Role of Omentin, Vaspin, Cardiotrophin-1, TWEAK and NOV/CCN3 in Obesity and Diabetes Development

 , , ,
, , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Omentin
2.1. Omentin and Obesity
2.2. Omentin and Alterations in Glycemic Control
2.3. Signaling Pathways Involved in the Metabolic Actions of Omentin
3. Vaspin
3.1. Vaspin and Obesity
3.2. Vaspin and Alterations in Glycemic Control
3.3. Signaling Pathways Involved in the Metabolic Actions of Vaspin
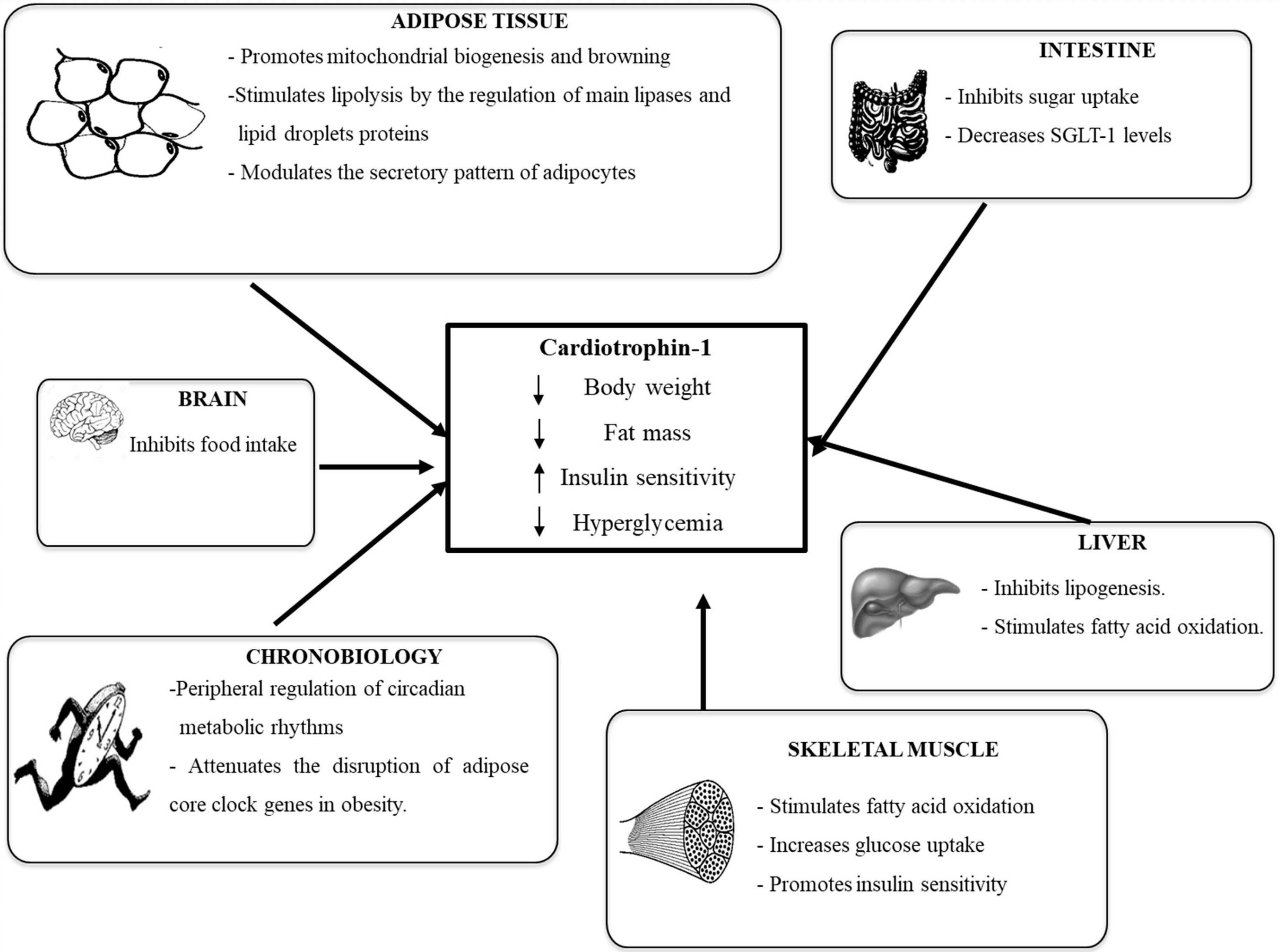
4. Cardiotrophin-1
4.1. Cardiotrophin-1 and Obesity
4.2. Cardiotrophin-1 and Alterations in Glycemic Control
4.3. CT-1 Receptor and Signaling Pathways Involved in the Metabolic Actions of CT-1
5. TWEAK
5.1. TWEAK, Obesity and Alterations in Glycemic Control
5.2. TWEAK Aeceptors and Signaling Pathways Involved in the Metabolic Actions of TWEAK
6. NOV/CCN3
6.1. NOV/CCN3 and Obesity
6.2. NOV/CCN3 and Alterations in Glycemic Control
7. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Ezquerro, S.; Méndez-Giménez, L.; Becerril, S.; Frühbeck, G. Revisiting the adipocyte: A model for integration of cytokine signaling in the regulation of energy metabolism. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E691–E714. [Google Scholar] [CrossRef] [PubMed]
- Dusserre, E.; Moulin, P.; Vidal, H. Differences in mrna expression of the proteins secreted by the adipocytes in human subcutaneous and visceral adipose tissues. Biochim. Biophys. Acta 2000, 1500, 88–96. [Google Scholar] [CrossRef]
- Frühbeck, G.; Gómez-Ambrosi, J. Adipose tissue structure function and metabolism. In Encyclopedia of Human Nutrition; Caballero, B., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1–13. [Google Scholar]
- Yang, R.; Xu, A.; Pray, J.; Hu, H.; Jadhao, S.; Hansen, B.; Shuldiner, A.; Mclenithan, J.; Gong, D. Cloning of omentin, a new adipocytokine from omental fat tissue in humans. Diabetes 2003, 52, A1. [Google Scholar]
- Alexander, R.W. Understanding adipose-derived stromal vascular fraction (ad-svf) cell biology and use on the basis of cellular, chemical, structural and paracrine components: A concise review. J. Prolother. 2012, 4, 855–869. [Google Scholar]
- Yang, R.Z.; Lee, M.J.; Hu, H.; Pray, J.; Wu, H.B.; Hansen, B.C.; Shuldiner, A.R.; Fried, S.K.; McLenithan, J.C.; Gong, D.W. Identification of omentin as a novel depot-specific adipokine in human adipose tissue: Possible role in modulating insulin action. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1253–E1261. [Google Scholar] [CrossRef] [PubMed]
- Komiya, T.; Tanigawa, Y.; Hirohashi, S. Cloning of the novel gene intelectin, which is expressed in intestinal paneth cells in mice. Biochem. Biophys. Res. Commun. 1998, 251, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Mao, F.; Gong, D.W.; Damcott, C.; Sabra, M.; Yang, R.; Pollin, T.I.; Tanner, K.; Ott, S.; Mclenithan, J.C.; Fried, S.; et al. Systematic analysis of omentin 1 and omentin 2 on 1q23 as candidate genes for type 2 diabetes in the old order amish. Diabetes 2004, 53–59. [Google Scholar]
- Lee, J.A.; Park, H.S.; Song, Y.S.; Jang, Y.J.; Kim, J.H.; Lee, Y.J.; Heo, Y.S. Relationship between vaspin gene expression and abdominal fat distribution of korean women. Endocr. J. 2011, 58, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Díaz, A.; Arana-Martínez, J.C.; Carbó, R.; Espinosa-Cervantes, R.; Sánchez-Muñoz, F. Omentin: Role in insulin resistance, inflammation and cardiovascular protection. Arch. Cardiol. Mex. 2016, 86, 233–243. [Google Scholar] [PubMed]
- Kuperman, D.A.; Lewis, C.C.; Woodruff, P.G.; Rodriguez, M.W.; Yang, Y.H.; Dolganov, G.M.; Fahy, J.V.; Erle, D.J. Dissecting asthma using focused transgenic modeling and functional genomics. J. Allergy Clin. Immunol. 2005, 116, 305–311. [Google Scholar] [CrossRef] [PubMed]
- De Souza Batista, C.M.; Yang, R.-Z.; Lee, M.-J.; Glynn, N.M.; Yu, D.-Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin plasma levels and gene expression are decreased in obesity. Diabetes 2007, 56, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Adya, R.; Farhatullah, S.; Lewandowski, K.C.; O'Hare, P.; Lehnert, H.; Randeva, H.S. Omentin-1, a novel adipokine, is decreased in overweight insulin-resistant women with polycystic ovary syndrome: Ex vivo and in vivo regulation of omentin-1 by insulin and glucose. Diabetes 2008, 57, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Auguet, T.; Quintero, Y.; Riesco, D.; Morancho, B.; Terra, X.; Crescenti, A.; Broch, M.; Aguilar, C.; Olona, M.; Porras, J.A.; et al. New adipokines vaspin and omentin. Circulating levels and gene expression in adipose tissue from morbidly obese women. BMC Med. Genet. 2011, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Cătoi, A.F.; Suciu, Ş.; Pârvu, A.E.; Copăescu, C.; Galea, R.F.; Buzoianu, A.D.; Vereşiu, I.A.; Cătoi, C.; Pop, I.D. Increased chemerin and decreased omentin-1 levels in morbidly obese patients are correlated with insulin resistance, oxidative stress and chronic inflammation. Clujul Med. 2014, 87, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Oświęcimska, J.; Suwała, A.; Świętochowska, E.; Ostrowska, Z.; Gorczyca, P.; Ziora-Jakutowicz, K.; Machura, E.; Szczepańska, M.; Kukla, M.; Stojewska, M.; et al. Serum omentin levels in adolescent girls with anorexia nervosa and obesity. Physiol. Res. 2015, 64, 701–709. [Google Scholar] [PubMed]
- Catli, G.; Anik, A.; Abaci, A.; Kume, T.; Bober, E. Low omentin-1 levels are related with clinical and metabolic parameters in obese children. Exp. Clin. Endocrinol. Diabetes 2013, 121, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Prats-Puig, A.; Bassols, J.; Bargalló, E.; Mas-Parareda, M.; Ribot, R.; Soriano-Rodríguez, P.; Berengüí, À.; Díaz, M.; de Zegher, F.; Ibánez, L.; et al. Toward an early marker of metabolic dysfunction: Omentin-1 in prepubertal children. Obesity 2011, 19, 1905–1907. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Catalán, V.; Ortega, F.; Gómez-Ambrosi, J.; Ricart, W.; Frühbeck, G.; Fernández-Real, J.M. Circulating omentin concentration increases after weight loss. Nutr. Metab. 2010, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Urbanová, M.; Dostálová, I.; Trachta, P.; Drápalová, J.; Kaválková, P.; Haluzíková, D.; Matoulek, M.; Lacinová, Z.; Mráz, M.; Kasalický, M.; et al. Serum concentrations and subcutaneous adipose tissue mrna expression of omentin in morbid obesity and type 2 diabetes mellitus: The effect of very-low-calorie diet, physical activity and laparoscopic sleeve gastrectomy. Physiol. Res. 2014, 63, 207–218. [Google Scholar] [PubMed]
- Saremi, A.; Asghari, M.; Ghorbani, A. Effects of aerobic training on serum omentin-1 and cardiometabolic risk factors in overweight and obese men. J. Sports Sci. 2010, 28, 993–998. [Google Scholar] [CrossRef] [PubMed]
- AminiLari, Z.; Fararouei, M.; Amanat, S.; Sinaei, E.; Dianatinasab, S.; AminiLari, M.; Daneshi, N.; Dianatinasab, M. The effect of 12 weeks aerobic, resistance, and combined exercises on omentin-1 levels and insulin resistance among type 2 diabetic middle-aged women. Diabetes Metab. J. 2017, 41, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Zehsaz, F.; Farhangi, N.; Ghahramani, M. The response of circulating omentin-1 concentration to 16-week exercise training in male children with obesity. Phys. Sportsmed. 2016, 44, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Faramarzi, M.; Banitalebi, E.; Nori, S.; Farzin, S.; Taghavian, Z. Effects of rhythmic aerobic exercise plus core stability training on serum omentin, chemerin and vaspin levels and insulin resistance of overweight women. J. Sports Med. Phys. Fit. 2016, 56, 476–482. [Google Scholar]
- Jialal, I.; Devaraj, S.; Kaur, H.; Adams-Huet, B.; Bremer, A.A. Increased chemerin and decreased omentin-1 in both adipose tissue and plasma in nascent metabolic syndrome. J. Clin. Endocrinol. Metab. 2013, 98, E514–E517. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.J.; Wang, J.P.; Liu, X.T.; Zheng, Q.S.; Xue, Y.S.; Wang, B.; Zhao, L.Y. Serum omentin-1 levels are inversely associated with the presence and severity of coronary artery disease in patients with metabolic syndrome. Biomarkers 2011, 16, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Vu, A.; Sidhom, M.S.; Bredbeck, B.C.; Kosmiski, L.A.; Aquilante, C.L. Evaluation of the relationship between circulating omentin-1 concentrations and components of the metabolic syndrome in adults without type 2 diabetes or cardiovascular disease. Diabetol. Metab. Syndr. 2014, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Y.; Guo, L.; Li, Q. Changes of serum omentin-1 levels in normal subjects and in patients with impaired glucose regulation and with newly diagnosed and untreated type 2 diabetes. Diabetes Res. Clin. Pract. 2010, 88, 29–33. [Google Scholar] [CrossRef] [PubMed]
- El-Mesallamy, H.O.; El-Derany, M.O.; Hamdy, N.M. Serum omentin-1 and chemerin levels are interrelated in patients with type 2 diabetes mellitus with or without ischaemic heart disease. Diabet. Med. 2011, 28, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Akour, A.; Kasabri, V.; Boulatova, N.; Bustanji, Y.; Naffa, R.; Hyasat, D.; Khawaja, N.; Bustanji, H.; Zayed, A.; Momani, M. Levels of metabolic markers in drug-naive prediabetic and type 2 diabetic patients. Acta Diabetol. 2017, 54, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Wittenbecher, C.; Menzel, J.; Carstensen-Kirberg, M.; Biemann, R.; di Giuseppe, R.; Fritsche, A.; Isermann, B.; Herder, C.; Aleksandrova, K.; Boeing, H.; et al. Omentin-1, adiponectin, and the risk of developing type 2 diabetes. Diabetes Care 2016, 39, e79–e80. [Google Scholar] [CrossRef] [PubMed]
- Polkowska, A.; Szczepaniak, I.; Bossowski, A. Assessment of serum concentrations of ghrelin, obestatin, omentin-1, and apelin in children with type 1 diabetes. BioMed Res. Int. 2016, 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, L.; Di Nisio, C.; Recinella, L.; Chiavaroli, A.; Leone, S.; Ferrante, C.; Orlando, G.; Vacca, M. Effects of vaspin, chemerin and omentin-1 on feeding behavior and hypothalamic peptide gene expression in the rat. Peptides 2011, 32, 1866–1871. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, L.; Orlando, G.; Ferrante, C.; Recinella, L.; Leone, S.; Chiavaroli, A.; Di Nisio, C.; Shohreh, R.; Manippa, F.; Ricciuti, A.; et al. Orexigenic effects of omentin-1 related to decreased cart and crh gene expression and increased norepinephrine synthesis and release in the hypothalamus. Peptides 2013, 44, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Eichelmann, F.; Rudovich, N.; Pfeiffer, A.F.; Schulze, M.B.; Giuseppe, R.D.; Boeing, H.; Aleksandrova, K. Novel adipokines: Methodological utility in human obesity research. Int. J. Obes. 2017, 41, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Heiker, J.T. Vaspin (serpina12) in obesity, insulin resistance, and inflammation. J. Pept. Sci. 2014, 20, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Hida, K.; Wada, J.; Zhang, H.; Hiragushi, K.; Tsuchiyama, Y.; Shikata, K.; Makino, H. Identification of genes specifically expressed in the accumulated visceral adipose tissue of OLETF rats. J. Lipid Res. 2000, 41, 1615–1622. [Google Scholar] [PubMed]
- Hida, K.; Wada, J.; Eguchi, J.; Zhang, H.; Baba, M.; Seida, A.; Hashimoto, I.; Okada, T.; Yasuhara, A.; Nakatsuka, A.; et al. Visceral adipose tissue-derived serine protease inhibitor: A unique insulin-sensitizing adipocytokine in obesity. Proc. Natl. Acad. Sci. USA 2005, 102, 10610–10615. [Google Scholar] [CrossRef] [PubMed]
- Körner, A.; Neef, M.; Friebe, D.; Erbs, S.; Kratzsch, J.; Dittrich, K.; Blüher, S.; Kapellen, T.M.; Kovacs, P.; Stumvoll, M.; et al. Vaspin is related to gender, puberty and deteriorating insulin sensitivity in children. Int. J. Obes. 2011, 35, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Goktas, Z.; Owens, S.; Boylan, M.; Syn, D.; Shen, C.L.; Reed, D.B.; San Francisco, S.; Wang, S. Associations between tissue visfatin/nicotinamide, phosphoribosyltransferase (Nampt), retinol binding protein-4, and vaspin concentrations and insulin resistance in morbidly obese subjects. Mediat. Inflamm. 2013, 2013, 861496. [Google Scholar] [CrossRef] [PubMed]
- Klöting, N.; Berndt, J.; Kralisch, S.; Kovacs, P.; Fasshauer, M.; Schön, M.R.; Stumvoll, M.; Blüher, M. Vaspin gene expression in human adipose tissue: Association with obesity and type 2 diabetes. Biochem. Biophys. Res. Commun. 2006, 339, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Youn, B.S.; Klöting, N.; Kratzsch, J.; Lee, N.; Park, J.W.; Song, E.S.; Ruschke, K.; Oberbach, A.; Fasshauer, M.; Stumvoll, M.; et al. Serum vaspin concentrations in human obesity and type 2 diabetes. Diabetes 2008, 57, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Seeger, J.; Ziegelmeier, M.; Bachmann, A.; Lössner, U.; Kratzsch, J.; Blüher, M.; Stumvoll, M.; Fasshauer, M. Serum levels of the adipokine vaspin in relation to metabolic and renal parameters. J. Clin. Endocrinol. Metab. 2008, 93, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Von Loeffelholz, C.; Möhlig, M.; Arafat, A.M.; Isken, F.; Spranger, J.; Mai, K.; Randeva, H.S.; Pfeiffer, A.F.; Weickert, M.O. Circulating vaspin is unrelated to insulin sensitivity in a cohort of nondiabetic humans. Eur. J. Endocrinol. 2010, 162, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Kwak, S.H.; Lee, Y.; Moon, M.K.; Lim, S.; Park, Y.J.; Jang, H.C.; Kim, M.S. Plasma vaspin concentrations are elevated in metabolic syndrome in men and are correlated with coronary atherosclerosis in women. Clin. Endocrinol. 2011, 75, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.; Mirzaei, K.; Abdurahman, A.A.; Keshavarz, S.A.; Hossein-Nezhad, A. Mediatory effect of circulating vaspin on resting metabolic rate in obese individuals. Eur. J. Nutr. 2016, 55, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Aust, G.; Richter, O.; Rohm, S.; Kerner, C.; Hauss, J.; Klöting, N.; Ruschke, K.; Kovacs, P.; Youn, B.S.; Blüher, M. Vaspin serum concentrations in patients with carotid stenosis. Atherosclerosis 2009, 204, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Suleymanoglu, S.; Tascilar, E.; Pirgon, O.; Tapan, S.; Meral, C.; Abaci, A. Vaspin and its correlation with insulin sensitivity indices in obese children. Diabetes Res. Clin. Pract. 2009, 84, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Handisurya, A.; Riedl, M.; Vila, G.; Maier, C.; Clodi, M.; Prikoszovich, T.; Ludvik, B.; Prager, G.; Luger, A.; Kautzky-Willer, A. Serum vaspin concentrations in relation to insulin sensitivity following RYGB-induced weight loss. Obes. Surg. 2010, 20, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Park, H.S.; Park, C.Y.; Song, Y.S.; Jang, Y.J. Association between serum vaspin concentrations and visceral adipose tissue in Korean subjects. Metabolism 2010, 59, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Heutling, D.; Chen, J.; Farhatullah, S.; Adya, R.; Keay, S.D.; Kennedy, C.R.; Lehnert, H.; Randeva, H.S. Metformin decreases the adipokine vaspin in overweight women with polycystic ovary syndrome concomitant with improvement in insulin sensitivity and a decrease in insulin resistance. Diabetes 2008, 57, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wen, J.; Lu, Y.; Ji, H.; Zhuang, J.; Su, Y.; Liu, B.; Li, H.; Xu, Y. Impact of age on plasma vaspin concentration in a group of normal chinese people. J. Endocrinol. Investig. 2017, 40, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Sperling, M.; Grzelak, T.; Pelczyńska, M.; Jasinska, P.; Bogdanski, P.; Pupek-Musialik, D.; Czyzewska, K. Concentrations of omentin and vaspin versus insulin resistance in obese individuals. Biomed. Pharmacother. 2016, 83, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.J.; Lee, M.; Park, H.S.; Han, K.; Cho, G.J.; Hwang, T.G.; Kim, J.H.; Lee, S.H.; Lee, H.Y.; Kim, S.M. Elevated vaspin and leptin levels are associated with obesity in prepubertal Korean children. Endocr. J. 2013, 60, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.; Youn, B.S.; Kim, D.W.; Kim, E.H.; Park, J.W.; Namkoong, C.; Jeong, J.Y.; Yoon, S.Y.; Park, J.Y.; Lee, K.U.; et al. Circadian rhythm of serum vaspin in healthy male volunteers: Relation to meals. J. Clin. Endocrinol. Metab. 2010, 95, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, P.; Miehle, K.; Sandner, B.; Stumvoll, M.; Blüher, M. Insulin administration acutely decreases vaspin serum concentrations in humans. Obes. Facts 2013, 6, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Oberbach, A.; Kirsch, K.; Lehmann, S.; Schlichting, N.; Fasshauer, M.; Zarse, K.; Stumvoll, M.; Ristow, M.; Blüher, M.; Kovacs, P. Serum vaspin concentrations are decreased after exercise-induced oxidative stress. Obes. Facts 2010, 3, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.K.; Han, T.K.; Kang, H.S. Combined effects of body mass index and cardio/respiratory fitness on serum vaspin concentrations in korean young men. Eur. J. Appl. Physiol. 2010, 108, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Saboori, S.; Hosseinzadeh-Attar, M.J.; Yousefi Rad, E.; Hosseini, M.; Mirzaei, K.; Ahmadivand, Z. The comparison of serum vaspin and visfatin concentrations in obese and normal weight women. Diabetes Metab. Syndr. 2015, 9, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Dong, Z.; Qian, Y.; Han, Y. Obese type 2 diabetes mellitus patients have higher serum vaspin concentrations. J. Diabetes 2016, 8, 445–447. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Li, Y.; Wang, C.; Luo, C.; Liu, L.; Chuo, F.; Li, Q.; Sun, C. Higher vaspin levels in subjects with obesity and type 2 diabetes mellitus: A meta-analysis. Diabetes. Res. Clin. Pract. 2014, 106, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Vaspin in obesity and diabetes: Pathophysiological and clinical significance. Endocrine 2012, 41, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Martos-Moreno, G.; Kratzsch, J.; Körner, A.; Barrios, V.; Hawkins, F.; Kiess, W.; Argente, J. Serum visfatin and vaspin levels in prepubertal children: Effect of obesity and weight loss after behavior modifications on their secretion and relationship with glucose metabolism. Int. J. Obes. 2011, 35, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Lee, H.J.; Park, H.S.; Kang, J.H.; Kim, K.S.; Song, Y.S.; Jang, Y.J. Effects of weight reduction on serum vaspin concentrations in obese subjects: Modification by insulin resistance. Obesity 2010, 18, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Golpaie, A.; Tajik, N.; Masoudkabir, F.; Karbaschian, Z.; Talebpour, M.; Hoseini, M.; Hosseinzadeh-Attar, M.J. Short-term effect of weight loss through restrictive bariatric surgery on serum levels of vaspin in morbidly obese subjects. Eur. Cytokine Netw. 2011, 22, 181–186. [Google Scholar] [PubMed]
- Wada, J. Vaspin: A novel serpin with insulin-sensitizing effects. Expert Opin. Investig. Drugs 2008, 17, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, L.; Yang, M.; Liu, H.; Liu, D.; Yang, H.; Boden, G.; Yang, G. Short-term continuous subcutaneous insulin infusion decreases the plasma vaspin levels in patients with type 2 diabetes mellitus concomitant with improvement in insulin sensitivity. Eur. J. Endocrinol. 2011, 164, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Kadoglou, N.P.; Gkontopoulos, A.; Kapelouzou, A.; Fotiadis, G.; Theofilogiannakos, E.K.; Kottas, G.; Lampropoulos, S. Serum levels of vaspin and visfatin in patients with coronary artery disease-kozani study. Clin. Chim. Acta 2011, 412, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Jian, W.; Peng, W.; Xiao, S.; Li, H.; Jin, J.; Qin, L.; Dong, Y.; Su, Q. Role of serum vaspin in progression of type 2 diabetes: A 2-year cohort study. PLoS ONE 2014, 9, e94763. [Google Scholar] [CrossRef] [PubMed]
- Tönjes, A.; Fasshauer, M.; Kratzsch, J.; Stumvoll, M.; Blüher, M. Adipokine pattern in subjects with impaired fasting glucose and impaired glucose tolerance in comparison to normal glucose tolerance and diabetes. PLoS ONE 2010, 5, e13911. [Google Scholar] [CrossRef] [PubMed]
- Mm, W.Q.; Fan, J.; Khor, S.; Song, M.; Hong, W.; Dai, X. Serum vaspin levels and vaspin mRNA expression in subcutaneous adipose tissue in women with gestational diabetes mellitus. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 182, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Heiker, J.T.; Klöting, N.; Kovacs, P.; Kuettner, E.B.; Sträter, N.; Schultz, S.; Kern, M.; Stumvoll, M.; Blüher, M.; Beck-Sickinger, A.G. Vaspin inhibits kallikrein 7 by serpin mechanism. Cell. Mol. Life Sci. 2013, 70, 2569–2583. [Google Scholar] [CrossRef] [PubMed]
- Pennica, D.; Shaw, K.J.; Swanson, T.A.; Moore, M.W.; Shelton, D.L.; Zioncheck, K.A.; Rosenthal, A.; Taga, T.; Paoni, N.F.; Wood, W.I. Cardiotrophin-1. Biological activities and binding to the leukemia inhibitory factor receptor/gp130 signaling complex. J. Biol. Chem. 1995, 270, 10915–10922. [Google Scholar] [CrossRef] [PubMed]
- Pennica, D.; Swanson, T.A.; Shaw, K.J.; Kuang, W.J.; Gray, C.L.; Beatty, B.G.; Wood, W.I. Human cardiotrophin-1: Protein and gene structure, biological and binding activities, and chromosomal localization. Cytokine 1996, 8, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Saito, Y.; Miyamoto, Y.; Kuwahara, K.; Ogawa, E.; Nakagawa, O.; Harada, M.; Masuda, I.; Nakao, K. cDNA cloning of rat cardiotrophin-1 (CT-1): Augmented expression of ct-1 gene in ventricle of genetically hypertensive rats. Biochem. Biophys. Res. Commun. 1996, 219, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Pennica, D.; King, K.L.; Shaw, K.J.; Luis, E.; Rullamas, J.; Luoh, S.M.; Darbonne, W.C.; Knutzon, D.S.; Yen, R.; Chien, K.R. Expression cloning of cardiotrophin 1, a cytokine that induces cardiac myocyte hypertrophy. Proc. Natl. Acad. Sci. USA 1995, 92, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S.; Scheller, J.; Schaper, F. “Family reunion” a structured view on the composition of the receptor complexes of interleukin-6-type and interleukin-12-type cytokines. Cytokine Growth Factor. Rev. 2015, 26, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Gadient, R.A.; Patterson, P.H. Leukemia inhibitory factor, interleukin 6, and other cytokines using the gp130 transducing receptor: Roles in inflammation and injury. Stem Cells 1999, 17, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Natal, C.; Fortuño, M.A.; Restituto, P.; Bazán, A.; Colina, I.; Díez, J.; Varo, N. Cardiotrophin-1 is expressed in adipose tissue and upregulated in the metabolic syndrome. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E52–E60. [Google Scholar] [CrossRef] [PubMed]
- López-Yoldi, M.; Marcos-Gomez, B.; Romero-Lozano, M.A.; Sáinz, N.; Prieto, J.; Martínez, J.A.; Bustos, M.; Moreno-Aliaga, M.J. Cardiotrophin-1 regulates adipokine production in 3T3-L1 adipocytes and adipose tissue from obese mice. J. Cell. Physiol. 2017, 232, 2469–2477. [Google Scholar] [CrossRef] [PubMed]
- López-Yoldi, M.; Moreno-Aliaga, M.J.; Bustos, M. Cardiotrophin-1: A multifaceted cytokine. Cytokine Growth Factor Rev. 2015, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- López-Yoldi, M.; Stanhope, K.L.; Garaulet, M.; Chen, X.G.; Marcos-Gómez, B.; Carrasco-Benso, M.P.; Santa Maria, E.M.; Escoté, X.; Lee, V.; Nunez, M.V.; et al. Role of cardiotrophin-1 in the regulation of metabolic circadian rhythms and adipose core clock genes in mice and characterization of 24-h circulating CT-1 profiles in normal-weight and overweight/obese subjects. FASEB J. 2017, 31, 1639–1649. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Madrid, J.A. Chronobiology, genetics and metabolic syndrome. Curr. Opin. Lipidol. 2009, 20, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Van der Spek, R.; Kreier, F.; Fliers, E.; Kalsbeek, A. Circadian rhythms in white adipose tissue. Prog. Brain Res. 2012, 199, 183–201. [Google Scholar] [PubMed]
- Moreno-Aliaga, M.J.; Pérez-Echarri, N.; Marcos-Gómez, B.; Larequi, E.; Gil-Bea, F.J.; Viollet, B.; Gimenez, I.; Martínez, J.A.; Prieto, J.; Bustos, M. Cardiotrophin-1 is a key regulator of glucose and lipid metabolism. Cell. Metab. 2011, 14, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Castaño, D.; Larequi, E.; Belza, I.; Astudillo, A.M.; Martínez-Ansó, E.; Balsinde, J.; Argemi, J.; Aragon, T.; Moreno-Aliaga, M.J.; Muntane, J.; et al. Cardiotrophin-1 eliminates hepatic steatosis in obese mice by mechanisms involving ampk activation. J. Hepatol. 2014, 60, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Matute, P.; Pichel, J.G.; Iñiguez, M.; Recio-Fernández, E.; Pérez-Martínez, L.; Torrens, R.; Blanco, J.R.; Oteo, J.A. Maraviroc ameliorates the increased adipose tissue macrophage recruitment induced by a high-fat diet in a mouse model of obesity. Antivir. Ther. 2017, 22, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Infantes, D.; White, U.A.; Elks, C.M.; Morrison, R.F.; Gimble, J.M.; Considine, R.V.; Ferrante, A.W.; Ravussin, E.; Stephens, J.M. Oncostatin M is produced in adipose tissue and is regulated in conditions of obesity and type 2 diabetes. J. Clin. Endocrinol. Metab. 2014, 99, E217–E225. [Google Scholar] [CrossRef] [PubMed]
- Malavazos, A.E.; Ermetici, F.; Morricone, L.; Delnevo, A.; Coman, C.; Ambrosi, B.; Corsi, M.M. Association of increased plasma cardiotrophin-1 with left ventricular mass indexes in normotensive morbid obesity. Hypertension 2008, 51, e8. [Google Scholar] [CrossRef] [PubMed]
- Rendo-Urteaga, T.; García-Calzón, S.; Martínez-Ansó, E.; Chueca, M.; Oyarzabal, M.; Azcona-Sanjulián, M.C.; Bustos, M.; Moreno-Aliaga, M.J.; Martínez, J.A.; Marti, A. Decreased cardiotrophin-1 levels are associated with a lower risk of developing the metabolic syndrome in overweight/obese children after a weight loss program. Metabolism 2013, 62, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.C.; Lu, F.H.; Wu, H.T.; Ou, H.Y.; Yang, Y.C.; Wu, J.S.; Chang, C.J. Cardiotrophin-1 is inversely associated with obesity in non-diabetic individuals. Sci. Rep. 2015, 5, 17438. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Fritzenwanger, M.; Figulla, H.R. Cardiotrophin-1 in adolescents: Impact of obesity and blood pressure. Hypertension 2008, 52, e6. [Google Scholar] [CrossRef] [PubMed]
- Schroers, A.; Hecht, O.; Kallen, K.J.; Pachta, M.; Rose-John, S.; Grötzinger, J. Dynamics of the gp130 cytokine complex: A model for assembly on the cellular membrane. Protein Sci. 2005, 14, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Robledo, O.; Fourcin, M.; Chevalier, S.; Guillet, C.; Auguste, P.; Pouplard-Barthelaix, A.; Pennica, D.; Gascan, H. Signaling of the cardiotrophin-1 receptor. Evidence for a third receptor component. J. Biol. Chem. 1997, 272, 4855–4863. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.G.; Pennica, D.; Patterson, P.H. Cardiotrophin-1 induces the same neuropeptides in sympathetic neurons as do neuropoietic cytokines. J. Neurochem. 1997, 69, 2278–2284. [Google Scholar] [CrossRef] [PubMed]
- Zvonic, S.; Hogan, J.C.; Arbour-Reily, P.; Mynatt, R.L.; Stephens, J.M. Effects of cardiotrophin on adipocytes. J. Biol. Chem. 2004, 279, 47572–47579. [Google Scholar] [CrossRef] [PubMed]
- López-Yoldi, M.; Fernández-Galilea, M.; Laiglesia, L.M.; Larequi, E.; Prieto, J.; Martínez, J.A.; Bustos, M.; Moreno-Aliaga, M.J. Cardiotrophin-1 stimulates lipolysis through the regulation of main adipose tissue lipases. J. Lipid Res. 2014, 55, 2634–2643. [Google Scholar] [CrossRef] [PubMed]
- Romacho, T.; Elsen, M.; Röhrborn, D.; Eckel, J. Adipose tissue and its role in organ crosstalk. Acta Physiol. 2014, 210, 733–753. [Google Scholar] [CrossRef] [PubMed]
- López-Yoldi, M.; Castilla-Madrigal, R.; Lostao, M.P.; Barber, A.; Prieto, J.; Martínez, J.A.; Bustos, M.; Moreno-Aliaga, M.J. Cardiotrophin-1 decreases intestinal sugar uptake in mice and in caco-2 cells. Acta Physiol. 2016, 217, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Chao, E.C. Sglt-2 inhibitors: A new mechanism for glycemic control. Clin. Diabetes 2014, 32, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Chicheportiche, Y.; Bourdon, P.R.; Xu, H.; Hsu, Y.M.; Scott, H.; Hession, C.; Garcia, I.; Browning, J.L. Tweak, a new secreted ligand in the tumor necrosis factor family that weakly induces apoptosis. J. Biol. Chem. 1997, 272, 32401–32410. [Google Scholar] [CrossRef] [PubMed]
- Tiller, G.; Laumen, H.; Fischer-Posovszky, P.; Finck, A.; Skurk, T.; Keuper, M.; Brinkmann, U.; Wabitsch, M.; Link, D.; Hauner, H. Light (TNFSF14) inhibits adipose differentiation without affecting adipocyte metabolism. Int. J. Obes. 2011, 35, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Tajrishi, M.M.; Zheng, T.S.; Burkly, L.C.; Kumar, A. The TWEAK-Fn14 pathway: A potent regulator of skeletal muscle biology in health and disease. Cytokine Growth Factor Rev. 2014, 25, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Burkly, L.C. Regulation of tissue responses: The TWEAK/Fn14 pathway and other TNF/TNFR superfamily members that activate non-canonical nfκb signaling. Front. Immunol. 2015, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Ishidoh, K.; Kayagaki, N.; Kojima, Y.; Yamaguchi, N.; Nakano, H.; Kominami, E.; Okumura, K.; Yagita, H. Multiple pathways of tweak-induced cell death. J. Immunol. 2002, 168, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Burkly, L.C.; Michaelson, J.S.; Zheng, T.S. TWEAK/Fn pathway: An immunological switch for shaping tissue responses. Immunol. Rev. 2011, 244, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Kawakita, T.; Shiraki, K.; Yamanaka, Y.; Yamaguchi, Y.; Saitou, Y.; Enokimura, N.; Yamamoto, N.; Okano, H.; Sugimoto, K.; Murata, K.; et al. Functional expression of tweak in human hepatocellular carcinoma: Possible implication in cell proliferation and tumor angiogenesis. Biochem. Biophys. Res. Commun. 2004, 318, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.L.; McDonough, W.S.; Donohue, P.J.; Winkles, J.A.; Berens, T.J.; Ross, K.R.; Hoelzinger, D.B.; Beaudry, C.; Coons, S.W.; Berens, M.E. The human fn14 receptor gene is up-regulated in migrating glioma cells in vitro and overexpressed in advanced glial tumors. Am. J. Pathol. 2003, 162, 1313–1321. [Google Scholar] [CrossRef]
- Nakayama, M.; Harada, N.; Okumura, K.; Yagita, H. Characterization of murine tweak and its receptor (fn14) by monoclonal antibodies. Biochem. Biophys. Res. Commun. 2003, 306, 819–825. [Google Scholar] [CrossRef]
- Chacón, M.R.; Richart, C.; Gómez, J.M.; Megía, A.; Vilarrasa, N.; Fernández-Real, J.M.; García-España, A.; Miranda, M.; Masdevall, C.; Ricard, W.; et al. Expression of tweak and its receptor fn14 in human subcutaneous adipose tissue. Relationship with other inflammatory cytokines in obesity. Cytokine 2006, 33, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Girgenrath, M.; Weng, S.; Kostek, C.A.; Browning, B.; Wang, M.; Brown, S.A.; Winkles, J.A.; Michaelson, J.S.; Allaire, N.; Schneider, P.; et al. Tweak, via its receptor fn14, is a novel regulator of mesenchymal progenitor cells and skeletal muscle regeneration. EMBO J. 2006, 25, 5826–5839. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, A.; Ambrose, C.; Parr, M.; Lincecum, J.M.; Wang, M.Z.; Zheng, T.S.; Browning, B.; Michaelson, J.S.; Baetscher, M.; Baestcher, M.; et al. Tweak induces liver progenitor cell proliferation. J. Clin. Investig. 2005, 115, 2330–2340. [Google Scholar] [CrossRef] [PubMed]
- Novoyatleva, T.; Schymura, Y.; Janssen, W.; Strobl, F.; Swiercz, J.M.; Patra, C.; Posern, G.; Wietelmann, A.; Zheng, T.S.; Schermuly, R.T.; et al. Deletion of Fn14 receptor protects from right heart fibrosis and dysfunction. Basic Res. Cardiol. 2013, 108, 325. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Bartolomé, J.; Llauradó, G.; Rodriguez, M.M.; Fernandez-Real, J.M.; Garcia-Fontgivell, J.F.; Puig, J.; Maymó-Masip, E.; Vendrell, J.; Chacón, M.R. Reduced circulating levels of stweak are associated with nafld and may affect hepatocyte triglyceride accumulation. Int. J. Obes. 2016, 40, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Tirnitz-Parker, J.E.; Viebahn, C.S.; Jakubowski, A.; Klopcic, B.R.; Olynyk, J.K.; Yeoh, G.C.; Knight, B. Tumor necrosis factor-like weak inducer of apoptosis is a mitogen for liver progenitor cells. Hepatology 2010, 52, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Mustonen, E.; Säkkinen, H.; Tokola, H.; Isopoussu, E.; Aro, J.; Leskinen, H.; Ruskoaho, H.; Rysä, J. Tumour necrosis factor-like weak inducer of apoptosis (TWEAK) and its receptor fn14 during cardiac remodelling in rats. Acta Physiol. 2010, 199, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Dohi, T.; Borodovsky, A.; Wu, P.; Shearstone, J.R.; Kawashima, R.; Runkel, L.; Rajman, L.; Dong, X.; Scott, M.L.; Michaelson, J.S.; et al. Tweak/fn14 pathway: A nonredundant role in intestinal damage in mice through a tweak/intestinal epithelial cell axis. Gastroenterology 2009, 136, 912–923. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Bhatnagar, S.; Kumar, A.; Lach-Trifilieff, E.; Wauters, S.; Li, H.; Makonchuk, D.Y.; Glass, D.J. The TWEAK/Fn4 system is a critical regulator of denervation-induced skeletal muscle atrophy in mice. J. Cell. Biol. 2010, 188, 833–849. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-García, B.; Martín-Ventura, J.L.; Martínez, E.; Sánchez, S.; Hernández, G.; Ortega, L.; Ortiz, A.; Egido, J.; Blanco-Colio, L.M. Fn14 is upregulated in cytokine-stimulated vascular smooth muscle cells and is expressed in human carotid atherosclerotic plaques: Modulation by atorvastatin. Stroke 2006, 37, 2044–2053. [Google Scholar] [CrossRef] [PubMed]
- Justo, P.; Sanz, A.B.; Sanchez-Niño, M.D.; Winkles, J.A.; Lorz, C.; Egido, J.; Ortiz, A. Cytokine cooperation in renal tubular cell injury: The role of tweak. Kidney Int. 2006, 70, 1750–1758. [Google Scholar] [CrossRef] [PubMed]
- Vendrell, J.; Maymó-Masip, E.; Tinahones, F.; García-España, A.; Megia, A.; Caubet, E.; García-Fuentes, E.; Chacón, M.R. Tumor necrosis-like weak inducer of apoptosis as a proinflammatory cytokine in human adipocyte cells: Up-regulation in severe obesity is mediated by inflammation but not hypoxia. J. Clin. Endocrinol. Metab. 2010, 95, 2983–2992. [Google Scholar] [CrossRef] [PubMed]
- Llauradó, G.; González-Clemente, J.M.; Maymó-Masip, E.; Subías, D.; Vendrell, J.; Chacón, M.R. Serum levels of TWEAK and scavenger receptor CD163 in type 1 diabetes mellitus: Relationship with cardiovascular risk factors. A case-control study. PLoS ONE 2012, 7, e43919. [Google Scholar] [CrossRef] [PubMed]
- Maymó-Masip, E.; Fernández-Veledo, S.; Garcia España, A.; Vázquez-Carballo, A.; Tinahones, F.J.; García-Fuentes, E.; Garrifo-Sanchez, L.; Rodriguez, M.E.M.; Vendrell, J.; Chacón, M.R. The rise of soluble TWEAK levels in severely obese subjects after bariatric surgery may affect adipocyte-cytokine production induced by TNFα. J. Clin. Endocrinol. Metab. 2013, 98, E1323–E1333. [Google Scholar] [CrossRef] [PubMed]
- Maecker, H.; Varfolomeev, E.; Kischkel, F.; Lawrence, D.; LeBlanc, H.; Lee, W.; Hurst, S.; Danilenko, D.; Li, J.; Filvaroff, E.; et al. Tweak attenuates the transition from innate to adaptive immunity. Cell 2005, 123, 931–944. [Google Scholar] [CrossRef] [PubMed]
- Alexaki, V.I.; Notas, G.; Pelekanou, V.; Kampa, M.; Valkanou, M.; Theodoropoulos, P.; Stathopoulos, E.N.; Tsapis, A.; Castanas, E. Adipocytes as immune cells: Differential expression of tweak, baff, and april and their receptors (Fn14, BAFF-R, TACI, and BCMA) at different stages of normal and pathological adipose tissue development. J. Immunol. 2009, 183, 5948–5956. [Google Scholar] [CrossRef] [PubMed]
- Tiller, G.; Fischer-Posovszky, P.; Laumen, H.; Finck, A.; Skurk, T.; Keuper, M.; Brinkmann, U.; Wabitsch, M.; Link, D.; Hauner, H. Effects of tweak (TNF superfamily member 12) on differentiation, metabolism, and secretory function of human primary preadipocytes and adipocytes. Endocrinology 2009, 150, 5373–5383. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.; Strissel, K.J.; DeFuria, J.; Wang, J.; Wu, D.; Burkly, L.C.; Obin, M.S. Deletion of TNF-like weak inducer of apoptosis (tweak) protects mice from adipose and systemic impacts of severe obesity. Obesity 2014, 22, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Kralisch, S.; Ziegelmeier, M.; Bachmann, A.; Seeger, J.; Lössner, U.; Blüher, M.; Stumvoll, M.; Fasshauer, M. Serum levels of the atherosclerosis biomarker stweak are decreased in type 2 diabetes and end-stage renal disease. Atherosclerosis 2008, 199, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.I.; Sonmez, A.; Ortiz, A.; Saglam, M.; Kilic, S.; Eyileten, T.; Caglar, K.; Oguz, Y.; Vural, A.; Çakar, M.; et al. Soluble tweak and ptx3 in nondialysis ckd patients: Impact on endothelial dysfunction and cardiovascular outcomes. Clin. J. Am. Soc. Nephrol. 2011, 6, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Carrero, J.J.; Ortiz, A.; Qureshi, A.R.; Martín-Ventura, J.L.; Bárány, P.; Heimbürger, O.; Marrón, B.; Metry, G.; Snaedal, S.; Lindholm, B.; et al. Additive effects of soluble tweak and inflammation on mortality in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2009, 4, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.I.; Carrero, J.J.; Ortiz, A.; Martín-Ventura, J.L.; Sonmez, A.; Saglam, M.; Yaman, H.; Yenicesu, M.; Egido, J.; Blanco-Colio, L.M. Soluble tweak plasma levels as a novel biomarker of endothelial function in patients with chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 1716–1723. [Google Scholar] [CrossRef] [PubMed]
- López-Dupla, M.; Maymó-Masip, E.; Martínez, E.; Domingo, P.; Leal, M.; Peraire, J.; Viladés, C.; Veloso, S.; Arnedo, M.; Ferrando-Martínez, S.; et al. HIV-1/HAART-related lipodystrophy syndrome (HALS) is associated with decreased circulating sTWEAK levels. PLoS ONE 2015, 10, e0144789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Ogura, Y.; Tajrishi, M.M.; Kumar, A. Elevated levels of tweak in skeletal muscle promote visceral obesity, insulin resistance, and metabolic dysfunction. FASEB J. 2015, 29, 988–1002. [Google Scholar] [CrossRef] [PubMed]
- Wiley, S.R.; Cassiano, L.; Lofton, T.; Davis-Smith, T.; Winkles, J.A.; Lindner, V.; Liu, H.; Daniel, T.O.; Smith, C.A.; Fanslow, W.C. A novel tnf receptor family member binds tweak and is implicated in angiogenesis. Immunity 2001, 15, 837–846. [Google Scholar] [CrossRef]
- Feng, S.L.; Guo, Y.; Factor, V.M.; Thorgeirsson, S.S.; Bell, D.W.; Testa, J.R.; Peifley, K.A.; Winkles, J.A. The fn14 immediate-early response gene is induced during liver regeneration and highly expressed in both human and murine hepatocellular carcinomas. Am. J. Pathol. 2000, 156, 1253–1261. [Google Scholar] [CrossRef]
- Brown, S.A.; Hanscom, H.N.; Vu, H.; Brew, S.A.; Winkles, J.A. Tweak binding to the fn14 cysteine-rich domain depends on charged residues located in both the a1 and d2 modules. Biochem. J. 2006, 397, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Bover, L.C.; Cardó-Vila, M.; Kuniyasu, A.; Sun, J.; Rangel, R.; Takeya, M.; Aggarwal, B.B.; Arap, W.; Pasqualini, R. A previously unrecognized protein-protein interaction between tweak and cd163: Potential biological implications. J. Immunol. 2007, 178, 8183–8194. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.A.; Muñoz-García, B.; Martín-Ventura, J.L.; Madrigal-Matute, J.; Orbe, J.; Páramo, J.A.; Ortega, L.; Egido, J.; Blanco-Colio, L.M. The cd163-expressing macrophages recognize and internalize tweak: Potential consequences in atherosclerosis. Atherosclerosis 2009, 207, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Polek, T.C.; Talpaz, M.; Darnay, B.G.; Spivak-Kroizman, T. Tweak mediates signal transduction and differentiation of raw264.7 cells in the absence of fn14/tweakr. Evidence for a second tweak receptor. J. Biol. Chem. 2003, 278, 32317–32323. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.A.; Richards, C.M.; Hanscom, H.N.; Feng, S.L.; Winkles, J.A. The fn14 cytoplasmic tail binds tumour-necrosis-factor-receptor-associated factors 1, 2, 3 and 5 and mediates nuclear factor-kappab activation. Biochem. J. 2003, 371, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Potrovita, I.; Zhang, W.; Burkly, L.; Hahm, K.; Lincecum, J.; Wang, M.Z.; Maurer, M.H.; Rossner, M.; Schneider, A.; Schwaninger, M. Tumor necrosis factor-like weak inducer of apoptosis-induced neurodegeneration. J. Neurosci. 2004, 24, 8237–8244. [Google Scholar] [CrossRef] [PubMed]
- Chicheportiche, Y.; Chicheportiche, R.; Sizing, I.; Thompson, J.; Benjamin, C.B.; Ambrose, C.; Dayer, J.M. Proinflammatory activity of tweak on human dermal fibroblasts and synoviocytes: Blocking and enhancing effects of anti-tweak monoclonal antibodies. Arthritis Res. 2002, 4, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Saas, P.; Boucraut, J.; Walker, P.R.; Quiquerez, A.L.; Billot, M.; Desplat-Jego, S.; Chicheportiche, Y.; Dietrich, P.Y. Tweak stimulation of astrocytes and the proinflammatory consequences. Glia 2000, 32, 102–107. [Google Scholar] [CrossRef]
- Xu, H.; Okamoto, A.; Ichikawa, J.; Ando, T.; Tasaka, K.; Masuyama, K.; Ogawa, H.; Yagita, H.; Okumura, K.; Nakao, A. TWEAK/Fn14 interaction stimulates human bronchial epithelial cells to produce IL-8 and GM-CSF. Biochem. Biophys. Res. Commun. 2004, 318, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Donohue, P.J.; Richards, C.M.; Brown, S.A.; Hanscom, H.N.; Buschman, J.; Thangada, S.; Hla, T.; Williams, M.S.; Winkles, J.A. TWEAK is an endothelial cell growth and chemotactic factor that also potentiates FGF-2 and VEGF-A mitogenic activity. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Enwere, E.K.; Holbrook, J.; Lejmi-Mrad, R.; Vineham, J.; Timusk, K.; Sivaraj, B.; Isaac, M.; Uehling, D.; Al-awar, R.; LaCasse, E.; et al. Tweak and cIAP1 regulate myoblast fusion through the noncanonical NF-κB signaling pathway. Sci. Signal. 2012, 5, ra75. [Google Scholar] [CrossRef] [PubMed]
- Roos, C.; Wicovsky, A.; Müller, N.; Salzmann, S.; Rosenthal, T.; Kalthoff, H.; Trauzold, A.; Seher, A.; Henkler, F.; Kneitz, C.; et al. Soluble and transmembrane tnf-like weak inducer of apoptosis differentially activate the classical and noncanonical NF-κB pathway. J. Immunol. 2010, 185, 1593–1605. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Carballo, A.; Ceperuelo-Mallafré, V.; Chacón, M.R.; Maymó-Masip, E.; Lorenzo, M.; Porras, A.; Vendrell, J.; Fernández-Veledo, S. Tweak prevents TNF-α-induced insulin resistance through pp2a activation in human adipocytes. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E101–E112. [Google Scholar] [CrossRef] [PubMed]
- Bork, P. The modular architecture of a new family of growth regulators related to connective tissue growth factor. FEBS Lett. 1993, 327, 125–130. [Google Scholar] [CrossRef]
- Perbal, B. The CCN3 (NOV) cell growth regulator: A new tool for molecular medicine. Expert Rev. Mol. Diagn. 2003, 3, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Burren, C.P.; Wanek, D.; Mohan, S.; Cohen, P.; Guevara-Aguirre, J.; Rosenfeld, R.G. Serum levels of insulin-like growth factor binding proteins in ecuadorean children with growth hormone insensitivity. Acta Paediatr. 1999, 88, S185–S191. [Google Scholar] [CrossRef]
- Burren, C.P.; Wilson, E.M.; Hwa, V.; Oh, Y.; Rosenfeld, R.G. Binding properties and distribution of insulin-like growth factor binding protein-related protein 3 (IGFBP-rP3/NovH), an additional member of the igfbp superfamily. J. Clin. Endocrinol. Metab. 1999, 84, 1096–1103. [Google Scholar] [PubMed]
- Martinerie, C.; Gicquel, C.; Louvel, A.; Laurent, M.; Schofield, P.N.; Le Bouc, Y. Altered expression of novH is associated with human adrenocortical tumorigenesis. J. Clin. Endocrinol. Metab. 2001, 86, 3929–3940. [Google Scholar] [CrossRef] [PubMed]
- Pakradouni, J.; Le Goff, W.; Calmel, C.; Antoine, B.; Villard, E.; Frisdal, E.; Abifadel, M.; Tordjman, J.; Poitou, C.; Bonnefont-Rousselot, D.; et al. Plasma NOV/CCN3 levels are closely associated with obesity in patients with metabolic disorders. PLoS ONE 2013, 8, e66788. [Google Scholar] [CrossRef] [PubMed]
- Thibout, H.; Martinerie, C.; Créminon, C.; Godeau, F.; Boudou, P.; Le Bouc, Y.; Laurent, M. Characterization of human nov in biological fluids: An enzyme immunoassay for the quantification of human nov in sera from patients with diseases of the adrenal gland and of the nervous system. J. Clin. Endocrinol. Metab. 2003, 88, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Perbal, B. Pathogenic potential of myeloblastosis-associated viruses. Infect. Agents Dis. 1995, 4, 212–227. [Google Scholar] [PubMed]
- Perbal, B. Contribution of mav-1-induced nephroblastoma to the study of genes involved in human wilms’ tumor development. Crit. Rev. Oncog. 1994, 5, 589–613. [Google Scholar] [PubMed]
- Gellhaus, A.; Dong, X.; Propson, S.; Maass, K.; Klein-Hitpass, L.; Kibschull, M.; Traub, O.; Willecke, K.; Perbal, B.; Lye, S.J.; et al. Connexin43 interacts with nov: A possible mechanism for negative regulation of cell growth in choriocarcinoma cells. J. Biol. Chem. 2004, 279, 36931–36942. [Google Scholar] [CrossRef] [PubMed]
- Glukhova, L.; Angevin, E.; Lavialle, C.; Cadot, B.; Terrier-Lacombe, M.J.; Perbal, B.; Bernheim, A.; Goguel, A.F. Patterns of specific genomic alterations associated with poor prognosis in high-grade renal cell carcinomas. Cancer Genet. Cytogenet. 2001, 130, 105–110. [Google Scholar] [CrossRef]
- Maillard, M.; Cadot, B.; Ball, R.Y.; Sethia, K.; Edwards, D.R.; Perbal, B.; Tatoud, R. Differential expression of the ccn3 (nov) proto-oncogene in human prostate cell lines and tissues. Mol. Pathol. 2001, 54, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Manara, M.C.; Perbal, B.; Benini, S.; Strammiello, R.; Cerisano, V.; Perdichizzi, S.; Serra, M.; Astolfi, A.; Bertoni, F.; Alami, J.; et al. The expression of CCN3(nov) gene in musculoskeletal tumors. Am. J. Pathol. 2002, 160, 849–859. [Google Scholar] [CrossRef]
- Yu, C.; Le, A.T.; Yeger, H.; Perbal, B.; Alman, B.A. Nov (ccn3) regulation in the growth plate and ccn family member expression in cartilage neoplasia. J. Pathol. 2003, 201, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Lau, L.F. Functions and mechanisms of action of ccn matricellular proteins. Int. J. Biochem. Cell. Biol. 2009, 41, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Kular, L.; Pakradouni, J.; Kitabgi, P.; Laurent, M.; Martinerie, C. The ccn family: A new class of inflammation modulators? Biochimie 2011, 93, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Martinerie, C.; Garcia, M.; Huong Do, T.T.; Antoine, B.; Moldes, M.; Dorothee, G.; Kazazian, C.; Auclair, M.; Buyse, M.; Ledent, T.; et al. NOV/CCN3: A new adipocytokine involved in obesity-associated insulin resistance. Diabetes 2016, 65, 2502–2515. [Google Scholar] [CrossRef] [PubMed]
- Paradis, R.; Lazar, N.; Antinozzi, P.; Perbal, B.; Buteau, J. Nov/ccn3, a novel transcriptional target of foxo1, impairs pancreatic β-cell function. PLoS ONE 2013, 8, e64957. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, T.; Hiraoka, S.; Takemoto, M.; Koshizaka, M.; Tokuyama, H.; Tokuyama, T.; Watanabe, A.; Fujimoto, M.; Kawamura, H.; Sato, S.; et al. Ccn3 inhibits neointimal hyperplasia through modulation of smooth muscle cell growth and migration. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 675–682. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escoté, X.; Gómez-Zorita, S.; López-Yoldi, M.; Milton-Laskibar, I.; Fernández-Quintela, A.; Martínez, J.A.; Moreno-Aliaga, M.J.; Portillo, M.P. Role of Omentin, Vaspin, Cardiotrophin-1, TWEAK and NOV/CCN3 in Obesity and Diabetes Development. Int. J. Mol. Sci. 2017, 18, 1770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18081770
Escoté X, Gómez-Zorita S, López-Yoldi M, Milton-Laskibar I, Fernández-Quintela A, Martínez JA, Moreno-Aliaga MJ, Portillo MP. Role of Omentin, Vaspin, Cardiotrophin-1, TWEAK and NOV/CCN3 in Obesity and Diabetes Development. International Journal of Molecular Sciences. 2017; 18(8):1770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18081770
Chicago/Turabian StyleEscoté, Xavier, Saioa Gómez-Zorita, Miguel López-Yoldi, Iñaki Milton-Laskibar, Alfredo Fernández-Quintela, J. Alfredo Martínez, María J. Moreno-Aliaga, and María P. Portillo. 2017. "Role of Omentin, Vaspin, Cardiotrophin-1, TWEAK and NOV/CCN3 in Obesity and Diabetes Development" International Journal of Molecular Sciences 18, no. 8: 1770. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18081770