
Stability Profiles and Therapeutic Effect of Cu/Zn Superoxide Dismutase Chemically Coupled to O-Quaternary Chitosan Derivatives against Dextran Sodium Sulfate-Induced Colitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
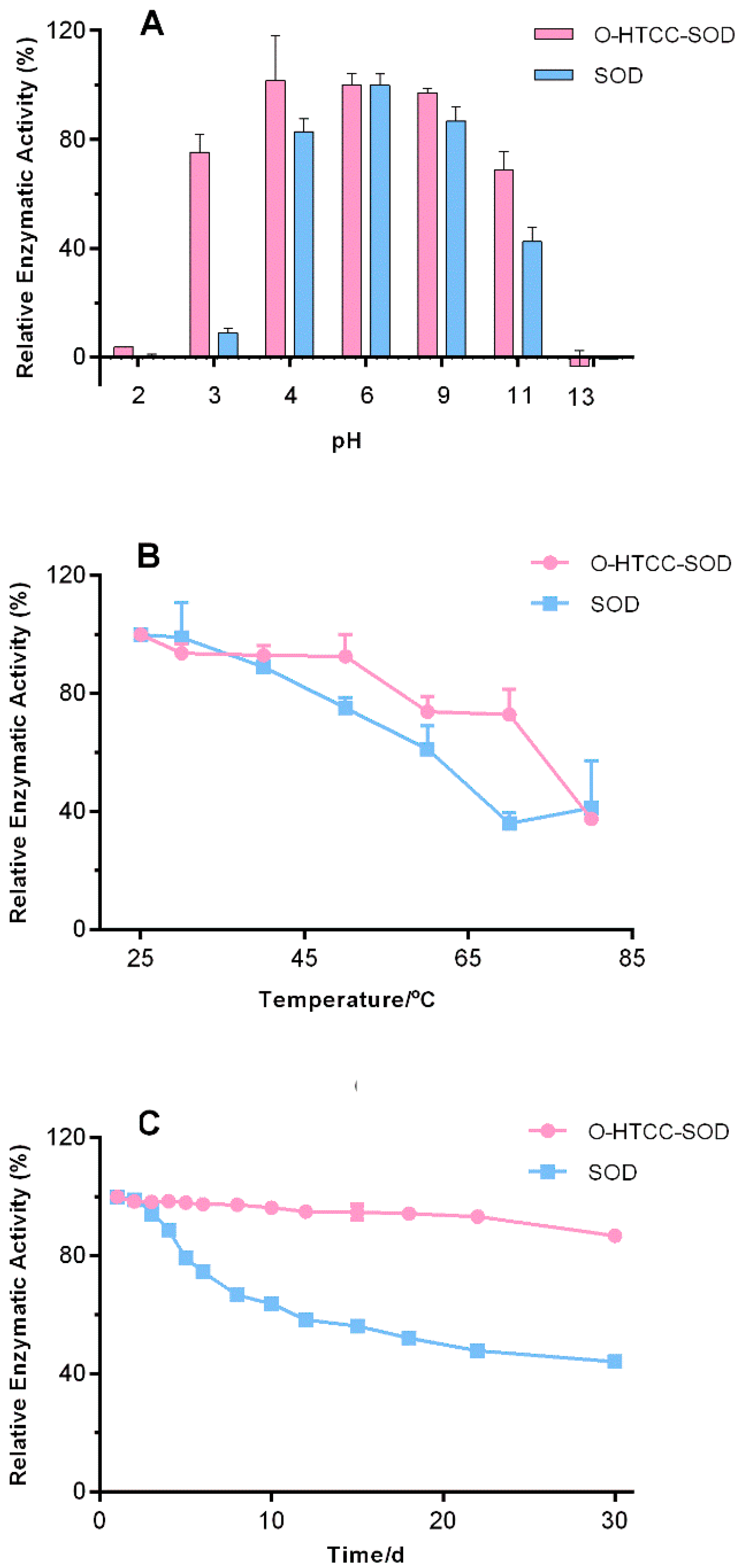
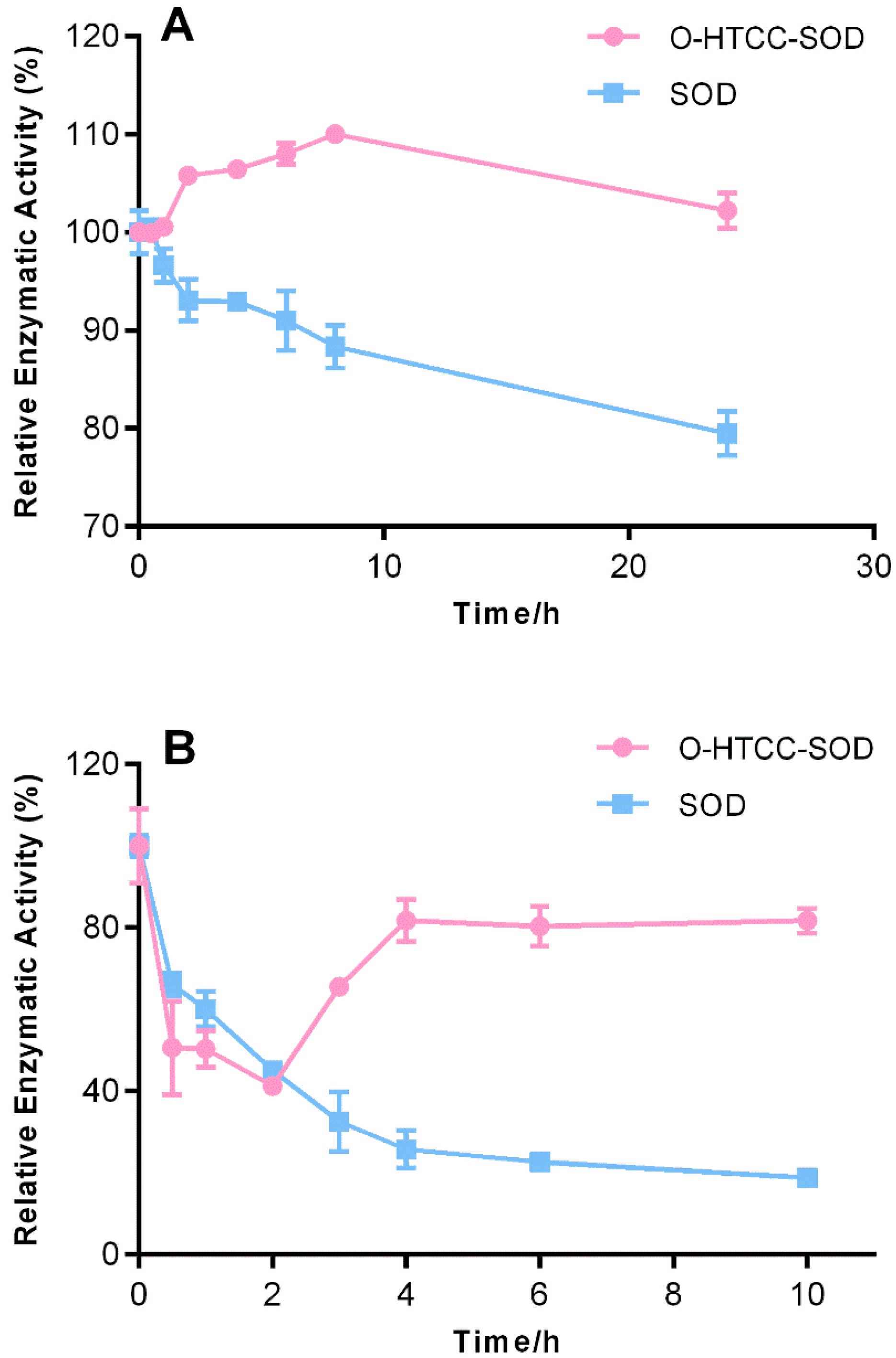
2.1. Stability Profiles of O-Quaternary Chitosan (O-HTCC)-Cu/Zn Superoxide Dismutase (SOD)
2.2. O-HTCC-SOD Reduced Pro-Inflammatory Cytokine Release in Lipopolysaccharides (LPS)-Stimulated Macrophages
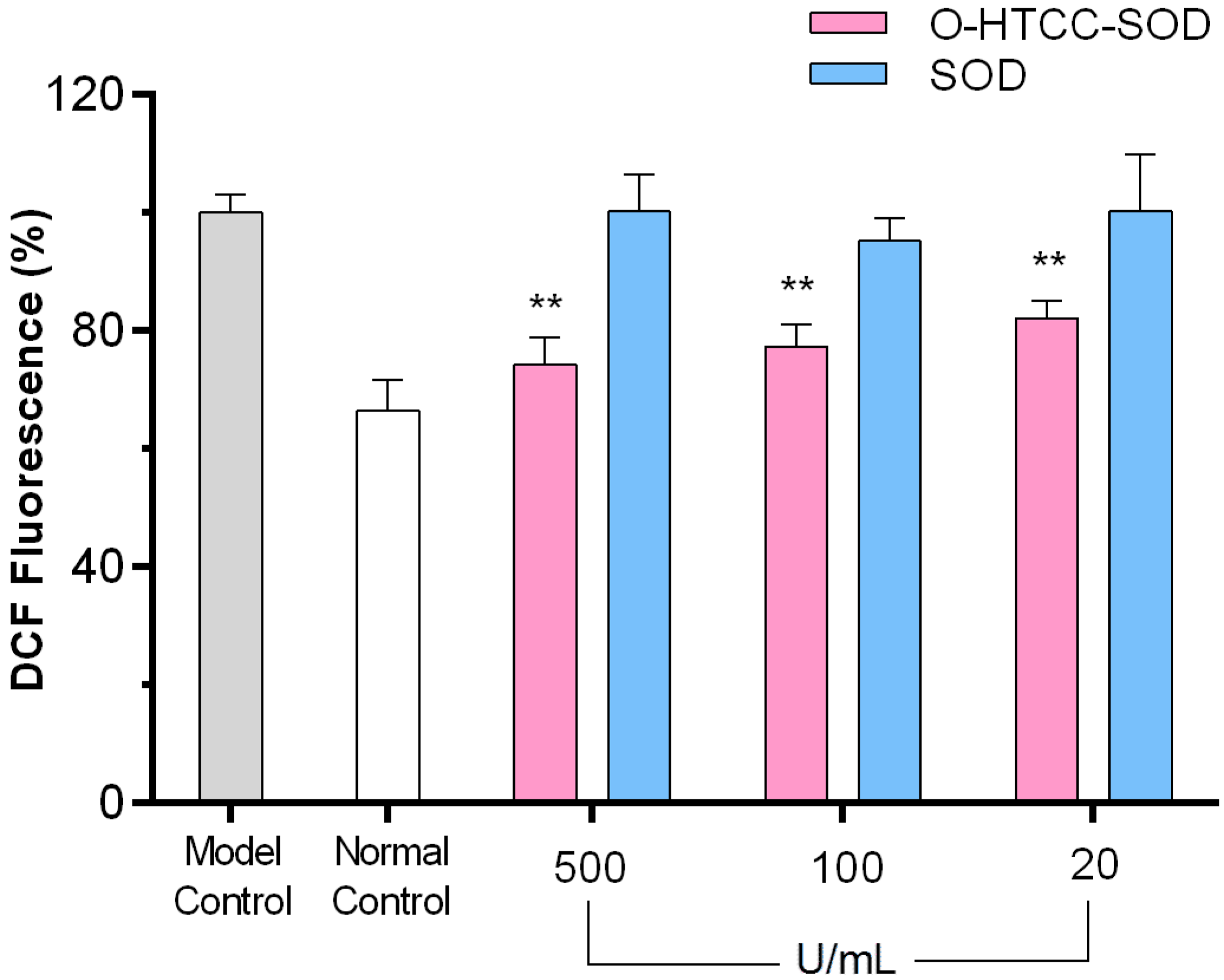
2.3. O-HTCC-SOD Inhibited Intracellular Reactive Oxygen Species (ROS) Production in LPS-Stimulated Macrophages
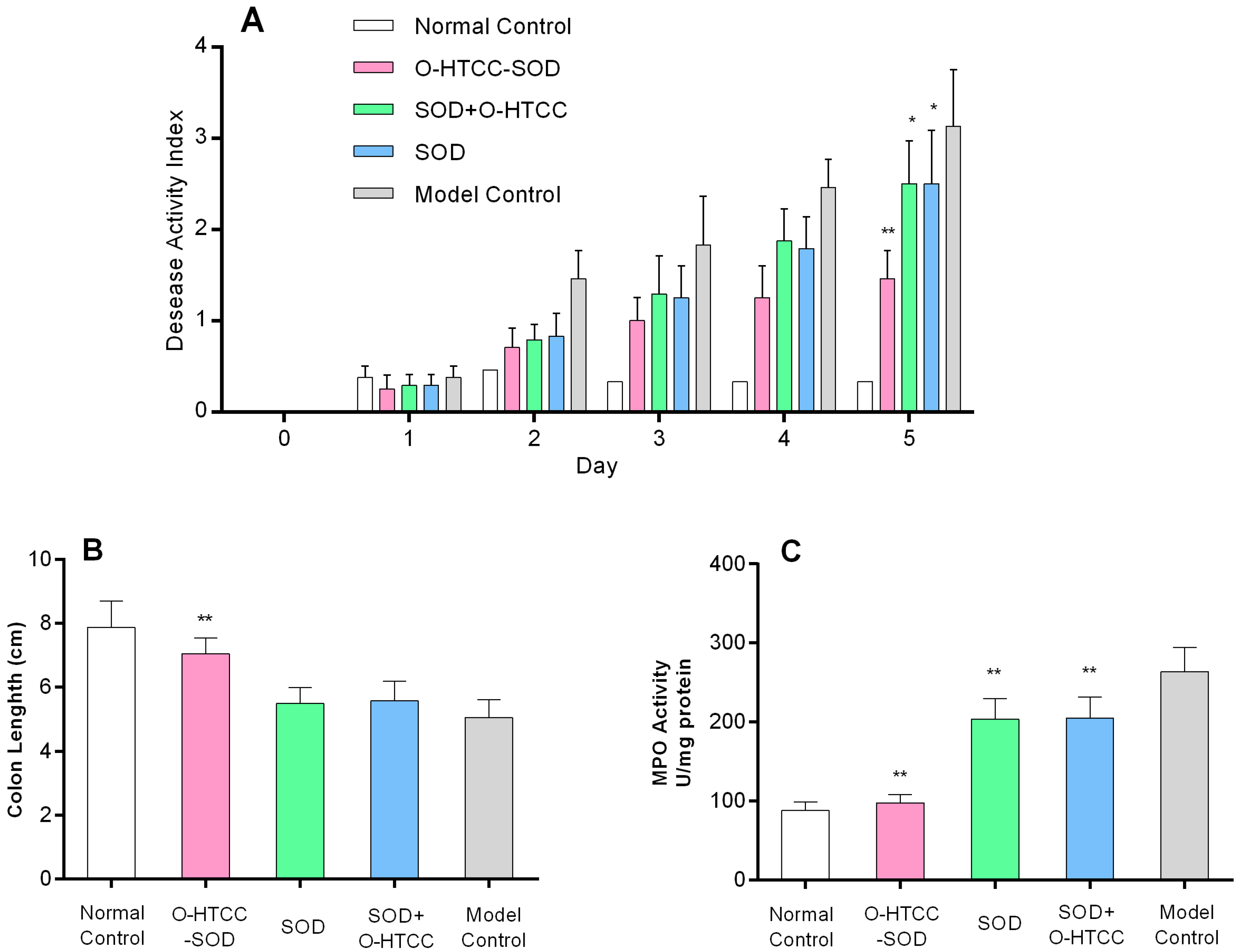
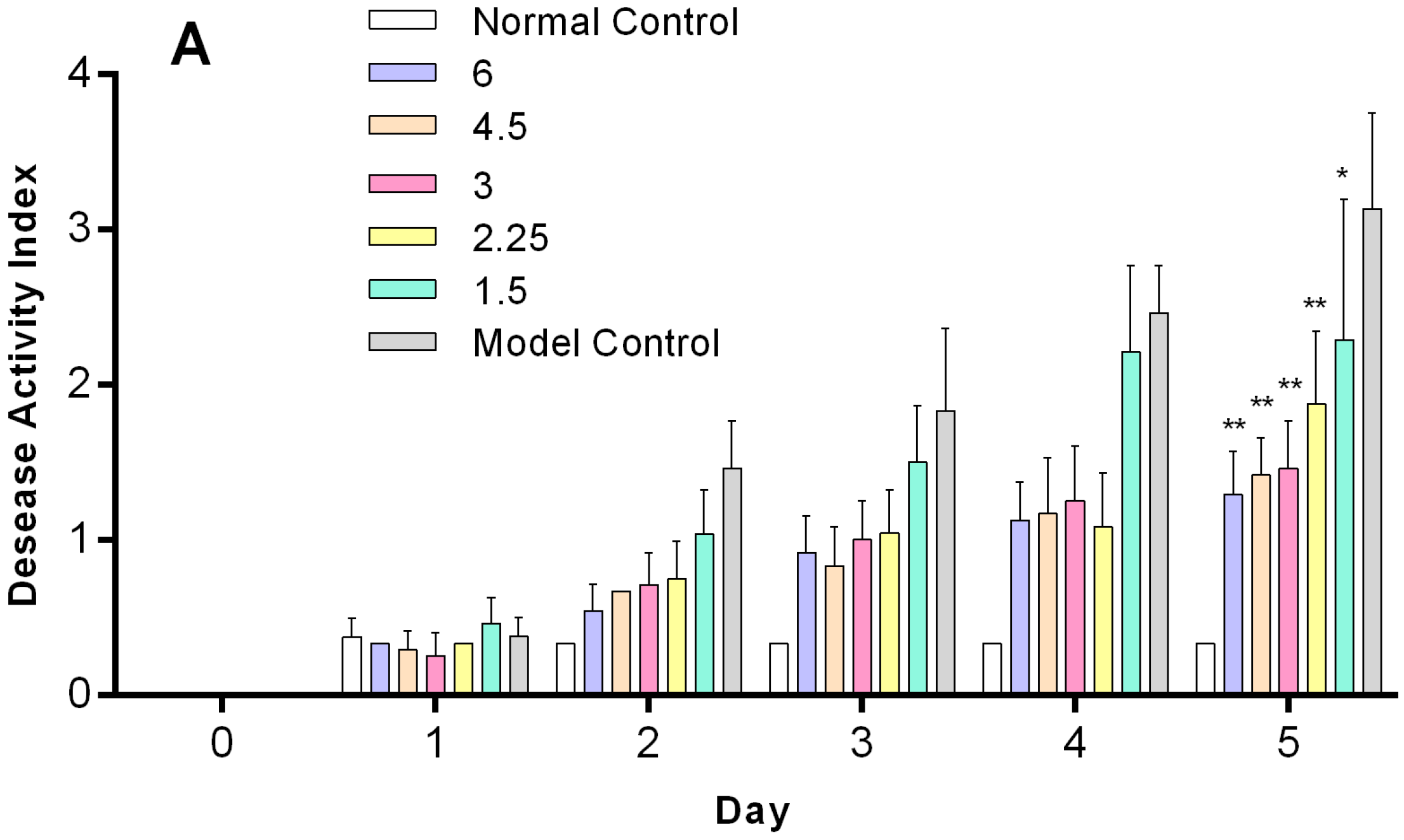
2.4. O-HTCC-SOD Attenuated the Severity of Dextran Sodium Sulfate (DSS)-Induced Colitis
2.5. O-HTCC-SOD Reduced Neutrophil Infiltration into the Colon of DSS-Treated Mice
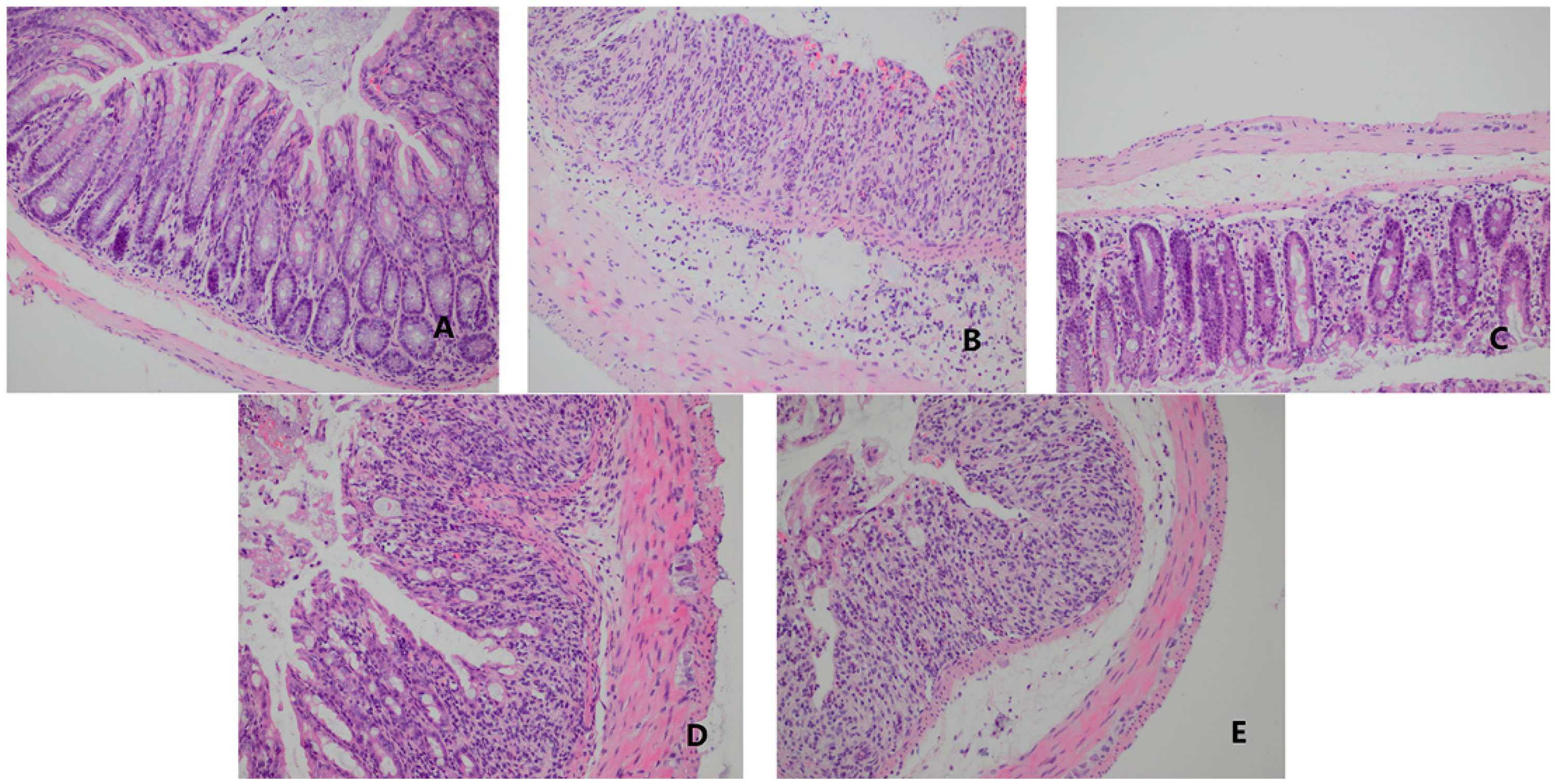
2.6. O-HTCC-SOD Diminished Colonic Histopathological Changes
2.7. Dose–Response Profile of O-HTCC-SOD
3. Materials and Methods
3.1. Materials
3.2. Experimental Animals
3.3. Preparation of O-HTCC-SOD Conjugate
3.4. Enzymatic Activity Assay
3.5. Stability Studies
3.6. Preparation and Viability Assay of Mouse Peritoneal Macrophages
3.7. LPS-Induced Cytokine Production in Macrophages
3.8. LPS-Induced ROS Production in Macrophages
3.9. Models of Experimental Colitis and Treatment
3.10. Evaluation of Colitis Progression
3.11. Detection of MPO Activity in Colon Tissues
3.12. Histopathological Analysis
3.13. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sugamura, K.; Keaney, J.F., Jr. Reactive oxygen species in cardiovascular disease. Free Radic. Biol. Med. 2011, 51, 978–992. [Google Scholar] [CrossRef] [PubMed]
- Schumacker, P.T. Reactive oxygen species in cancer: A dance with the devil. Cancer Cell 2015, 27, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Keating, D.J. Mitochondrial dysfunction, oxidative stress, regulation of exocytosis and their relevance to neurodegenerative diseases. J. Neurochem. 2008, 104, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Gelderman, K.A.; Hultqvist, M.; Olsson, L.M.; Bauer, K.; Pizzolla, A.; Olofsson, P.; Holmdahl, R. Rheumatoid arthritis: The role of reactive oxygen species in disease development and therapeutic strategies. Antioxid. Redox Signal. 2007, 9, 1541–1567. [Google Scholar] [CrossRef] [PubMed]
- Ratnam, D.V.; Ankola, D.D.; Bhardwaj, V.; Sahana, D.K.; Kumar, M.N. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective. J. Control. Release 2006, 113, 189–207. [Google Scholar] [CrossRef] [PubMed]
- De Souza, H.S.; Fiocchi, C. Immunopathogenesis of IBD: Current state of the art. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sun, Y.; Ding, Y.; Wang, X.; Zhou, Y.; Li, W.; Huang, S.; Li, Z.; Kong, L.; Guo, Q.; et al. Gl-v9, a new synthetic flavonoid derivative, ameliorates DSS-induced colitis against oxidative stress by up-regulating TRX-1 expression via activation of AMPK/FOXO3a pathway. Oncotarget 2015, 6, 26291–26307. [Google Scholar] [CrossRef] [PubMed]
- Holmstrom, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. Ros function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.F.; Teixeira, M.; Valentine, J.S. Superoxide dismutases and superoxide reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef] [PubMed]
- Keshavarzian, A.; Morgan, G.; Sedghi, S.; Gordon, J.H.; Doria, M. Role of reactive oxygen metabolites in experimental colitis. Gut 1990, 31, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Deng, C.S.; Chen, D.J.; Zhou, Y.; Xiao, J.Q. Role of copper zinc superoxide dismutase in the short-term treatment of acetic acid-induced colitis in rats. Acta Gastroenterol. Latinoam. 1996, 26, 227–230. [Google Scholar] [PubMed]
- Cuzzocrea, S.; Mazzon, E.; Dugo, L.; Caputi, A.P.; Riley, D.P.; Salvemini, D. Protective effects of M40403, a superoxide dismutase mimetic, in a rodent model of colitis. Eur. J. Pharmacol. 2001, 432, 79–89. [Google Scholar] [CrossRef]
- Manickam, D.S.; Brynskikh, A.M.; Kopanic, J.L.; Sorgen, P.L.; Klyachko, N.L.; Batrakova, E.V.; Bronich, T.K.; Kabanov, A.V. Well-defined cross-linked antioxidant nanozymes for treatment of ischemic brain injury. J. Control. Release 2012, 162, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Burton, G.F.; McKinney, J.S.; Young, H.; Ellis, E.F. Brain and tissue distribution of polyethylene glycol-conjugated superoxide dismutase in rats. Stroke 1992, 23, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, T.; Tanaka, K.-I.; Tasaka, Y.; Namba, T.; Suzuki, J.; Ishihara, T.; Okamoto, S.; Hibi, T.; Takenaga, M.; Igarashi, R.; et al. Therapeutic effect of lecithinized superoxide dismutase against colitis. J. Pharmacol. Exp. Ther. 2009, 328, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.H.; Ji, X.H.; Zhao, N.; Song, C.X.; Wang, F.S.; Liu, C.H. The conjugation of Cu/Zn superoxide dismutase (SOD) to O-(2-hydroxyl) propyl-3-trimethyl ammonium chitosan chloride (O-HTCC) enhances its therapeutic potential against radiation-induced oxidative damage. Polym. Chem. 2016, 7, 1826–1835. [Google Scholar] [CrossRef]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Zhang, P.; Sun, F.; Tsao, C.; Liu, S.; Jain, P.; Sinclair, A.; Hung, H.C.; Bai, T.; Wu, K.; Jiang, S. Zwitterionic gel encapsulation promotes protein stability, enhances pharmacokinetics, and reduces immunogenicity. Proc. Natl. Acad. Sci. USA 2015, 112, 12046–12051. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Kim, S.H.; Decker, C.G.; Wong, D.Y.; Loo, J.A.; Maynard, H.D. A heparin-mimicking polymer conjugate stabilizes basic fibroblast growth factor. Nat. Chem. 2013, 5, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Fidalgo, S.; Cardeno, A.; Sanchez-Hidalgo, M.; Aparicio-Soto, M.; Villegas, I.; Rosillo, M.A.; de la Lastra, C.A. Dietary unsaponifiable fraction from extra virgin olive oil supplementation attenuates acute ulcerative colitis in mice. Eur. J. Pharm. Sci. 2013, 48, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Brune, B.; Dehne, N.; Grossmann, N.; Jung, M.; Namgaladze, D.; Schmid, T.; von Knethen, A.; Weigert, A. Redox control of inflammation in macrophages. Antioxid. Redox Signal. 2013, 19, 595–637. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Fuss, I.J. Proinflammatory cytokines in the pathogenesis of inflammatory bowel diseases. Gastroenterology 2011, 140, 1756–1767. [Google Scholar] [CrossRef] [PubMed]
- Mahida, Y.R.; Rolfe, V.E. Host-bacterial interactions in inflammatory bowel disease. Clin. Sci. 2004, 107, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Conner, E.M.; Brand, S.J.; Davis, J.M.; Kang, D.Y.; Grisham, M.B. Role of reactive metabolites of oxygen and nitrogen in inflammatory bowel disease: Toxins, mediators, and modulators of gene expression. Inflamm. Bowel Dis. 1996, 2, 133–147. [Google Scholar] [PubMed]
- Grisham, M.B. Oxidants and free radicals in inflammatory bowel disease. Lancet 1994, 344, 859–861. [Google Scholar] [CrossRef]
- Kruidenier, L.; Verspaget, H.W. Oxidative stress as a pathogenic factor in inflammatory bowel disease—Radicals or ridiculous? Aliment. Pharm. Ther. 2002, 16, 1997–2015. [Google Scholar] [CrossRef]
- Weigmann, B.; Neufert, C.; Neurath, M.F.; Wirtz, S. Chemically induced mouse models of intestinal inflammation. Nat. Protoc. 2007, 2, 541–546. [Google Scholar]
- Perše, M.; Cerar, A. Dextran sodium sulphate colitis mouse model: Traps and tricks. J. Biomed. Biotechnol. 2012, 2012, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Ota, H.; Sasagawa, S.; Sakatani, T.; Fujikura, T. Assay method for myeloperoxidase in human polymorphonuclear leukocytes. Anal. Biochem. 1983, 132, 345–352. [Google Scholar] [CrossRef]
- Wan, A.; Xu, Q.; Sun, Y.; Li, H. Antioxidant activity of high molecular weight chitosan and N,O-quaternized chitosans. J. Agric. Food Chem. 2013, 61, 6921–6928. [Google Scholar] [CrossRef] [PubMed]
- Curotto, E.; Aros, F. Quantitative determination of chitosan and the percentage of free amino groups. Anal. Biochem. 1993, 211, 240–241. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Garcia, L.; Herránz, S.; Luque, A.; Hortelano, S. Critical role of p38 MAPK in IL-4-induced alternative activation of peritoneal macrophages: Molecular immunology. Eur. J. Immunol. 2015, 45, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Salimuddin; Nagasaki, A.; Gotoh, T.; Isobe, H.; Mori, M. Regulation of the genes for arginase isoforms and related enzymes in mouse macrophages by lipopolysaccharide. Am. J. Physiol. 1999, 277, 110–117. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Yang, X.; Yao, H.; Chen, Y.; Sun, L.; Li, Y.; Ma, X.; Duan, S.; Li, X.; Xiang, R.; Han, J.; et al. Inhibition of glutathione production induces macrophage CD36 expression and enhances cellular-oxidized low density lipoprotein (OXLDL) uptake. J. Biol. Chem. 2015, 290, 21788–21799. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.S.; Murthy, S.N.S.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–250. [Google Scholar] [PubMed]
- Shin, J.S.; Cho, E.J.; Choi, H.E.; Seo, J.H.; An, H.J.; Park, H.J.; Cho, Y.W.; Lee, K.T. Anti-inflammatory effect of a standardized triterpenoid-rich fraction isolated from rubus coreanus on dextran sodium sulfate-induced acute colitis in mice and LPS-induced macrophages. J. Ethnopharmacol. 2014, 158 Pt A, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef] [PubMed]








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, N.; Feng, Z.; Shao, M.; Cao, J.; Wang, F.; Liu, C. Stability Profiles and Therapeutic Effect of Cu/Zn Superoxide Dismutase Chemically Coupled to O-Quaternary Chitosan Derivatives against Dextran Sodium Sulfate-Induced Colitis. Int. J. Mol. Sci. 2017, 18, 1121. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061121
Zhao N, Feng Z, Shao M, Cao J, Wang F, Liu C. Stability Profiles and Therapeutic Effect of Cu/Zn Superoxide Dismutase Chemically Coupled to O-Quaternary Chitosan Derivatives against Dextran Sodium Sulfate-Induced Colitis. International Journal of Molecular Sciences. 2017; 18(6):1121. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061121
Chicago/Turabian StyleZhao, Nan, Zhaolong Feng, Meng Shao, Jichao Cao, Fengshan Wang, and Chunhui Liu. 2017. "Stability Profiles and Therapeutic Effect of Cu/Zn Superoxide Dismutase Chemically Coupled to O-Quaternary Chitosan Derivatives against Dextran Sodium Sulfate-Induced Colitis" International Journal of Molecular Sciences 18, no. 6: 1121. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061121