translocation, and inflamed lesions [113]. Oral glutamine supplementation (3% in drinking water) ameliorated abdominal radiation-induced mucosal injury and reduced bacterial translocation in gut mucosa of rats [114]. Injection of glutamine (0.75 g/kg BW) in mice with sepsis model ameliorated sepsis-induced inflammatory reactions by modulating intestinal intraepithelial lymphocytes [115,116].
Based on these positive results in animal models, human studies have been conducted in an attempt to support the efficacy of glutamine supplementation in improving disease status. However, only a limited number of studies concluded that glutamine supplementation has a beneficial effect in intestinal diseases. In a systematic review by García-de-Lorenzo et al., glutamine-enriched diets were shown to improve immunologic aspects in trauma patients and to ameliorate mucositis in post-chemotherapy patients [117]. The authors determined how much glutamine is required to observe improved clinical outcomes: 21 g glutamine/day for 28 days for Crohn’s disease, and 42 g/day for 21 days for short bowel syndrome. In a randomized controlled trial, Benjamin et al. reported that glutamine supplementation (0.5 g/kg BW; 2 months) in patients with Crohn’s disease in remission phase reduced the intestinal permeability and morphology [118].
However, a number of studies did not observe any improved outcomes from glutamine supplementation. In recent randomized and controlled trails, termed Scottish Intensive Care Glutamine or Selenium Evaluative Trial (SIGNET), effects of a parental administration of glutamine (0.1 to 0.2 g/kg BW/day) were evaluated in 500 patients with critical illness [119]. However, the SIGNET study did not show any beneficial outcome of glutamine supplementation. More recently, Reducing Death Due to Oxidative Stress (REDOXS), a randomized and controlled trial, examined glutamine supplementation (0.6 to 0.8 g/kg BW/day) in 1223 critically ill patients [120]. Not only was there no effect of glutamine supplementation on rates of organ failure or infectious complication, but also patients who received enteral glutamine treatment showed a trend toward elevated rate of death at 6 months. Similarly, glutamine supplementation on intestinal diseases do not clearly support efficacy of glutamine supplementation. Akobeng et al. examined the effect of glutamine-enriched polymeric diet (8 g/day for 4 weeks) in 18 children with active Crohn’s disease, and found no changes in parameters tested, including intestinal permeability [121]. Similarly, no significant effect of oral glutamine supplements (21 g/day for 4 weeks) was observed in 14 Crohn’s disease patients [122]. In short bowel syndrome patients, six studies have examined the effect of glutamine support, but no improvement in the surrogate parameters was found [123,124,125,126,127,128]. Although some studies showed favorable effects, the clinical efficacy of glutamine supplementation in intestinal diseases remains a controversial issue.
Alpers et al. pointed out limitations of current clinical studies on glutamine supplementation [129], providing some clues for these inconsistent results. The first thing to consider is whether it is true that humans are really glutamine deficient during the diseases. The “conditionally essential glutamine” theory is still a prediction and is uncertain. Because the levels of amino acids including glutamine dynamically and constantly are changed in tissues and plasma, and plasma levels do not always reflect tissue levels, it is challenging to determine whether glutamine is conditionally essential. In addition, the reduced plasma concentration during critical illness is not specific for glutamine, but it occurs for most amino acids [129]. Parenteral glutamine supplementation in critically ill patients did not restore muscle glutamine depletion of the patients, which raises a question for glutamine deficiency in the patients [130]. Furthermore, plasma glutamine level in seriously ill patients did not predict mortality of the patients [131], suggesting that plasma glutamine status may not be correlated with severity of disease. Therefore, an effort to validate a glutamine deficiency state in the pathologic conditions should be preceded before predicting clinical efficacy of glutamine supplementation.
Various experimental designs could have influenced the indefinite results of clinical studies. First, glutamine was administered in two different ways: total parental nutrition and enteral nutrition. Generally, it is stated that enteral nutrition is safer for prolonged period than parental nutrition, whereas parental nutrition is often recognized as being better for achieving targeted calorie requirement, especially in critically ill patients [132]. Route of administration influences the contribution of glutamine [133]. In patients with acute ulcerative colitis, total enteral nutrition was shown to be nutritionally effective as well as produce fewer complications compared to enteral nutrition [134]. Given total parenteral nutrition produces changes in intestinal morphology and function [135], glutamine supplementation via parenteral nutrition might cause complications in intestine. Second, a wide variety of dose, time, and mode of supplementation was used. Dose of glutamine used in the studies varied up to 5-fold [121,122,136], and treatment period varied from 2 days [124] to 8 weeks [128]. As glutamine couples with alanine and glycine, glutamine complex with alanine and glycine is less susceptible to degradation than free glutamine. Many studies used glutamine-containing dipeptides, which might affect the discrepancy of outcomes. Third, a wide range of clinical courses of patients was used in the studies. Short-term glutamine administration during a flare-up phase could give a greater impact on outcomes than other phases. Additionally, a relatively small sample size showed greater efficacy of glutamine supplementation in critically ill patients [137,138,139] and in IBD patients [117,118]. Therefore, a well-controlled clinical trial with a sufficiently sized population would be required to determine the efficacy of glutamine supplementation in intestinal diseases.
4. Conclusions
In this review, we covered the roles of glutamine in the intestine, including the regulation of enterocyte proliferation, maintenance of tight-junction proteins, modulation of inflammatory pathways, such as NF-κB and STAT signaling, and protection against apoptosis and cellular stresses, which are summarized in Figure 1. Although significant progress has been made in uncovering the functions of glutamine, most of these are based on observational studies. Therefore, future research should focus on the mechanisms underlying glutamine actions. Additionally, current data from clinical trials do not support the use of glutamine supplementation in patients with intestinal diseases, despite in vitro and animal model studies having shown significant beneficial effects. Thus, future human studies should be more standardized to increase their power.
Acknowledgments
This work was supported by Brain Korea 21 PLUS Project for Bioactive Nutrition, Yonsei University, Korea.
Author Contributions
Min-Hyun Kim performed the comprehensive literature search and wrote the paper. Hyeyoung Kim performed the initial literature search as well as revised and edited the paper. All authors agree with the edited version.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Calder, P. Glutamine and the immune system. Clin. Nutr. 1994, 13, 2–8. [Google Scholar] [CrossRef]
- Albrecht, J.; Sidoryk-Wegrzynowicz, M.; Zielinska, M.; Aschner, M. Roles of glutamine in neurotransmission. Neuron. Glia Biol. 2010, 6, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Amores-Sanchez, M.I.; Medina, M.A. Glutamine, as a precursor of glutathione, and oxidative stress. Mol. Genet. Metab. 1999, 67, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Coster, J.; McCauley, R.; Hall, J. Glutamine: Metabolism and application in nutrition support. Asia Pac. J. Clin. Nutr. 2004, 13, 25–31. [Google Scholar] [PubMed]
- Patience, J.F. A review of the role of acid-base balance in amino acid nutrition. J. Anim. Sci. 1990, 68, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Curi, R.; Pithon Curi, T.C.; Murphy, C.J.; Garcia, C.; Pires de Melo, M. Glutamine metabolism by lymphocytes, macrophages, and neutrophils: Its importance in health and disease. J. Nutr. Biochem. 1999, 10, 316–324. [Google Scholar] [CrossRef]
- Newsholme, P. Why is l-glutamine metabolism important to cells of the immune system in health, postinjury, surgery or infection? J. Nutr. 2001, 131, 2515S–2522S. [Google Scholar] [PubMed]
- DeBerardinis, R.J.; Cheng, T. Q’s next: The diverse functions of glutamine in metabolism, cell biology and cancer. Oncogene 2010, 29, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of cancer cell metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kim, H. Oncogenes and tumor suppressors regulate glutamine metabolism in cancer cells. J. Cancer Prev. 2013, 18, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Lacey, J.M.; Wilmore, D.W. Is glutamine a conditionally essential amino acid? Nutr. Rev. 1990, 48, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Wischmeyer, P.E. Glutamine: Mode of action in critical illness. Crit. Care Med. 2007, 35, S541–S544. [Google Scholar] [CrossRef] [PubMed]
- Karinch, A.M.; Pan, M.; Lin, C.M.; Strange, R.; Souba, W.W. Glutamine metabolism in sepsis and infection. J. Nutr. 2001, 131, 2535S–2538S. [Google Scholar] [PubMed]
- Askanazi, J.; Carpentier, Y.A.; Michelsen, C.B.; Elwyn, D.H.; Furst, P.; Kantrowitz, L.R.; Gump, F.E.; Kinney, J.M. Muscle and plasma amino acids following injury. Influence of intercurrent infection. Ann. Surg. 1980, 192, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Intestinal mucosal amino acid catabolism. J. Nutr. 1998, 128, 1249–1252. [Google Scholar] [PubMed]
- Newsholme, E.A.; Carrie, A.L. Quantitative aspects of glucose and glutamine metabolism by intestinal cells. Gut 1994, 35, S13–S17. [Google Scholar] [CrossRef] [PubMed]
- Dechelotte, P.; Darmaun, D.; Rongier, M.; Hecketsweiler, B.; Rigal, O.; Desjeux, J.F. Absorption and metabolic effects of enterally administered glutamine in humans. Am. J. Physiol. 1991, 260, G677–G682. [Google Scholar] [PubMed]
- Hankard, R.G.; Darmaun, D.; Sager, B.K.; D’Amore, D.; Parsons, W.R.; Haymond, M. Response of glutamine metabolism to exogenous glutamine in humans. Am. J. Physiol. 1995, 269, E663–E670. [Google Scholar] [PubMed]
- Evans, M.A.; Shronts, E.P. Intestinal fuels: Glutamine, short-chain fatty acids, and dietary fiber. J. Am. Diet. Assoc. 1992, 92, 1239–1246. [Google Scholar] [PubMed]
- Wang, B.; Wu, G.; Zhou, Z.; Dai, Z.; Sun, Y.; Ji, Y.; Li, W.; Wang, W.; Liu, C.; Han, F.; et al. Glutamine and intestinal barrier function. Amino Acids 2015, 47, 2143–2154. [Google Scholar] [CrossRef] [PubMed]
- McCauley, R.; Kong, S.E.; Hall, J. Glutamine and nucleotide metabolism within enterocytes. J. Parenter. Enter. Nutr. 1998, 22, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Glutamine as an immunonutrient. Yonsei Med. J. 2011, 52, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Soeters, P.B.; Grecu, I. Have we enough glutamine and how does it work? A clinician’s view. Ann. Nutr. Metab. 2012, 60, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Novak, F.; Heyland, D.K.; Avenell, A.; Drover, J.W.; Su, X. Glutamine supplementation in serious illness: A systematic review of the evidence. Crit. Care Med. 2002, 30, 2022–2029. [Google Scholar] [CrossRef] [PubMed]
- Bongers, T.; Griffiths, R.D.; McArdle, A. Exogenous glutamine: The clinical evidence. Crit. Care Med. 2007, 35, S545–S552. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.A.; Alison, M. The Biology of Epithelial Cell Populations; Oxford University Press: New York, NY, USA, 1984; Volume 1. [Google Scholar]
- Van der Flier, L.G.; Clevers, H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Bjerknes, M.; Cheng, H. Gastrointestinal stem cells. II. Intestinal stem cells. Am. J. Physiol. Gastrointest. Liv. Physiol. 2005, 289, G381–G387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, J.M.; Argenzio, R.A.; Chen, W.; Rippe, R.A.; Westwick, J.K.; Cox, A.D.; Berschneider, H.M.; Brenner, D.A. l-glutamine stimulates intestinal cell proliferation and activates mitogen-activated protein kinases. Am. J. Physiol. 1997, 272, G943–G953. [Google Scholar] [PubMed]
- Ko, T.C.; Beauchamp, R.D.; Townsend, C.M., Jr.; Thompson, J.C. Glutamine is essential for epidermal growth factor-stimulated intestinal cell proliferation. Surgery 1993, 114, 147–153. [Google Scholar] [PubMed]
- Ziegler, T.R.; Mantell, M.P.; Chow, J.C.; Rombeau, J.L.; Smith, R.J. Gut adaptation and the insulin-like growth factor system: Regulation by glutamine and IGF-I administration. Am. J. Physiol. 1996, 271, G866–G875. [Google Scholar] [PubMed]
- Blikslager, A.T.; Rhoads, J.M.; Bristol, D.G.; Roberts, M.C.; Argenzio, R.A. Glutamine and transforming growth factor-α stimulate extracellular regulated kinases and enhance recovery of villous surface area in porcine ischemic-injured intestine. Surgery 1999, 125, 186–194. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, L.; Betanzos, A.; Nava, P.; Jaramillo, B. Tight junction proteins. Prog. Biophys. Mol. Biol. 2003, 81, 1–44. [Google Scholar] [CrossRef]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Mitic, L.L.; Anderson, J.M. Molecular architecture of tight junctions. Annu. Rev. Physiol. 1998, 60, 121–142. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Van Itallie, C.M. Tight junctions and the molecular basis for regulation of paracellular permeability. Am. J. Physiol. 1995, 269, G467–G475. [Google Scholar] [PubMed]
- Harhaj, N.S.; Antonetti, D.A. Regulation of tight junctions and loss of barrier function in pathophysiology. Int. J. Biochem. Cell Biol. 2004, 36, 1206–1237. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, J.; Kojima, T.; Ogasawara, N.; Kamekura, R.; Kurose, M.; Go, M.; Harimaya, A.; Murata, M.; Osanai, M.; Chiba, H.; et al. Protein kinase C enhances tight junction barrier function of human nasal epithelial cells in primary culture by transcriptional regulation. Mol. Pharmacol. 2008, 74, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Basuroy, S.; Seth, A.; Elias, B.; Naren, A.P.; Rao, R. MAPK interacts with occludin and mediates EGF-induced prevention of tight junction disruption by hydrogen peroxide. Biochem. J. 2006, 393 Pt 1, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R.; Rill, B.K.; Carlson, S.L.; Carnes, D.; Kerner, R.; Mrsny, R.J.; Madara, J.L. Physiological regulation of epithelial tight junctions is associated with myosin light-chain phosphorylation. Am. J. Physiol.Cell Physiol. 1997, 273, C1378–C1385. [Google Scholar]
- Rigor, R.R.; Shen, Q.; Pivetti, C.D.; Wu, M.H.; Yuan, S.Y. Myosin light chain kinase signaling in endothelial barrier dysfunction. Med. Res. Rev. 2013, 33, 911–933. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Lewis, P.; Samuelson, D.; Liboni, K.; Neu, J. Glutamine regulates Caco-2 cell tight junction proteins. Am. J. Physiol. Gastrointest. Liv. Physiol. 2004, 287, G726–G733. [Google Scholar] [CrossRef] [PubMed]
- DeMarco, V.G.; Li, N.; Thomas, J.; West, C.M.; Neu, J. Glutamine and barrier function in cultured Caco-2 epithelial cell monolayers. J. Nutr. 2003, 133, 2176–2179. [Google Scholar] [PubMed]
- Beutheu, S.; Ghouzali, I.; Galas, L.; Dechelotte, P.; Coeffier, M. Glutamine and arginine improve permeability and tight junction protein expression in methotrexate-treated Caco-2 cells. Clin. Nutr. 2013, 32, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.; Basuroy, S.; Sheth, P.; Rao, R.K. l-Glutamine ameliorates acetaldehyde-induced increase in paracellular permeability in Caco-2 cell monolayer. Am. J. Physiol. Gastrointest. Liv. Physiol. 2004, 287, G510–G517. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mariscal, L.; Tapia, R.; Chamorro, D. Crosstalk of tight junction components with signaling pathways. Biochim. Biophys. Acta 2008, 1778, 729–756. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Neu, J. Glutamine deprivation alters intestinal tight junctions via a PI3-K/Akt mediated pathway in Caco-2 cells. J. Nutr. 2009, 139, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, A.; Furuse, M.; Saitou, M.; Ando-Akatsuka, Y.; Tsukita, S. Possible involvement of phosphorylation of occludin in tight junction formation. J. Cell Biol. 1997, 137, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, R.; Dyukova, E.; Singh, N.K.; Ohba, M.; Mobley, J.A.; Rao, G.N. Vascular endothelial tight junctions and barrier function are disrupted by 15 S-hydroxyeicosatetraenoic acid partly via protein kinase Cϵ-mediated zona occludens-1 phosphorylation at threonine 770/772. J. Biol. Chem. 2014, 289, 3148–3163. [Google Scholar] [CrossRef] [PubMed]
- Dorfel, M.J.; Westphal, J.K.; Bellmann, C.; Krug, S.M.; Cording, J.; Mittag, S.; Tauber, R.; Fromm, M.; Blasig, I.E.; Huber, O. CK2-dependent phosphorylation of occludin regulates the interaction with ZO-proteins and tight junction integrity. Cell Commun. Signal. CCS 2013, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wu, Z.; Ji, Y.; Sun, K.; Dai, Z.; Wu, G. l-Glutamine Enhances Tight Junction Integrity by Activating CaMK Kinase 2–AMP-Activated Protein Kinase Signaling in Intestinal Porcine Epithelial Cells. J. Nutr. 2016, 146, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Slavov, D.; Wischmeyer, P.E. Glutamine-mediated dual regulation of heat shock transcription factor-1 activation and expression. J. Biol. Chem. 2012, 287, 40400–40413. [Google Scholar] [CrossRef] [PubMed]
- Ullman, T.A.; Itzkowitz, S.H. Intestinal inflammation and cancer. Gastroenterology 2011, 140, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, J.M.; Wu, G. Glutamine, arginine, and leucine signaling in the intestine. Amino Acids 2009, 37, 111–122. [Google Scholar]
- Baltimore, D. NF-κB is 25. Nat. Immunol. 2011, 12, 683. [Google Scholar] [CrossRef] [PubMed]
- Ganeshan, K.; Chawla, A. Metabolic regulation of immune responses. Annu. Rev. Immunol. 2014, 32, 609–634. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.C.; Chu, C.C.; Ko, T.L.; Yeh, C.L.; Yeh, S.L. Effects of alanyl-glutamine dipeptide on the expression of colon-inflammatory mediators during the recovery phase of colitis induced by dextran sulfate sodium. Eur. J. Nutr. 2013, 52, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Sufit, A.J.; Wischmeyer, P.E. Glutamine therapy improves outcome of in vitro and in vivo experimental colitis models. J. Parenter. Enter. Nutr. 2011, 35, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Haynes, T.E.; Li, P.; Li, X.; Shimotori, K.; Sato, H.; Flynn, N.E.; Wang, J.; Knabe, D.A.; Wu, G. l-Glutamine or l-alanyl-l-glutamine prevents oxidant- or endotoxin-induced death of neonatal enterocytes. Amino Acids 2009, 37, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, V.; Eaves-Pyles, T.; Odoms, K.; Quaid, G.; Shanley, T.P.; Wong, H.R. Heat shock inhibits activation of NF-κB in the absence of heat shock factor-1. Biochem. Biophys. Res. Commun. 2002, 291, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Liboni, K.C.; Li, N.; Scumpia, P.O.; Neu, J. Glutamine modulates LPS-induced IL-8 production through IkappaB/NF-κB in human fetal and adult intestinal epithelium. J. Nutr. 2005, 135, 245–251. [Google Scholar] [PubMed]
- Hubert-Buron, A.; Leblond, J.; Jacquot, A.; Ducrotte, P.; Dechelotte, P.; Coeffier, M. Glutamine pretreatment reduces IL-8 production in human intestinal epithelial cells by limiting IκBα ubiquitination. J. Nutr. 2006, 136, 1461–1465. [Google Scholar] [PubMed]
- Kretzmann, N.A.; Fillmann, H.; Mauriz, J.L.; Marroni, C.A.; Marroni, N.; Gonzalez-Gallego, J.; Tunon, M.J. Effects of glutamine on proinflammatory gene expression and activation of nuclear factor κB and signal transducers and activators of transcription in TNBS-induced colitis. Inflamm. Bowel Dis. 2008, 14, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.H. STAT signaling in inflammation. Jak-Stat 2013, 2, e24198. [Google Scholar] [CrossRef] [PubMed]
- Ihle, J.N. The Stat family in cytokine signaling. Curr. Opin. Cell Biol. 2001, 13, 211–217. [Google Scholar] [CrossRef]
- Liboni, K.; Li, N.; Neu, J. Mechanism of glutamine-mediated amelioration of lipopolysaccharide-induced IL-8 production in Caco-2 cells. Cytokine 2004, 26, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Kubes, P.; McCafferty, D.M. Nitric oxide and intestinal inflammation. J. Am. Med. 2000, 109, 150–158. [Google Scholar] [CrossRef]
- Hecker, M.; Sessa, W.C.; Harris, H.J.; Anggard, E.E.; Vane, J.R. The metabolism of l-arginine and its significance for the biosynthesis of endothelium-derived relaxing factor: Cultured endothelial cells recycle l-citrulline to l-arginine. Proc. Natl. Acad. Sci. USA 1990, 87, 8612–8616. [Google Scholar] [CrossRef] [PubMed]
- Swierkosz, T.A.; Mitchell, J.A.; Sessa, W.C.; Hecker, M.; Vane, J.R. l-Glutamine inhibits the release of endothelium-derived relaxing factor from the rabbit aorta. Biochem. Biophys. Res. Commun. 1990, 172, 143–148. [Google Scholar] [CrossRef]
- Houdijk, A.P.; Visser, J.J.; Rijnsburger, E.R.; Teerlink, T.; van Leeuwen, P.A. Dietary glutamine supplementation reduces plasma nitrate levels in rats. Clin. Nutr. 1998, 17, 11–14. [Google Scholar] [CrossRef]
- Suh, G.J.; Youn, Y.K.; Song, H.G.; Rhee, J.E.; Jung, S.E. The effect of glutamine on inducible nitric oxide synthase gene expression in intestinal ischemia-reperfusion injury. Nutr. Res. 2003, 23, 131–140. [Google Scholar]
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Mates, J.M.; Perez-Gomez, C.; Nunez de Castro, I.; Asenjo, M.; Marquez, J. Glutamine and its relationship with intracellular redox status, oxidative stress and cell proliferation/death. Int. J. Biochem. Cell Biol. 2002, 34, 439–458. [Google Scholar] [CrossRef]
- Watson, A.J.; Pritchard, D.M., VII. Apoptosis in intestinal epithelium: Lessons from transgenic and knockout mice. Am. J. Physiol. Gastrointest. Liv. Physiol. 2000, 278, G1–G5. [Google Scholar]
- Que, F.G.; Gores, G.J. Cell death by apoptosis: Basic concepts and disease relevance for the gastroenterologist. Gastroenterology 1996, 110, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Demehri, F.R.; Barrett, M.; Ralls, M.W.; Miyasaka, E.A.; Feng, Y.; Teitelbaum, D.H. Intestinal epithelial cell apoptosis and loss of barrier function in the setting of altered microbiota with enteral nutrient deprivation. Front. Cell. Infect. Microbiol. 2013, 3, 105. [Google Scholar] [CrossRef] [PubMed]
- Zatorski, H.; Marynowski, M.; Fichna, J. Is insulin-like growth factor 1 (IGF-1) system an attractive target inflammatory bowel diseases? Benefits and limitation of potential therapy. Pharmacol. Rep. 2016, 68, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Sanchez de Medina, F.; Romero-Calvo, I.; Mascaraque, C.; Martinez-Augustin, O. Intestinal inflammation and mucosal barrier function. Inflamm. Bowel Dis. 2014, 20, 2394–2404. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Koji, T.; Makiyama, K.; Kobayashi, N.; Nakane, P.K. Apoptosis of crypt epithelial cells in ulcerative colitis. J. Pathol. 1996, 180, 152–159. [Google Scholar] [CrossRef]
- Kim, J.M.; Eckmann, L.; Savidge, T.C.; Lowe, D.C.; Witthoft, T.; Kagnoff, M.F. Apoptosis of human intestinal epithelial cells after bacterial invasion. J. Clin. Investig. 1998, 102, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- Schumann, M.; Siegmund, B.; Schulzke, J.D.; Fromm, M. Celiac Disease: Role of the Epithelial Barrier. Cellular Mol. Gastroenterol. Hepatol. 2017, 3, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Hyoh, Y.; Ishizaka, S.; Horii, T.; Fujiwara, A.; Tegoshi, T.; Yamada, M.; Arizono, N. Activation of caspases in intestinal villus epithelial cells of normal and nematode infected rats. Gut 2002, 50, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Madesh, M.; Balasubramanian, K.A. Apoptosis in the intestinal epithelium: Its relevance in normal and pathophysiological conditions. J. Gastroenterol. Hepatol. 2000, 15, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Papaconstantinou, H.T.; Hwang, K.O.; Rajaraman, S.; Hellmich, M.R.; Townsend, C.M., Jr.; Ko, T.C. Glutamine deprivation induces apoptosis in intestinal epithelial cells. Surgery 1998, 124, 152–159, discussion 159–160. [Google Scholar] [CrossRef]
- Carneiro, B.A.; Fujii, J.; Brito, G.A.; Alcantara, C.; Oria, R.B.; Lima, A.A.; Obrig, T.; Guerrant, R.L. Caspase and bid involvement in Clostridium difficile toxin A-induced apoptosis and modulation of toxin A effects by glutamine and alanyl-glutamine in vivo and in vitro. Infect. Immun. 2006, 74, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Takayama, C.; Mukaizawa, F.; Fujita, T.; Ogawara, K.; Higaki, K.; Kimura, T. Amino acids suppress apoptosis induced by sodium laurate, an absorption enhancer. J. Pharm. Sci. 2009, 98, 4629–4638. [Google Scholar] [CrossRef] [PubMed]
- Roth, E.; Oehler, R.; Manhart, N.; Exner, R.; Wessner, B.; Strasser, E.; Spittler, A. Regulative potential of glutamine—Relation to glutathione metabolism. Nutrition 2002, 18, 217–221. [Google Scholar] [CrossRef]
- Hall, A. The role of glutathione in the regulation of apoptosis. Eur. J. Clin. Investig. 1999, 29, 238–245. [Google Scholar] [CrossRef]
- Fan, T.J.; Han, L.H.; Cong, R.S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Larson, S.D.; Li, J.; Chung, D.H.; Evers, B.M. Molecular mechanisms contributing to glutamine-mediated intestinal cell survival. Am. J. Physiol. Gastrointest. Liv. Physiol. 2007, 293, G1262–G1271. [Google Scholar] [CrossRef] [PubMed]
- Wischmeyer, P.E. Glutamine and heat shock protein expression. Nutrition 2002, 18, 225–228. [Google Scholar] [CrossRef]
- Garrido, C.; Gurbuxani, S.; Ravagnan, L.; Kroemer, G. Heat shock proteins: Endogenous modulators of apoptotic cell death. Biochem. Biophys. Res. Commun. 2001, 286, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Singleton, K.D.; Serkova, N.; Beckey, V.E.; Wischmeyer, P.E. Glutamine attenuates lung injury and improves survival after sepsis: Role of enhanced heat shock protein expression. Crit. Care Med. 2005, 33, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Ropeleski, M.J.; Riehm, J.; Baer, K.A.; Musch, M.W.; Chang, E.B. Anti-apoptotic effects of l-glutamine-mediated transcriptional modulation of the heat shock protein 72 during heat shock. Gastroenterology 2005, 129, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A.; Lee, A.-H.; Franke, A.; Glickman, J.N.; Zeissig, S.; Tilg, H.; Nieuwenhuis, E.E.; Higgins, D.E.; Schreiber, S.; Glimcher, L.H. XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease. Cell 2008, 134, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Crespo, I.; San-Miguel, B.; Prause, C.; Marroni, N.; Cuevas, M.J.; Gonzalez-Gallego, J.; Tunon, M.J. Glutamine treatment attenuates endoplasmic reticulum stress and apoptosis in TNBS-induced colitis. PLoS ONE 2012, 7, e50407. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Hampe, J.; Franke, A.; Rosenstiel, P.; Till, A.; Teuber, M.; Huse, K.; Albrecht, M.; Mayr, G.; de La Vega, F.M.; Briggs, J.; et al. A genome-wide association scan of nonsynonymous SNPs identifies a susceptibility variant for Crohn disease in ATG16L1. Nat. Genet. 2007, 39, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Rioux, J.D.; Xavier, R.J.; Taylor, K.D.; Silverberg, M.S.; Goyette, P.; Huett, A.; Green, T.; Kuballa, P.; Barmada, M.M.; Datta, L.W.; et al. Genome-wide association study identifies new susceptibility loci for Crohn disease and implicates autophagy in disease pathogenesis. Nat. Genet. 2007, 39, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Fujita, N.; Jang, M.H.; Uematsu, S.; Yang, B.G.; Satoh, T.; Omori, H.; Noda, T.; Yamamoto, N.; Komatsu, M.; et al. Loss of the autophagy protein Atg16L1 enhances endotoxin-induced IL-1beta production. Nature 2008, 456, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Fujishima, Y.; Nishiumi, S.; Masuda, A.; Inoue, J.; Nguyen, N.M.; Irino, Y.; Komatsu, M.; Tanaka, K.; Kutsumi, H.; Azuma, T.; et al. Autophagy in the intestinal epithelium reduces endotoxin-induced inflammatory responses by inhibiting NF-kappaB activation. Arch. Biochem. Biophys. 2011, 506, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Cadwell, K.; Patel, K.K.; Komatsu, M.; Virgin, H.W.T.; Stappenbeck, T.S. A common role for Atg16L1, Atg5 and Atg7 in small intestinal Paneth cells and Crohn disease. Autophagy 2009, 5, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.K.; Stappenbeck, T.S. Autophagy and intestinal homeostasis. Annu. Rev. Physiol. 2013, 75, 241–262. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama, T.; Musch, M.W.; Ropeleski, M.J.; Tsubouchi, H.; Chang, E.B. Glutamine increases autophagy under Basal and stressed conditions in intestinal epithelial cells. Gastroenterology 2009, 136, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Ali, S.M.; Sabatini, D.M. Growing roles for the mTOR pathway. Curr. Opin. Cell Biol. 2005, 17, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Van der Vos, K.E.; Eliasson, P.; Proikas-Cezanne, T.; Vervoort, S.J.; van Boxtel, R.; Putker, M.; van Zutphen, I.J.; Mauthe, M.; Zellmer, S.; Pals, C.; et al. Modulation of glutamine metabolism by the PI(3)K-PKB-FOXO network regulates autophagy. Nat. Cell Biol. 2012, 14, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Sido, B.; Seel, C.; Hochlehnert, A.; Breitkreutz, R.; Droge, W. Low intestinal glutamine level and low glutaminase activity in Crohn’s disease: A rational for glutamine supplementation? Dig. Dis. Sci. 2006, 51, 2170–2179. [Google Scholar] [CrossRef] [PubMed]
- Hsiung, Y.C.; Liu, J.J.; Hou, Y.C.; Yeh, C.L.; Yeh, S.L. Effects of dietary glutamine on the homeostasis of CD4+ T cells in mice with dextran sulfate sodium-induced acute colitis. PLoS ONE 2014, 9, e84410. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.-H.; Liu, J.-J.; Yeh, S.-L.; Chen, W.-J.; Yeh, C.-L. Glutamine modulates acute dextran sulphate sodium-induced changes in small-intestinal intraepithelial γδ-T-lymphocyte expression in mice. Br. J. Nutr. 2014, 111, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Ameho, C.K.; Adjei, A.A.; Harrison, E.K.; Takeshita, K.; Morioka, T.; Arakaki, Y.; Ito, E.; Suzuki, I.; Kulkarni, A.D.; Kawajiri, A.; et al. Prophylactic effect of dietary glutamine supplementation on interleukin 8 and tumour necrosis factor α production in trinitrobenzene sulphonic acid induced colitis. Gut 1997, 41, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Souba, W.W.; Klimberg, V.S.; Hautamaki, R.D.; Mendenhall, W.H.; Bova, F.C.; Howard, R.J.; Bland, K.I.; Copeland, E.M. Oral glutamine reduces bacterial translocation following abdominal radiation. J. Surg. Res. 1990, 48, 1–5. [Google Scholar] [CrossRef]
- Lee, W.Y.; Hu, Y.M.; Ko, T.L.; Yeh, S.L.; Yeh, C.L. Glutamine modulates sepsis-induced changes to intestinal intraepithelial gammadeltaT lymphocyte expression in mice. Shock 2012, 38, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Tung, J.N.; Lee, W.Y.; Pai, M.H.; Chen, W.J.; Yeh, C.L.; Yeh, S.L. Glutamine modulates CD8αα(+) TCRαβ(+) intestinal intraepithelial lymphocyte expression in mice with polymicrobial sepsis. Nutrition 2013, 29, 911–917. [Google Scholar] [CrossRef] [PubMed]
- García-de-Lorenzo, A.; Zarazaga, A.; García-Luna, P.P.; Gonzalez-Huix, F.; López-Martínez, J.; Miján, A.; Quecedo, L.; Casimiro, C.; Usán, L.; del Llano, J. Clinical evidence for enteral nutritional support with glutamine: A systematic review. Nutrition 2003, 19, 805–811. [Google Scholar] [CrossRef]
- Benjamin, J.; Makharia, G.; Ahuja, V.; Rajan, K.A.; Kalaivani, M.; Gupta, S.D.; Joshi, Y.K. Glutamine and whey protein improve intestinal permeability and morphology in patients with Crohn’s disease: A randomized controlled trial. Dig. Dis. Sci. 2012, 57, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.J.; Avenell, A.; Noble, D.W.; Campbell, M.K.; Croal, B.L.; Simpson, W.G.; Vale, L.D.; Battison, C.G.; Jenkinson, D.J.; Cook, J.A. Randomised trial of glutamine, selenium, or both, to supplement parenteral nutrition for critically ill patients. BMJ 2011, 342, d1542. [Google Scholar] [CrossRef] [PubMed]
- Heyland, D.; Muscedere, J.; Wischmeyer, P.E.; Cook, D.; Jones, G.; Albert, M.; Elke, G.; Berger, M.M.; Day, A.G.; Canadian Critical Care Trials Group. A randomized trial of glutamine and antioxidants in critically ill patients. N. Engl. J. Med. 2013, 368, 1489–1497. [Google Scholar] [CrossRef] [PubMed]
- Akobeng, A.K.; Miller, V.; Stanton, J.; Elbadri, A.M.; Thomas, A.G. Double-blind randomized controlled trial of glutamine-enriched polymeric diet in the treatment of active Crohn’s disease. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Den Hond, E.; Hiele, M.; Peeters, M.; Ghoos, Y.; Rutgeerts, P. Effect of long-term oral glutamine supplements on small intestinal permeability in patients with Crohn’s disease. J. Parenter. Enter. Nutr. 1999, 23, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Scolapio, J.S.; Camilleri, M.; Fleming, C.R.; Oenning, L.; Burton, D.; Sebo, T.J.; Batts, K.; Kelly, D. Effect of growth hormone, glutamine, and diet on adaptation in short-bowel syndrome: A randomized, controlled study. Gastroenterology 1997, 113, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Beaugerie, L.; Carbonnel, F.; Hecketsweiler, B.; Dechelotte, P.; Gender, J.-P.; Cosnes, J. Effects of an isotonic oral rehydration solution, enriched with glutamine, on fluid and sodium absorption in patients with short-bowel syndrome. Aliment. Pharmacol. Ther. 1997, 11, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Scolapio, J.S. Effect of growth hormone, glutamine, and diet on body composition in short bowel syndrome: A randomized, controlled study. J. Parenter. Enter. Nutr. 1999, 23, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Szkudlarek, J.; Jeppesen, P.; Mortensen, P. Effect of high dose growth hormone with glutamine and no change in diet on intestinal absorption in short bowel patients: A randomised, double blind, crossover, placebo controlled study. Gut 2000, 47, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, P.; Szkudlarek, J.; Høy, C.-E.; Mortensen, P. Effect of high-dose growth hormone and glutamine on body composition, urine creatinine excretion, fatty acid absorption, and essential fatty acids status in short bowel patients. A randomized, double-blind, crossover, placebo-controlled study. Scand. J. Gastroenterol. 2001, 36, 48–54. [Google Scholar] [PubMed]
- Scolapio, J.; McGreevy, K.; Tennyson, G.; Burnett, O. Effect of glutamine in short-bowel syndrome. Clin. Nutr. 2001, 20, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Alpers, D.H. Glutamine: Do the data support the cause for glutamine supplementation in humans? Gastroenterology 2006, 130, S106–S116. [Google Scholar] [CrossRef] [PubMed]
- Gore, D.C.; Wolfe, R.R. Metabolic response of muscle to alanine, glutamine, and valine supplementation during severe illness. J. Parenter. Enter. Nutr. 2003, 27, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Oudemans-van Straaten, H.M.; Bosman, R.J.; Treskes, M.; van der Spoel, H.J.; Zandstra, D.F. Plasma glutamine depletion and patient outcome in acute ICU admissions. Intensiv. Care Med. 2001, 27, 84–90. [Google Scholar] [CrossRef]
- MacFie, J. Enteral versus parenteral nutrition. Br. J. Surg. 2000, 87, 1121–1122. [Google Scholar] [CrossRef] [PubMed]
- Boelens, P.G.; Melis, G.C.; van Leeuwen, P.A.; ten Have, G.A.; Deutz, N.E. Route of administration (enteral or parenteral) affects the contribution of l-glutamine to de novo L-arginine synthesis in mice: A stable-isotope study. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E683–E690. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Huix, F.; Fernandez-Banares, F.; Esteve-Comas, M.; Abad-Lacruz, A.; Cabre, E.; Acero, D.; Figa, M.; Guilera, M.; Humbert, P.; de León, R. Enteral versus parenteral nutrition as adjunct therapy in acute ulcerative colitis. Am. J. Gastroenterol. 1993, 88, 2. [Google Scholar]
- Buchman, A.L.; Moukarzel, A.A.; Bhuta, S.; Belle, M.; Ament, M.E.; Eckhert, C.D.; Hollander, D.; Gornbeln, J.; Kopple, J.D.; Vijayaroghavan, S.R. Parenteral nutrition is associated with intestinal morphologic and functional changes in humans. J. Parenter. Enter. Nutr. 1995, 19, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Noyer, C.M.; Simon, D.; Borczuk, A.; Brandt, L.J.; Lee, M.J.; Nehra, V. A double-blind placebo-controlled pilot study of glutamine therapy for abnormal intestinal permeability in patients with AIDS. Am. J. Gastroenterol. 1998, 93, 972–975. [Google Scholar] [CrossRef] [PubMed]
- Goeters, C.; Wenn, A.; Mertes, N.; Wempe, C.; Van Aken, H.; Stehle, P.; Bone, H.-G. Parenteral l-alanyl-l-glutamine improves 6-month outcome in critically ill patients. Crit. Care Med. 2002, 30, 2032–2037. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.D.; Jones, C.; Palmer, T.A. Six-month outcome of critically ill patients given glutamine-supplemented parenteral nutrition. Nutrition 1997, 13, 295–302. [Google Scholar] [CrossRef]
- Griffiths, R.D.; Allen, K.D.; Andrews, F.J.; Jones, C. Infection, multiple organ failure, and survival in the intensive care unit: Influence of glutamine-supplemented parenteral nutrition on acquired infection. Nutrition 2002, 18, 546–552. [Google Scholar] [CrossRef]
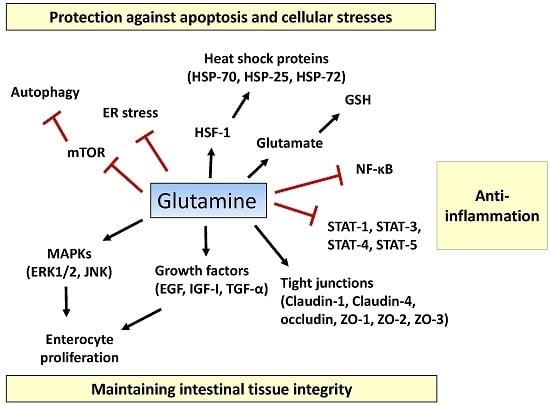
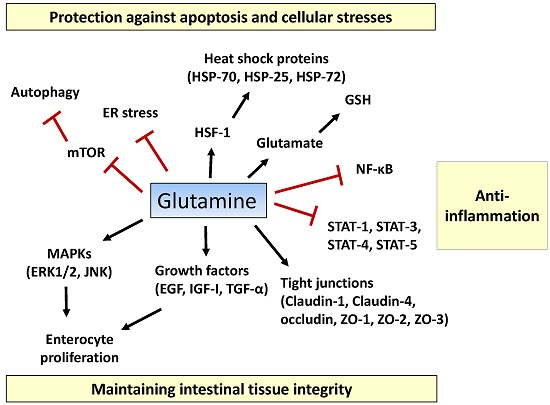
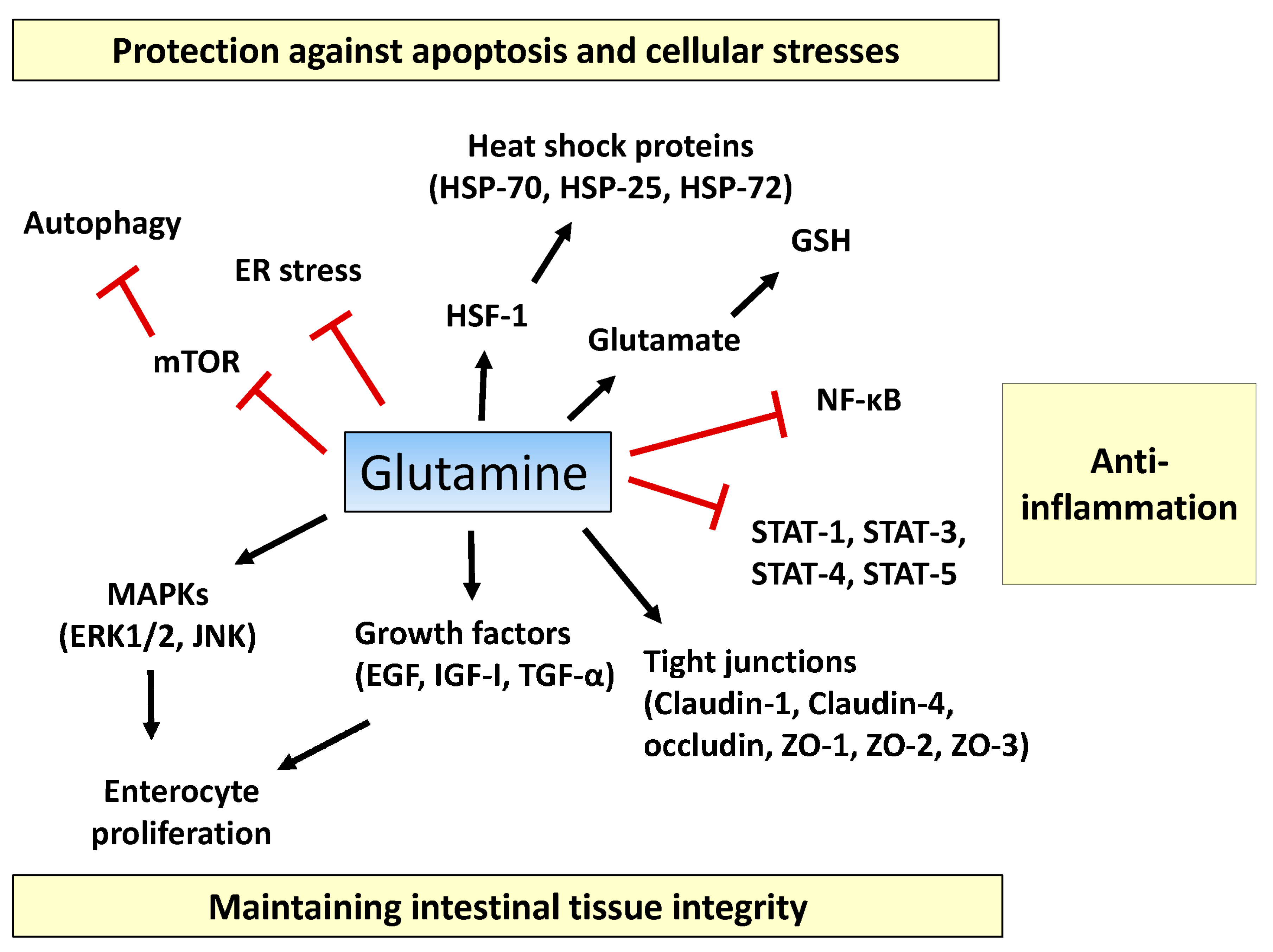
Figure 1.
Proposed action mechanisms of glutamine in intestinal cells. Glutamine maintains intestinal tissue integrity via promoting enterocyte proliferation, activation of mitogen-activated protein kinases (MAPKs) (ERK1/2, JNK1/2), optimizing the actions of growth factors (epidermal growth factor (EGF), insulin-like growth factor (IGF)-I, transforming growth factor (TGF)-α), and inducing expression of tight-junction proteins (claudin-1, claudin-4, occludin, zonula occludens (ZO)-1, ZO-2, and ZO-3). Pro-inflammatory signaling pathways such as the nuclear factor-κB (NF-κB) and signal transducers and activators of transcription (STAT) pathways are inhibited by glutamine. Glutamine suppresses extensive apoptosis by participating in the synthesis of glutathione (GSH) and by regulating heat shock factor (HSF)-1-mediated expression of heat shock proteins (HSPs). Glutamine ameliorates endoplasmic reticulum (ER) stress and promotes autophagy by inhibiting the mechanistic target of rapamycin (mTOR) pathway, thus protecting intestinal cells from stressful conditions. T bars mean inhibition while arrows represent stimulation.
Figure 1.
Proposed action mechanisms of glutamine in intestinal cells. Glutamine maintains intestinal tissue integrity via promoting enterocyte proliferation, activation of mitogen-activated protein kinases (MAPKs) (ERK1/2, JNK1/2), optimizing the actions of growth factors (epidermal growth factor (EGF), insulin-like growth factor (IGF)-I, transforming growth factor (TGF)-α), and inducing expression of tight-junction proteins (claudin-1, claudin-4, occludin, zonula occludens (ZO)-1, ZO-2, and ZO-3). Pro-inflammatory signaling pathways such as the nuclear factor-κB (NF-κB) and signal transducers and activators of transcription (STAT) pathways are inhibited by glutamine. Glutamine suppresses extensive apoptosis by participating in the synthesis of glutathione (GSH) and by regulating heat shock factor (HSF)-1-mediated expression of heat shock proteins (HSPs). Glutamine ameliorates endoplasmic reticulum (ER) stress and promotes autophagy by inhibiting the mechanistic target of rapamycin (mTOR) pathway, thus protecting intestinal cells from stressful conditions. T bars mean inhibition while arrows represent stimulation.

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, M.-H.; Kim, H. The Roles of Glutamine in the Intestine and Its Implication in Intestinal Diseases. Int. J. Mol. Sci. 2017, 18, 1051. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18051051
AMA Style
Kim M-H, Kim H. The Roles of Glutamine in the Intestine and Its Implication in Intestinal Diseases. International Journal of Molecular Sciences. 2017; 18(5):1051. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18051051
Chicago/Turabian StyleKim, Min-Hyun, and Hyeyoung Kim. 2017. "The Roles of Glutamine in the Intestine and Its Implication in Intestinal Diseases" International Journal of Molecular Sciences 18, no. 5: 1051. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18051051
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.