
An Isoform of Nedd4-2 Plays a Pivotal Role in Electrophysiological Cardiac Abnormalities

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Cardiac Phenotypic Evaluations in Neural Precursor Cell-Expressed Developmentally Downregulated Gene 4-2 (Nedd4-2) C2 Knock out (KO) Mice
2.1.1. Comparison of Electrocardiographic and Echocardiographic Findings between Wild-Type and Nedd4-2 C2 Knockout Mice under Resting Conditions
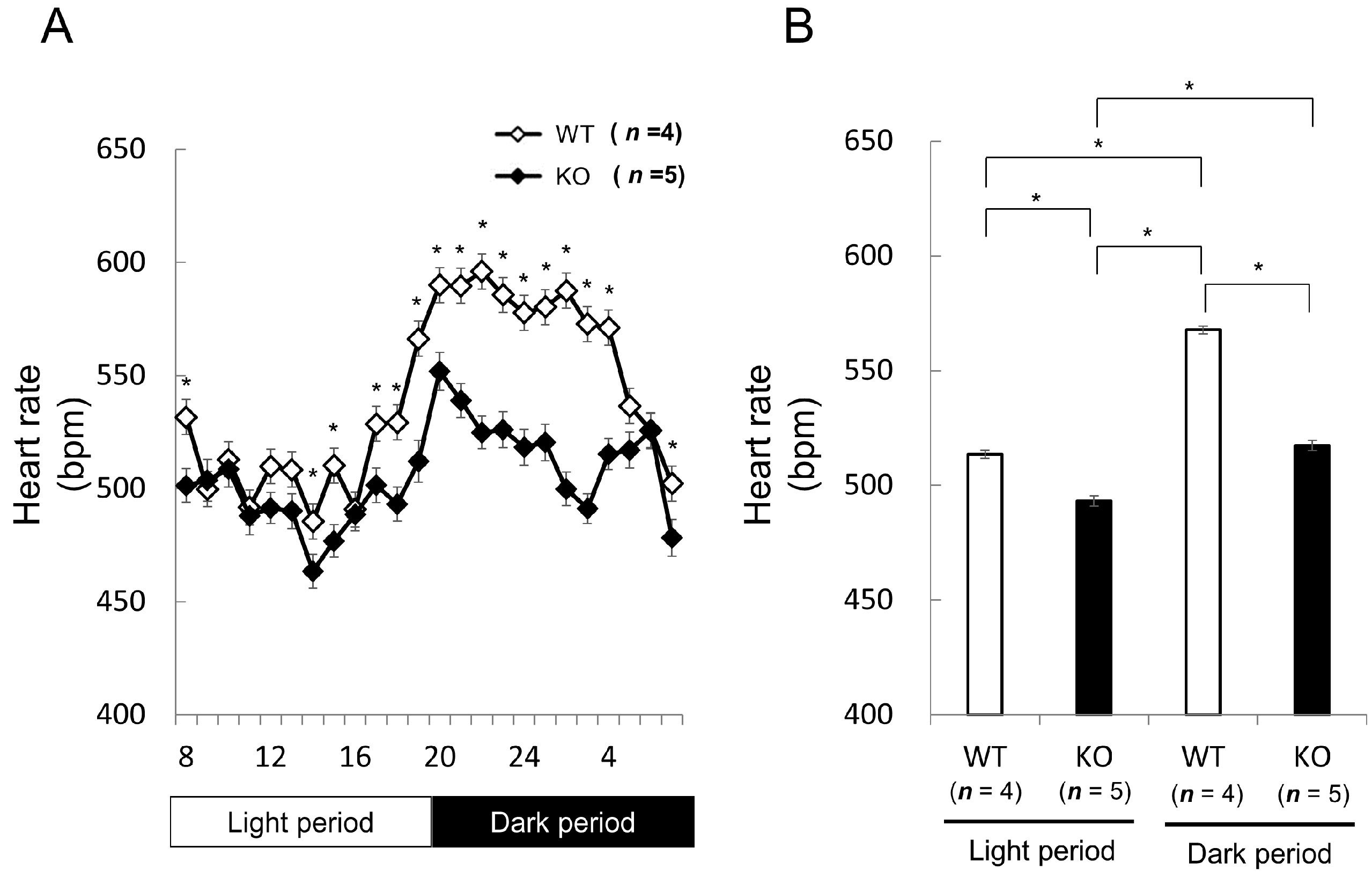
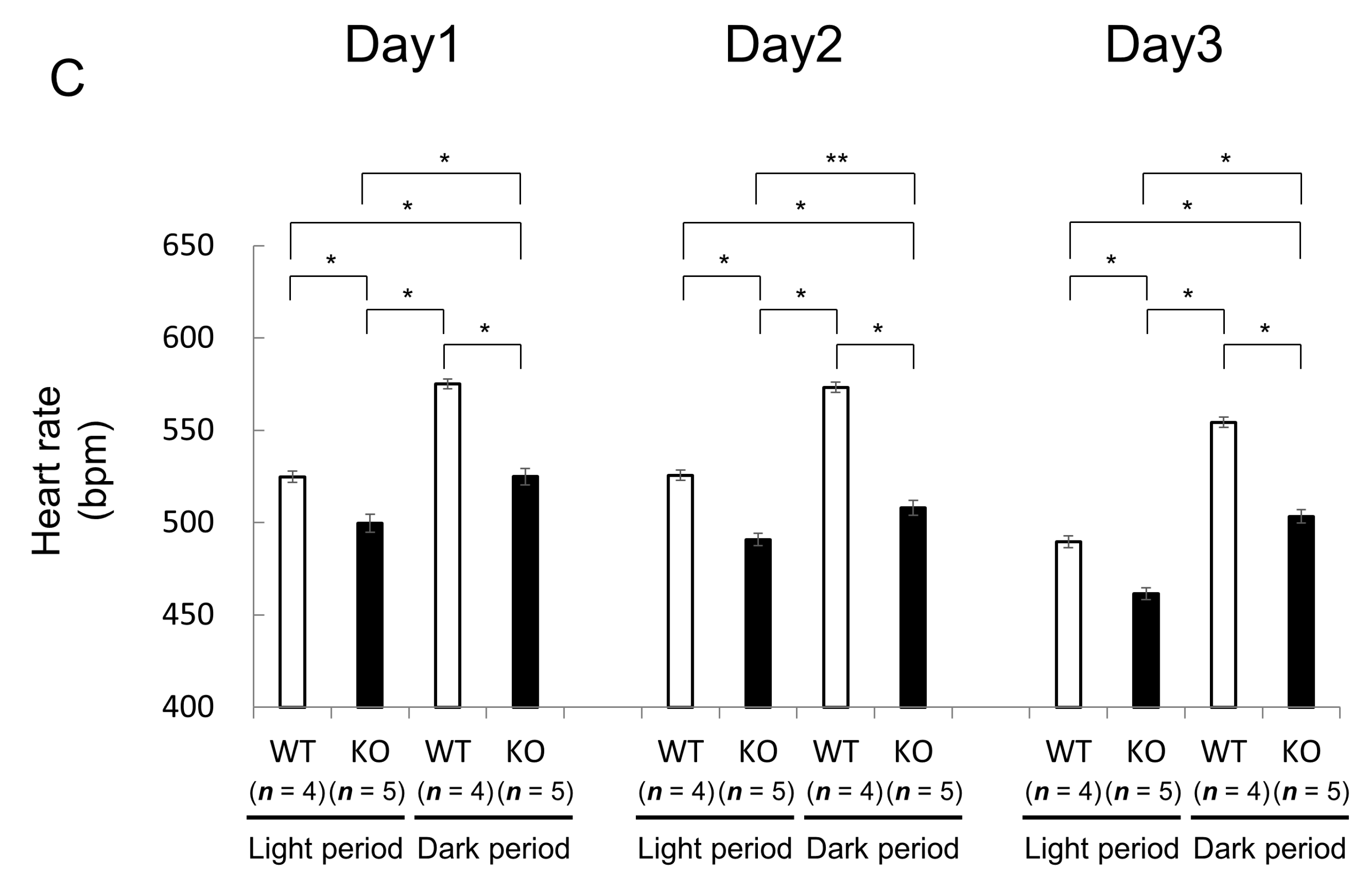
2.1.2. Comparison of Heart Rate and Heart Rate Variability between Wild-Type and Nedd4-2 C2 Knockout Mice in Resting Conditions
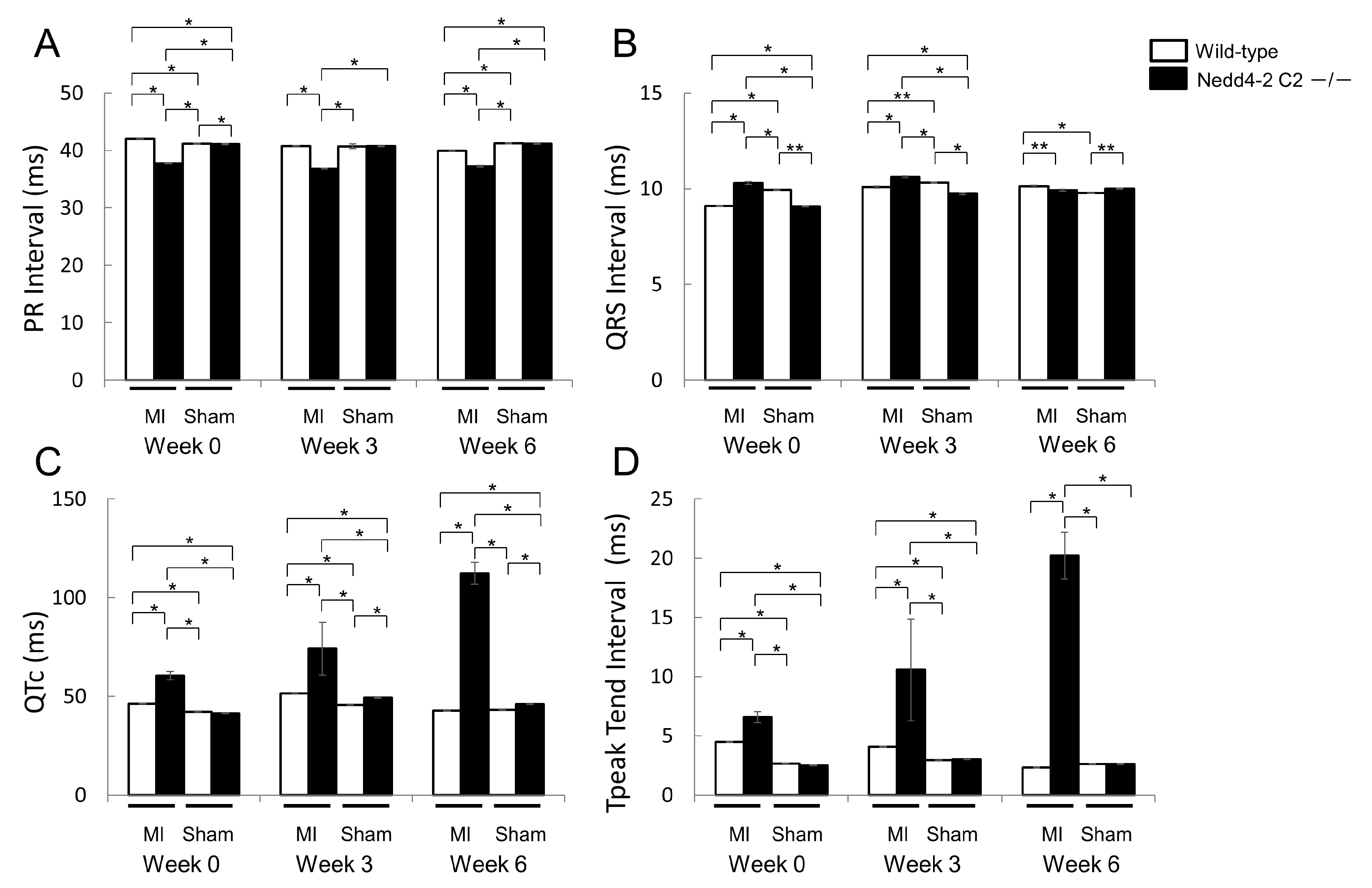
2.1.3. Comparison of Electrocardiographic and Echocardiographic Findings between Wild-Type and Nedd4-2 C2 Knockout Mice after Myocardial Infarction
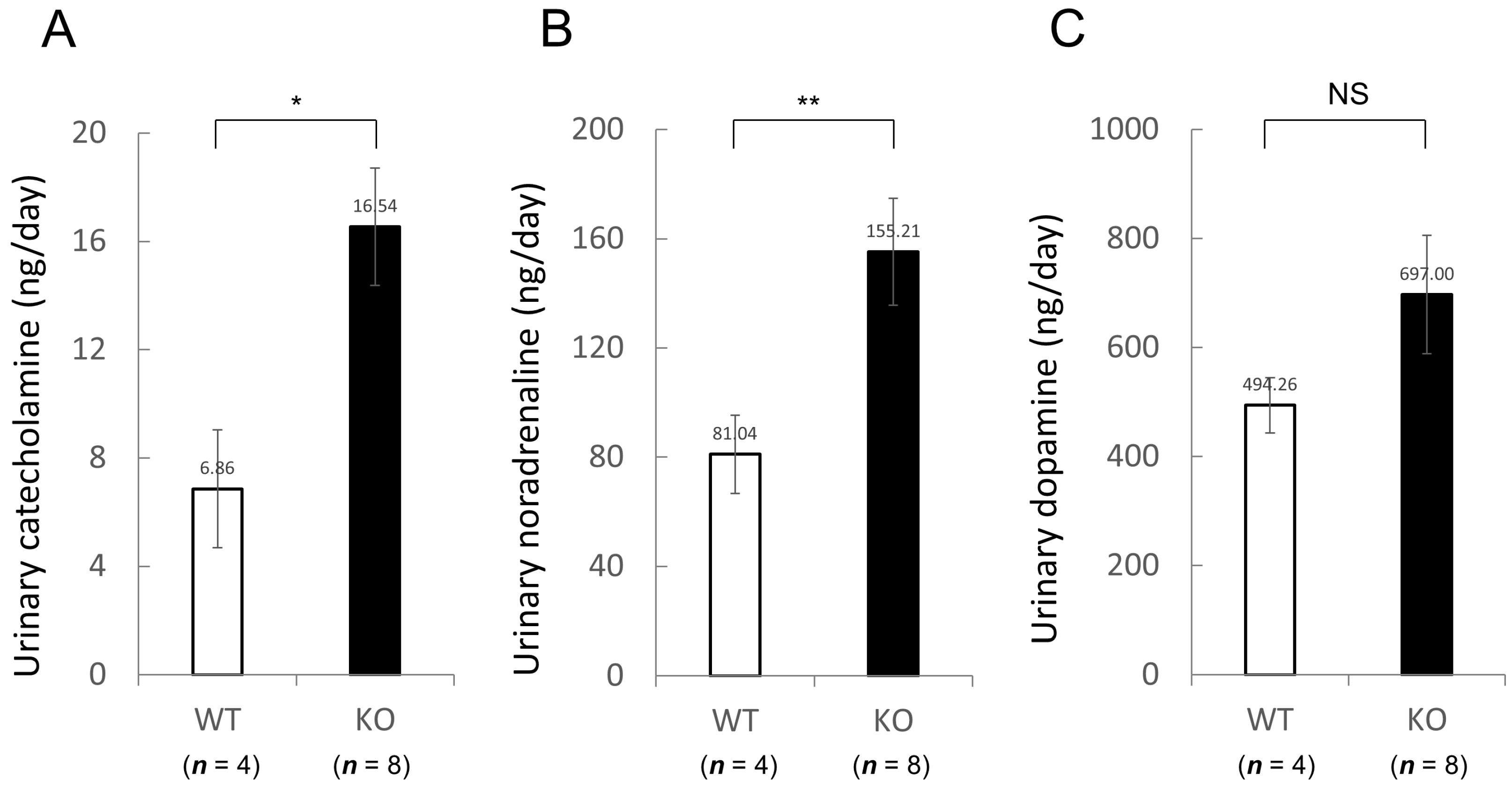
2.1.4. Comparison of Blood Chemistry and Urinalysis between Wild-Type and Nedd4-2 C2 Knockout Mice under Resting Conditions
3. Discussion
4. Materials and Methods
4.1. Generation of Nedd4-2 C2 Domain Knockout Mice
4.2. Electrocardiographic Analyses
4.3. Echocardiographic Analyses
4.4. Heart Rate and Heart Rate Variability Measurements
4.5. Examinations for Left Anterior Descending Artery Ligation Model
4.6. Catecholamine Determination
4.7. Blood Chemistry Analysis
4.8. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CKD | Chronic kidney disease |
| CVD | Cardiovascular disease |
| HRV | Heart rate variability |
| MI | Myocardial infarction |
| LAD | Left anterior descending (artery) |
References
- Tomiyama, H.; Yamashina, A. Vascular dysfunction: A key player in chronic cardio-renal syndrome. Intern. Med. 2015, 54, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Charytan, D.M.; Fishbane, S.; Malyszko, J.; McCullough, P.A.; Goldsmith, D. Cardiorenal syndrome and the role of the bone-mineral axis and anemia. Am. J. Kidney Dis. 2015, 66, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Schild, L.; Lu, Y.; Gautschi, I.; Schneeberger, E.; Lifton, R.P.; Rossier, B.C. Identification of a PY motif in the epithelial Na channel subunits as a target sequence for mutations causing channel activation found in Liddle syndrome. EMBO J. 1996, 15, 2381–2387. [Google Scholar] [PubMed]
- Dunn, D.M.; Ishigami, T.; Pankow, J.; von Niederhausern, A.; Alder, J.; Hunt, S.C.; Leppert, M.F.; Lalouel, J.M.; Weiss, R.B. Common variant of human Nedd4L activates a cryptic splice site to form a frameshifted transcript. J. Hum. Genet. 2002, 47, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Umemura, M.; Ishigami, T.; Tamura, K.; Sakai, M.; Miyagi, Y.; Nagahama, K.; Aoki, I.; Uchino, K.; Rohrwasser, A.; Lalouel, J.M.; et al. Transcriptional diversity and expression of Nedd4L gene in distal nephron. Biochem. Biophys. Res. Commun. 2006, 339, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, T.; Araki, N.; Minegishi, S.; Umemura, M.; Umemura, S. Genetic variation in Nedd4L, salt sensitivity, and hypertension: Human Nedd4L rs4149601 G allele generates evolutionary new isoform I with C2 domain. J. Hypertens. 2014, 32, 1905. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, T.; Araki, N.; Umemura, S. Human Nedd4L rs4149601 G allele generates evolutionary new isoform I with C2 domain. Hypertension 2010, 55, e10–e11. [Google Scholar] [CrossRef] [PubMed]
- Araki, N.; Umemura, M.; Miyagi, Y.; Yabana, M.; Miki, Y.; Tamura, K.; Uchino, K.; Aoki, R.; Goshima, Y.; Umemura, S.; et al. Expression, transcription, and possible antagonistic interaction of the human Nedd4L gene variant: Implications for essential hypertension. Hypertension 2008, 51, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Minegishi, S.; Ishigami, T.; Kino, T.; Chen, L.; Nakashima-Sasaki, R.; Araki, N.; Yatsu, K.; Fujita, M.; Umemura, S. An isoform of Nedd4-2 is critically involved in the renal adaptation to high salt intake in mice. Sci. Rep. 2016, 6, 27137. [Google Scholar] [CrossRef] [PubMed]
- Modell, S.M.; Lehmann, M.H. The long QT syndrome family of cardiac ion channelopathies: A HuGE review. Genet. Med. 2006, 8, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Abriel, H. Roles and regulation of the cardiac sodium channel Na v 1.5: Recent insights from experimental studies. Cardiovasc. Res. 2007, 76, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.Q.; Yan, C.C.; Zhang, X.; Yan, M.; Liu, L.R.; Geng, H.Z.; Lv, L.; Li, B.X. Mechanisms underlying probucol-induced hERG-channel deficiency. Drug. Des. Dev. Ther. 2015, 9, 3695–3704. [Google Scholar]
- Kang, Y.; Guo, J.; Yang, T.; Li, W.; Zhang, S. Regulation of the human ether-a-go-go-related gene (hERG) potassium channel by Nedd4 family interacting proteins (Ndfips). Biochem. J. 2015, 472, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, T.; Li, X.; Shallow, H.; Yang, T.; Li, W.; Xu, J.; Fridman, M.D.; Yang, X.; Zhang, S. Cell surface expression of human ether-a-go-go-related gene (hERG) channels is regulated by caveolin-3 protein via the ubiquitin ligase Nedd4-2. J. Biol. Chem. 2012, 287, 33132–33141. [Google Scholar] [CrossRef] [PubMed]
- Thireau, J.; Zhang, B.L.; Poisson, D.; Babuty, D. Heart rate variability in mice: A theoretical and practical guide. Exp. Physiol. 2008, 93, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Kuwahara, M.; Tsubone, H.; Sugano, S. Autonomic nervous function in mice and voles (Microtus arvalis): Investigation by power spectral analysis of heart rate variability. Lab. Anim. 1996, 30, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Jentzer, J.C.; Chawla, L.S. A clinical approach to the acute cardiorenal syndrome. Crit. Care Clin. 2015, 31, 685–703. [Google Scholar] [CrossRef] [PubMed]
- Kazory, A.; Elkayam, U. Cardiorenal interactions in acute decompensated heart failure: Contemporary concepts facing emerging controversies. J. Card. Fail. 2014, 20, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Lifton, R.P.; Gharavi, A.G.; Geller, D.S. Molecular mechanisms of human hypertension. Cell 2001, 104, 545–556. [Google Scholar] [CrossRef]
- Lifton, R.P. Genetic determinants of human hypertension. Proc. Natl. Acad. Sci. USA 1995, 92, 8545–8551. [Google Scholar] [CrossRef] [PubMed]
- Spoonamore, K.G.; Ware, S.M. Genetic testing and genetic counseling in patients with sudden death risk due to heritable arrhythmias. Heart Rhythm. 2016, 13, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Curran, M.E.; Splawski, I.; Burn, T.C.; Millholland, J.M.; VanRaay, T.J.; Shen, J.; Timothy, K.W.; Vincent, G.M.; de Jager, T.; et al. Positional cloning of a novel potassium channel gene: KVLQT1 mutations cause cardiac arrhythmias. Nat. Genet. 1996, 12, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Curran, M.E.; Splawski, I.; Timothy, K.W.; Vincent, G.M.; Green, E.D.; Keating, M.T. A molecular basis for cardiac arrhythmia: hERG mutations cause long QT syndrome. Cell 1995, 80, 795–803. [Google Scholar] [CrossRef]
- Wang, Q.; Shen, J.; Splawski, I.; Atkinson, D.; Li, Z.; Robinson, J.L.; Moss, A.J.; Towbin, J.A.; Keating, M.T. SCN5A mutations associated with an inherited cardiac arrhythmia, long QT syndrome. Cell 1995, 80, 805–811. [Google Scholar] [CrossRef]
- Brugada, R.; Hong, K.; Dumaine, R.; Cordeiro, J.; Gaita, F.; Borggrefe, M.; Menendez, T.M.; Brugada, J.; Pollevick, G.D.; Wolpert, C.; et al. Sudden death associated with short-QT syndrome linked to mutations in hERG. Circulation 2004, 109, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Kirsch, G.E.; Zhang, D.; Brugada, R.; Brugada, J.; Brugada, P.; Potenza, D.; Moya, A.; Borggrefe, M.; Breithardt, G.; et al. Genetic basis and molecular mechanism for idiopathic ventricular fibrillation. Nature 1998, 392, 293–296. [Google Scholar] [PubMed]
- Benson, D.W.; Wang, D.W.; Dyment, M.; Knilans, T.K.; Fish, F.A.; Strieper, M.J.; Rhodes, T.H.; George, A.L., Jr. Congenital sick sinus syndrome caused by recessive mutations in the cardiac sodium channel gene (SCN5A). J. Clin. Investig. 2003, 112, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Schott, J.J.; Alshinawi, C.; Kyndt, F.; Probst, V.; Hoorntje, T.M.; Hulsbeek, M.; Wilde, A.A.; Escande, D.; Mannens, M.M.; Le Marec, H. Cardiac conduction defects associate with mutations in SCN5A. Nat. Genet. 1999, 23, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Dahlberg, J.; Nilsson, L.O.; von Wowern, F.; Melander, O. Polymorphism in Nedd4L is associated with increased salt sensitivity, reduced levels of P-renin and increased levels of Nt-proANP. PLoS ONE 2007, 2, e432. [Google Scholar] [CrossRef] [PubMed]
- Fava, C.; von Wowern, F.; Berglund, G.; Carlson, J.; Hedblad, B.; Rosberg, L.; Burri, P.; Almgren, P.; Melander, O. 24-h ambulatory blood pressure is linked to chromosome 18q21-22 and genetic variation of Nedd4L associates with cross-sectional and longitudinal blood pressure in Swedes. Kidney Int. 2006, 70, 562–569. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ECG Waveform | Wild-Type | Nedd4-2 C2 −/− | p |
|---|---|---|---|
| (n = 5) | (n = 5) | ||
| PR Interval (ms) | 40.81 ± 0.10 | 40.38 ± 0.12 | 0.0104 |
| P Duration (ms) | 15.00 ± 0.34 | 17.46 ± 0.28 | <0.0001 |
| QRS Interval (ms) | 8.79 ± 0.02 | 9.97 ± 0.04 | <0.0001 |
| QT Interval (ms) | 13.34 ± 0.03 | 18.18 ± 0.44 | <0.0001 |
| QTc (ms) | 38.97 ± 0.08 | 53.52 ± 1.27 | <0.0001 |
| UCG Measurements | Wild-Type | Nedd4-2 C2 −/− | p |
|---|---|---|---|
| (n = 5) | (n = 5) | ||
| IVSTd (mm) | 0.82 ± 0.02 | 0.81 ± 0.02 | 0.7717 |
| LVIDd (mm) | 4.28 ± 0.07 | 4.29 ± 0.04 | 0.9484 |
| LVPWTd (mm) | 0.73 ± 0.02 | 0.75 ± 0.02 | 0.3991 |
| IVSTs (mm) | 1.21 ± 0.03 | 1.15 ± 0.04 | 0.2290 |
| LVIDs (mm) | 3.00 ± 0.08 | 3.00 ± 0.05 | 0.9781 |
| LVPWTs (mm) | 0.92 ± 0.02 | 0.97 ± 0.02 | 0.0961 |
| EDV (μL) | 80.14 ± 3.99 | 79.08 ± 2.29 | 0.8291 |
| ESV (μL) | 28.76 ± 2.35 | 27.58 ± 1.28 | 0.6799 |
| SV (μL) | 51.38 ± 2.28 | 51.50 ± 1.69 | 0.9675 |
| CO (mL/min) | 24.95 ± 1.12 | 24.44 ± 0.89 | 0.7310 |
| EF (%) | 65.20 ± 1.68 | 65.21 ± 1.28 | 0.9955 |
| FS (%) | 30.16 ± 1.14 | 29.96 ± 0.86 | 0.8946 |
| HR (bpm) | 485.79 ± 5.33 | 473.46 ± 4.11 | 0.0819 |
| UCG Measrements | Wild-Type | Nedd4-2 C2 −/− | Wild-Type | Nedd4-2 C2 −/− | p |
|---|---|---|---|---|---|
| MI | MI | Sham | Sham | ||
| (n = 4) | (n = 4) | (n = 4) | (n = 4) | (ANOVA) | |
| Week 0 | |||||
| IVSTd (mm) | 0.73 ± 0.02 | 0.72 ± 0.02 | 0.72 ± 0.02 | 0.75 ± 0.01 | 0.5464 |
| SV (μL) | 44.59 ± 3.69 | 31.67 ± 1.51 a | 45.80 ± 2.38 | 41.88 ± 1.18 | 0.0002 |
| CO (mL/min) | 22.81 ± 2.09 | 16.22 ± 0.75 a | 22.40 ± 1.28 | 20.97 ± 0.62 | 0.0017 |
| EF (%) | 63.57 ± 1.84 | 67.99 ± 1.17 | 68.25 ± 1.52 | 66.67 ± 1.18 | 0.1427 |
| FS (%) | 29.00 ± 1.15 | 31.76 ± 0.81 | 32.16 ± 1.07 | 31.14 ± 0.87 | 0.1860 |
| Week 3 | |||||
| IVSTd (mm) | 0.70 ± 0.03 | 0.69 ± 0.02 b | 0.76 ± 0.01 | 0.75 ± 0.01 | 0.0327 |
| SV (μL) | 57.07 ± 4.85 | 51.09 ± 1.87 | 50.67 ± 4.20 | 49.50 ± 1.77 | 0.3844 |
| CO (mL/min) | 27.42 ± 2.26 | 25.29 ± 0.90 | 24.44 ± 0.89 | 24.95 ± 1.12 | 0.3852 |
| EF (%) | 69.16 ± 1.23 | 68.81 ± 1.09 | 67.99 ± 3.13 | 68.17 ± 1.14 | 0.9708 |
| FS (%) | 32.60 ± 0.86 | 32.44 ± 0.76 | 33.68 ± 0.58 | 32.00 ± 0.80 | 0.4587 |
| Week 6 | |||||
| IVSTd (mm) | 0.75 ± 0.02 | 0.74 ± 0.02 | 0.77 ± 0.02 | 0.77 ± 0.02 | 0.5427 |
| SV (μL) | 58.81 ± 5.38 | 48.68 ± 1.62 | 54.80 ± 2.38 | 58.17 ± 2.69 | 0.1224 |
| CO (mL/min) | 29.93 ± 2.86 | 24.33 ± 0.66 | 27.05 ± 1.22 | 28.37 ± 1.39 | 0.1350 |
| EF (%) | 69.12 ± 1.11 | 70.45 ± 0.89 | 70.00 ± 0.89 | 69.86 ± 0.99 | 0.8338 |
| FS (%) | 32.63 ± 0.77 | 33.39 ± 0.63 | 33.35 ± 0.66 | 33.20 ± 0.73 | 0.8751 |
| Serum Markers | Wild-Type | Nedd4L C2 −/− | p |
|---|---|---|---|
| (n = 8) | (n = 9) | ||
| Serum Cr (mg/dL) | 0.16 ± 0.01 | 0.16 ± 0.01 | 0.6433 |
| Serum Na (mEq/L) | 147.88 ± 1.36 | 144.56 ± 1.07 | 0.0708 |
| Serum Ca (mg/dL) | 9.19 ± 0.25 | 8.50 ± 0.21 | 0.0506 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minegishi, S.; Ishigami, T.; Kawamura, H.; Kino, T.; Chen, L.; Nakashima-Sasaki, R.; Doi, H.; Azushima, K.; Wakui, H.; Chiba, Y.; et al. An Isoform of Nedd4-2 Plays a Pivotal Role in Electrophysiological Cardiac Abnormalities. Int. J. Mol. Sci. 2017, 18, 1268. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061268
Minegishi S, Ishigami T, Kawamura H, Kino T, Chen L, Nakashima-Sasaki R, Doi H, Azushima K, Wakui H, Chiba Y, et al. An Isoform of Nedd4-2 Plays a Pivotal Role in Electrophysiological Cardiac Abnormalities. International Journal of Molecular Sciences. 2017; 18(6):1268. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061268
Chicago/Turabian StyleMinegishi, Shintaro, Tomoaki Ishigami, Hisho Kawamura, Tabito Kino, Lin Chen, Rie Nakashima-Sasaki, Hiroshi Doi, Kengo Azushima, Hiromichi Wakui, Yumi Chiba, and et al. 2017. "An Isoform of Nedd4-2 Plays a Pivotal Role in Electrophysiological Cardiac Abnormalities" International Journal of Molecular Sciences 18, no. 6: 1268. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061268