Effects of Red Palm Oil on Myocardial Antioxidant Enzymes, Nitric Oxide Synthase and Heart Function in Spontaneously Hypertensive Rats

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Main Characteristics of Experimental Rats
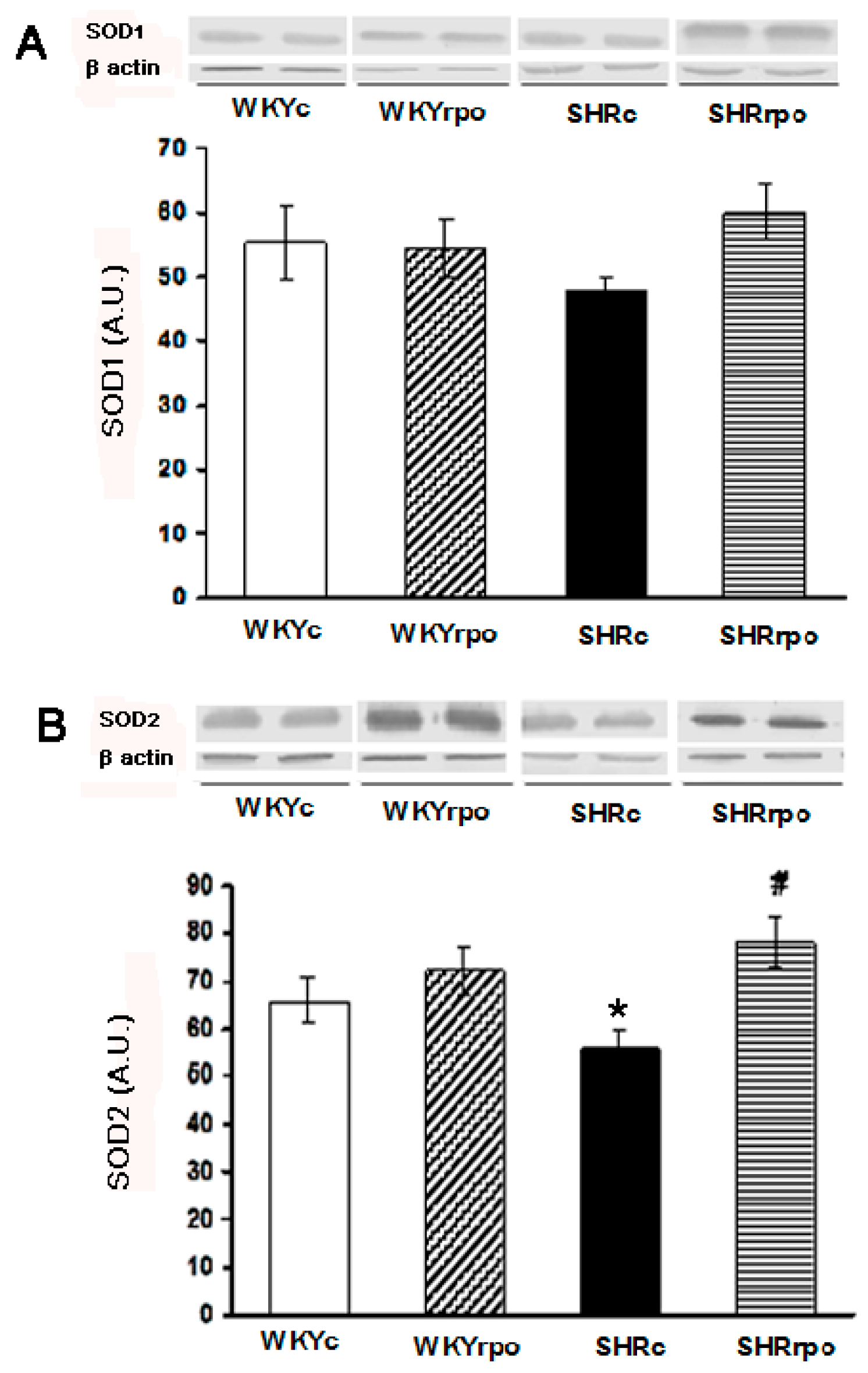
2.2. Effect of Dietary Red Palm Oil (RPO) on the Myocardial SOD1 and SOD2 Protein Expression in Normotensive and Hypertensive Rats
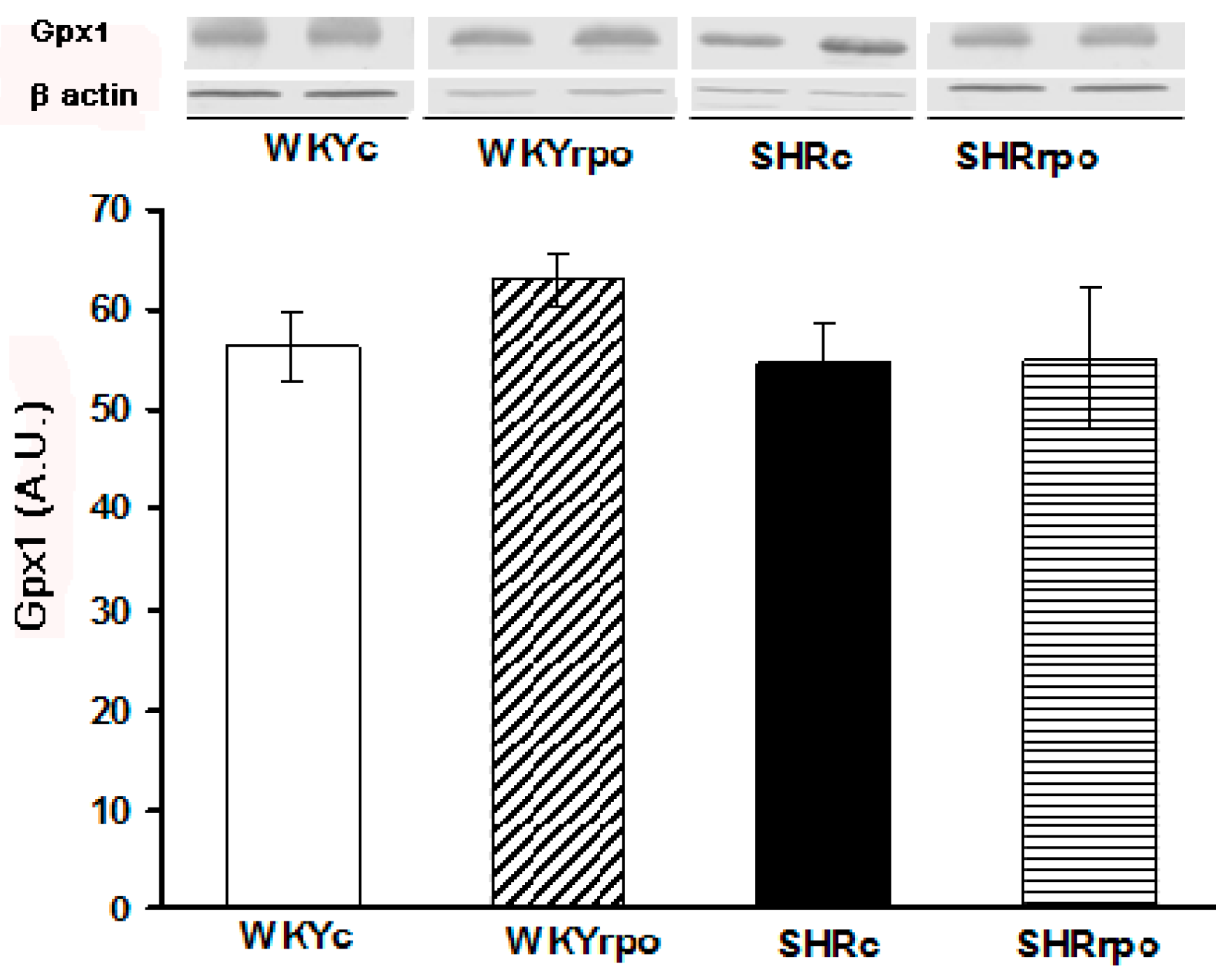
2.3. Effect of Dietary RPO on the Myocardial Gpx1 Protein Expression in Normotensive and Hypertensive Rats
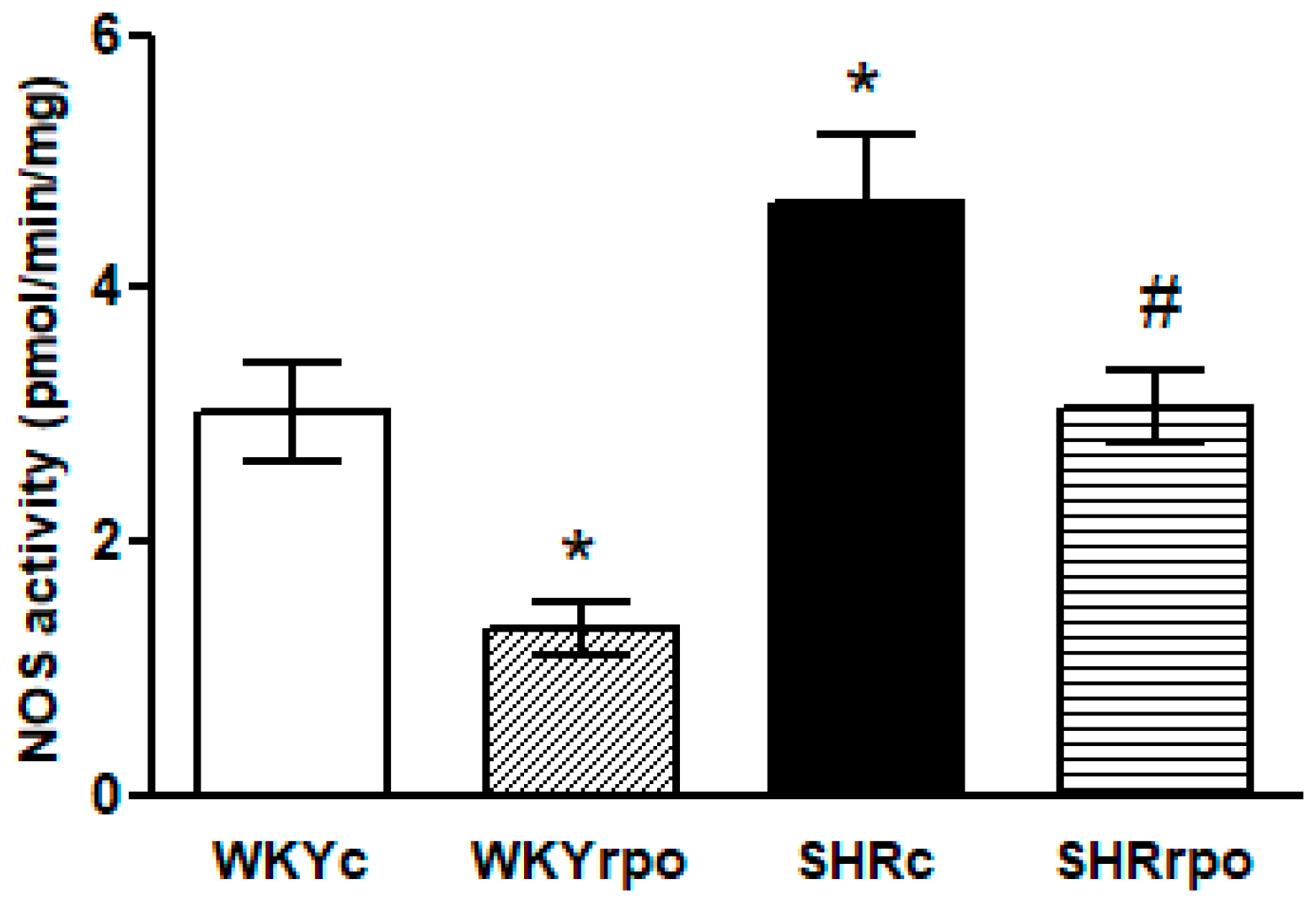
2.4. Effect of Dietary RPO Supplementation on the Nitric Oxide Synthase (NOS) Activity
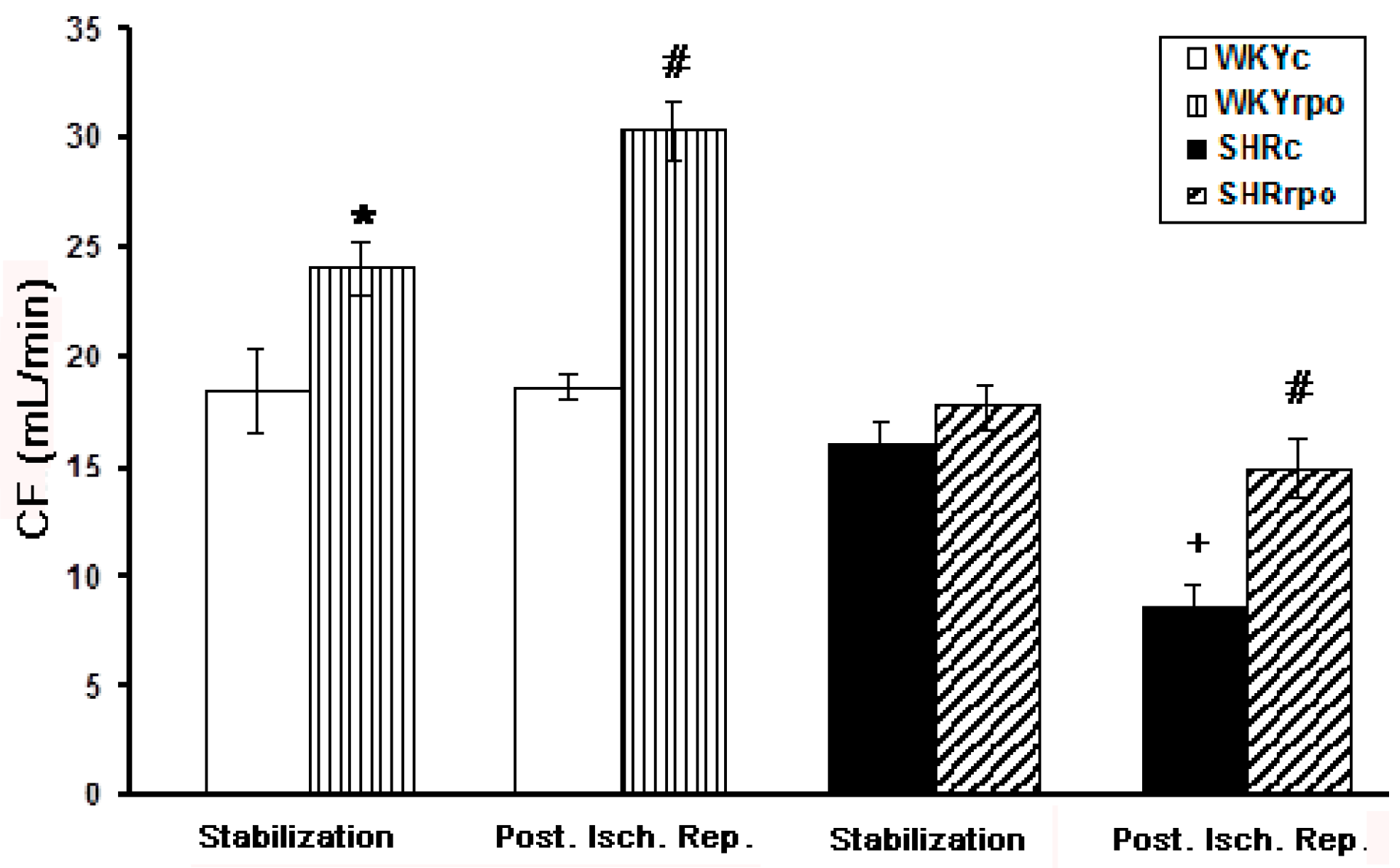
2.5. Effect of RPO Supplementation on the Coronary Flow of Langendorff-Perfused Heart of Normotensive and Hypertensive Rats during Basal Condition and at Early-Phase of Post-Ischemic Reperfusion
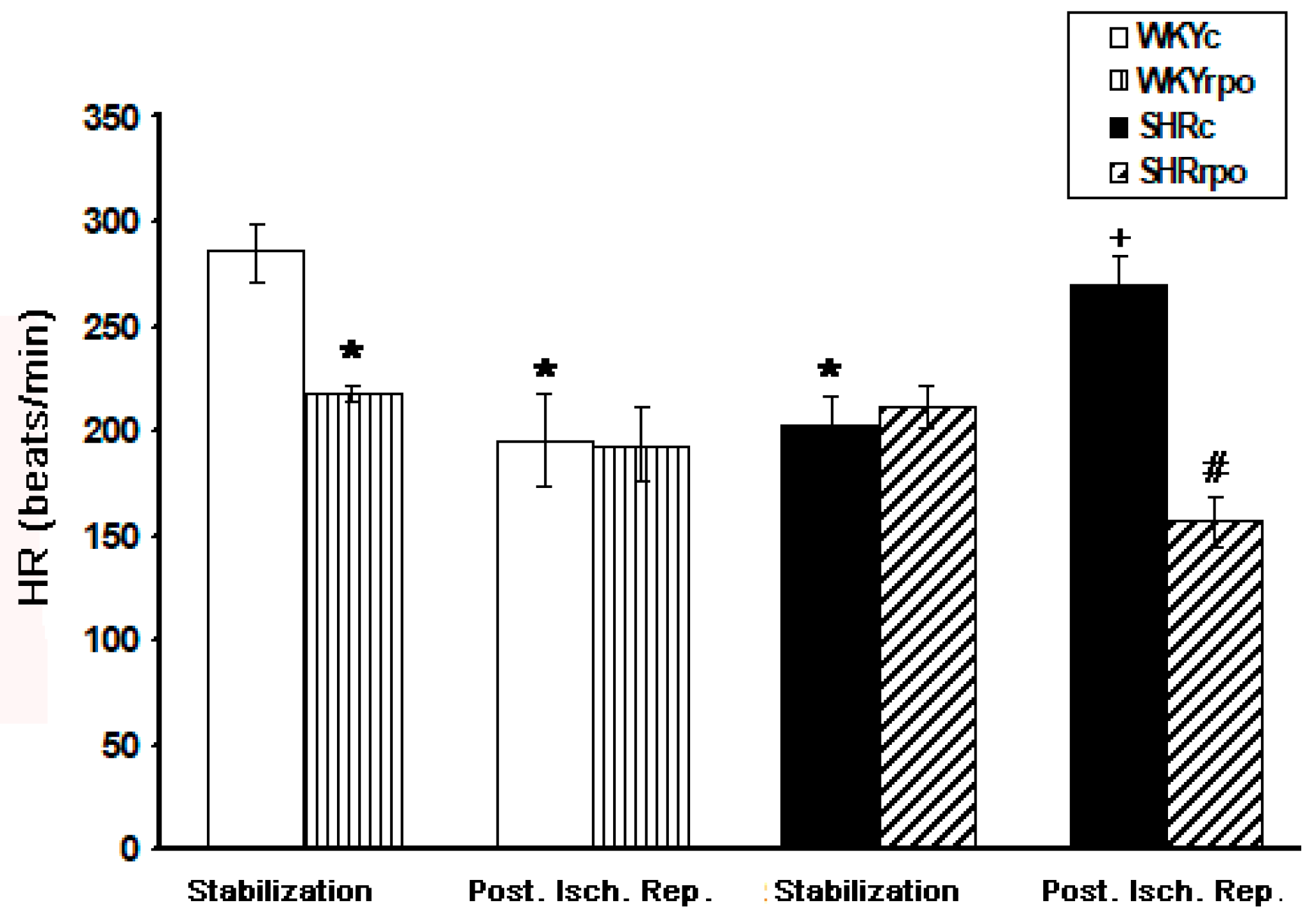
2.6. Effect of RPO Supplementation on the Heart Rate of Langendorff-Perfused Heart of Normotensive and Hypertensive Rats during Basal Condition and at Early-Phase of Post-Ischemic Reperfusion
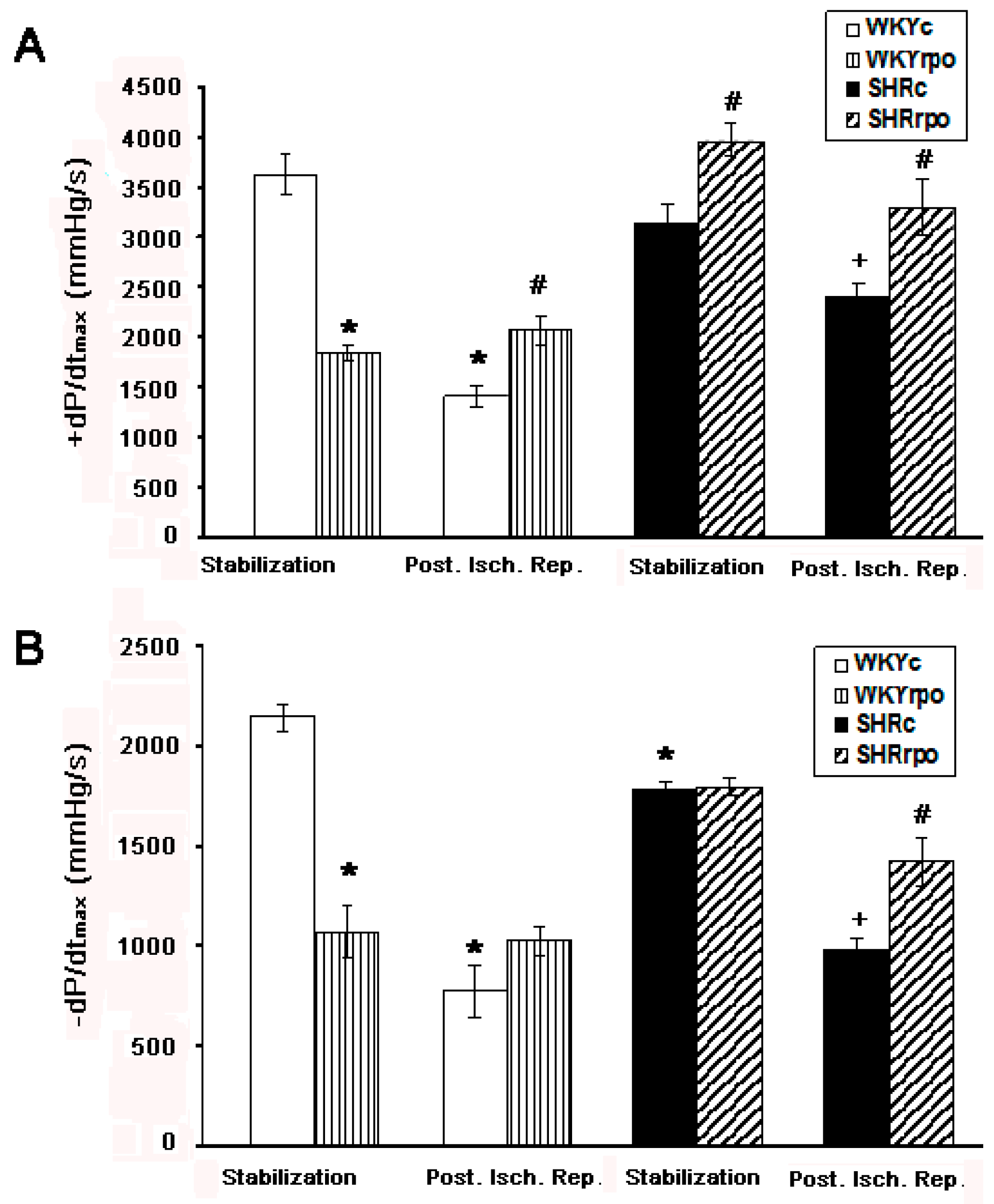
2.7. Contraction and Relaxation Function Assessed by +dP/dtmax and −dP/dtmax in Langendorff-Perfused Heart of WKY and SHR during Basal Condition and at Early-Phase of Post-Ischemic Reperfusion
3. Discussion
4. Materials and Methods
4.1. Animal Care and Ethical Considerations
4.2. Animal Monitoring and Tissue Sampling
4.3. Experimental Protocol of Isolated Langendorff-Perfused Heart
4.4. Measurement of NOS Activity
4.5. Analysis of Myocardial SOD1, SOD2 and Gpx1 Expression Using Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carretero, O.A.; Oparil, S. Essential Hypertension. Circulation 2000, 101, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Egan Benova, T.; Szeiffova Bacova, B.; Viczenczova, C.; Barancik, M.; Tribulova, N. Myocardial Connexin-43 is Implicated in the Prevention of Malignant Arrhythmia in Rats Suffering from Essential Hypertension. In Update on Essential Hypertension; Salazar-Sanchez, L., Ed.; InTech: Rijeka, Croatia, 2016; ISBN 978-953-51-2615-7. Available online: https://www.intechopen.com/books/update-on-essential-hypertension/myocardial-connexin-43-is-implicated-in-the-prevention-of-malignant-arrhythmia-in-rats-suffering-fro (accessed on 14 September 2016). [CrossRef]
- Mein, C.A.; Caulfield, M.J.; Dobson, R.J.; Munroe, P.B. Genetics of essential hypertension. Hum. Mol. Genet. 2004, 13, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.; Maresso, K.; Broeckel, U. Genetic determinants of hypertension: An update. Curr. Hypertens. Rep. 2008, 10, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Abate, N.I.; Mansour, Y.H.; Tuncel, M.; Arbique, D.; Chavoshan, B.; Kizilbash, A.; Howell-Stampley, T.; Vongpatanasin, W.; Victor, R.G. Overweight and Sympathetic Overactivity in Black Americans. Hypertension 2001, 38, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Heineke, J.; Molkentin, J.D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nat. Rev. Mol. Cell Biol. 2006, 7, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Egan Benova, T.; Szeiffova Bacova, B.; Viczenczova, C.; Diez, E.; Barancik, M.; Tribulova, N. Protection of cardiac cell-to-cell coupling attenuate myocardial remodeling and proarrhythmia induced by hypertension. Physiol. Res. 2016, 65, 29–42. [Google Scholar]
- Pokharel, S.; Sharma, U.C.; Pinto, Y.M. Left ventricular hypertrophy: Virtuous intentions, malign consequences. Int. J. Biochem. Cell Biol. 2003, 35, 802–806. [Google Scholar] [CrossRef]
- Vidt, D.G.; Prisant, L.M. Hypertensive heart disease. J. Clin. Hypertens. 2005, 7, 231–238. [Google Scholar] [CrossRef]
- Schnackenberg, C.G.; Welch, W.J.; Wilcox, C.S. Normalization of blood pressure and renal vascular resistance in SHR with a membrane permeable superoxide dismutase mimetic: Role of nitric oxide. Hypertension 1998, 32, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Wang, X.Q.; Oveisi, F.; Rad, B. Induction of oxidative stress by glutathione depletion causes severe hypertension in normal rats. Hypertension 2000, 36, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Redón, J.; Oliva, M.R.; Tormos, C.; Giner, V.; Chaves, J.; Iradi, A.; Sáez, G.T. Antioxidant activities and oxidative stress byproducts in human hypertension. Hypertension 2003, 41, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Lassègue, B.; Griendling, K.K. Reactive oxygen species in hypertension. An update. Am. J. Hypertens. 2004, 17, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Liang, K.; Ding, Y. Increased nitric oxide inactivation by reactive oxygen species in lead-induced hypertension. Kidney Int. 1999, 56, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Puzserova, A.; Bernatova, I. Blood pressure regulation in stress: Focus on nitric oxide-dependent mechanisms. Physiol. Res. 2016, 65, 309–342. [Google Scholar]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [PubMed]
- El Assar, M.; Angulo, J.; Rodríguez-Mañas, L. Oxidative stress and vascular inflammation in aging. Free Radic. Biol. Med. 2013, 65, 380–401. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.M.; Odendaal, L.; Du Toit, E.F.; Kupai, K.; Csont, T.; Ferdinandy, P.; van Rooyen, J. The effect of dietary red palm oil on the functional recovery of the ischaemic/reperfused isolated rat heart: The involvement of the PI3-Kinase signaling pathway. Lipids Health Dis. 2009, 8, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bester, D.J.; Kupai, K.; Tamas, C.; Szucs, G.; Csonka, C.; Esterhuyse, A.J.; Ferdinandy, P.; Van Rooyen, J. Dietary red palm oil supplementation reduces myocardial infarct size in an isolated perfused rat heart model. Lipids Health Dis. 2010, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Katengua-Thamahane, E.; Engelbrecht, A.M.; Esterhuyse, A.J.; Van Rooyen, J. Inhibition of Akt Attenuates RPO-Induced Cardioprotection. Cardiol. Res. Pract. 2012, 392457, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, R.; Subramaniam, K.M.; Radhakrishnan, A.K.; Choo, Y.M.; Teng, K.T. Health-promoting effects of red palm oil: Evidence from animal and human studies. Nutr. Rev. 2017, 75, 98–113. [Google Scholar] [CrossRef] [PubMed]
- Bester, D.; Esterhuyse, A.J.; Truter, E.J.; van Rooyen, J. Cardiovascular effects of edible oils: A comparison between four popular edible oils. Nutr. Res. Rev. 2010, 23, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Esterhuyse, J.S.; van Rooyen, J.; Strijdom, H.; Bester, D.; du Toit, E.F. Proposed mechanisms for red palm oil induced cardioprotection in a model of hyperlipidaemia in the rat. Prostaglandins Leukot Essent Fatty Acids 2006, 75, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Szucs, G.; Bester, D.J.; Kupai, K.; Csont, T.; Csonka, C.; Esterhuyse, A.J.; Ferdinandy, P.; Van Rooyen, J. Dietary red palm oil supplementation decreases infarct size in cholesterol fed rats. Lipids Health Dis. 2011, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Wergeland, A.; Bester, D.J.; Sishi, B.J.; Engelbrecht, A.M.; Jonassen, A.K.; Van Rooyen, J. Dietary red palm oil protects the heart against the cytotoxic effects of anthracycline. Cell Biochem. Funct. 2011, 29, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Bačová, B.; Radošinská, J.; Viczenczová, C.; Knezl, V.; Dosenko, V.; Beňova, T.; Navarová, J.; Gonçalvesová, E.; van Rooyen, J.; Weismann, P.; et al. Up-regulation of myocardial connexin-43 in spontaneously hypertensive rats fed red palm oil is most likely implicated in its anti-arrhythmic effects. Can. J. Physiol. Phamarcol. 2012, 90, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Boon, C.M.; Ng, M.H.; Choo, Y.M.; Mok, S.L. Super, Red Palm and Palm Oleins Improve the Blood Pressure, Heart Size, Aortic Media Thickness and Lipid Profile in Spontaneously Hypertensive Rats. PLoS ONE 2013, 8, 55908. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, R.M.; Zhang, H.; Vogel, H.; Cartwright, J., Jr.; Dionne, L.; Lu, N.; Huang, S.; Matzuk, M.M. Neurodegeneration, myocardial injury, and perinatal death in mitochondrial superoxide dismutase-deficient mice. Proc. Natl. Acad. Sci. USA 1996, 93, 9782–9787. [Google Scholar] [CrossRef] [PubMed]
- Kokoszka, J.E.; Coskun, P.; Esposito, L.A.; Wallace, D.C. Increased mitochondrial oxidative stress in the SOD2 [+/−] mouse results in the age related decline of mitochondrial function culminating in increased apoptosis. Proc. Natl. Acad. Sci. USA 2001, 98, 2278–2283. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Iturbe, B.; Sepassi, L.; Quiroz, Y.; Ni, Z.; Wallace, D.C.; Vaziri, N.D. Association of mitochondrial SOD deficiency with salt-sensitive hypertension and accelerated renal senescence. J. Appl. Physiol. 2007, 102, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Dikalova, A.E.; Bikineyeva, A.T.; Budzyn, K.; Nazarewicz, R.R.; McCann, L.; Lewis, W.; Harrison, D.G.; Dikalov, S.I. Therapeutic targeting of mitochondrial superoxide in hypertension. Circ. Res. 2010, 107, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.; Cenini, G.; Di Domenico, F.; Noel, T.; Wang, C.; Perluigi, M.; St Clair, D.K.; Butterfield, D.A. Basal brain oxidative and nitrative stress levels are finely regulated by the interplay between superoxide dismutase 2 and p. 53. J. Neurosci. Res. 2015, 93, 1728–1739. [Google Scholar] [CrossRef] [PubMed]
- Melov, S.; Coskun, P.; Patel, M.; Tuinstra, R.; Cottrell, B.; Jun, A.S.; Zastawny, T.H.; Dizdaroglu, M.; Goodman, S.I.; Huang, T.T.; et al. Mitochondrial disease in superoxide dismutase 2 mutant mice. Proc. Natl. Acad. Sci. USA 1999, 96, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Mügge, A.; Elwell, J.H.; Peterson, T.E.; Harrison, D.G. Release of intact endothelium-derived relaxing factor depends on endothelial superoxide dismutase activity. Am. J. Physiol. 1991, 260, 219–225. [Google Scholar]
- Cuzzocrea, S.; Mazzon, E.; Dugo, L.; di Paola, R.; Caputi, A.P.; Salvemini, D. Superoxide: A key player in hypertension. FASEB J. 2004, 18, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Tribulova, N.; Szeiffova Bacova, B.; Benova, T.; Viczenczova, C. Can we protect from malignant arrhythmias by modulation of cardiac cell-to-cell coupling? J. Electrocardiol. 2015, 48, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Radošinská, J.; Knezl, V.; Benová, T.; Urban, L.; Tribulová, N.; Slezák, J. Alterations of the intercellular coupling protein, connexin-43, during ventricular fibrillation and sinus rhythm restoration demonstrated in male and female rat hearts: A pilot study. Exp. Clin. Cardiol. 2011, 16, 116–120. [Google Scholar] [PubMed]
- Brinson, K.N.; Rafikova, O.; Sullivan, J.C. Female sex hormones protect against salt-sensitive hypertension but not essential hypertension. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Janega, P.; Klimentová, J.; Barta, A.; Kovácsová, M.; Vranková, S.; Cebová, M.; Čierna, Z.; Matúsková, Z.; Jakovljevic, V.; Pechánová, O. Red wine extract decreases pro-inflammatory markers, nuclear factor-κB and inducible NOS, in experimental metabolic syndrome. Food Funct. 2014, 5, 2202–2207. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Ni, Z.; Oveisi, F.; Trnavsky-Hobbs, D.L. Effect of antioxidant therapy on blood pressure and NO synthase expression in hypertensive rats. Hypertension 2000, 36, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst. Rev. 2012, 14, CD007176. [Google Scholar]
- Kloner, R.A.; Brown, D.A.; Csete, M.; Dai, W.; Downey, J.M.; Gottlieb, R.A.; Hale, S.L.; Shi, J. New and revisited approaches to preserving the reperfused myocardium. Nat. Rev. Cardiol. 2017, 14, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Ledvényiová-Farkašová, V.; Bernátová, I.; Balis, P.; Puzserova, A.; Barteková, M.; Gablovsky, I.; Ravingerová, T. Effect of crowding stress on tolerance to ischemia-reperfusion injury in young male and female hypertensive rats: Molecular mechanisms. Can. J. Physiol. Pharmacol. 2015, 93, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Benova, T.; Viczenczova, C.; Radosinska, J.; Bacova, B.; Knezl, V.; Dosenko, V.; Weismann, P.; Zeman, M.; Navarova, J.; Tribulova, N. Melatonin attenuates hypertension-related proarrhythmic myocardial maladaptation of connexin-43 and propensity of the heart to lethal arrhythmias. Can. J. Physiol. Pharmacol. 2013, 91, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Puzserova, A.; Slezak, P.; Balis, P.; Bernatova, I. Long-term social stress induces nitric oxide-independent endothelial dysfunction in normotensive rats. Stress 2013, 16, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Bernátová, I.; Pechánová, O.; Babál, P.; Kyselá, S.; Stvrtina, S.; Andriantsitohaina, R. Wine polyphenols improve cardiovascular remodeling and vascular function in NO-deficient hypertension. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | WKYc | WKYrpo | SHRc | SHRrpo |
|---|---|---|---|---|
| BP (mmHg) | 114.42 ± 11.53 | 120.71 ± 14.64 | 185.73 ± 12.52 * | 134.72 ± 10.94 # |
| BW (g) | 400.73 ± 20.22 | 378.21 ± 31.22 | 311.44 ± 18.11 * | 332.63 ± 20.94 |
| HW (g) | 1.33 ± 0.08 | 1.34 ± 0.09 | 1.51 ± 0.06 | 1.47 ± 0.04 |
| LVW (g) | 0.72 ± 0.03 | 0.74 ± 0.06 | 0.95 ± 0.03 * | 0.89 ± 0.07 |
| BG (mg/dL) | 5.49 ± 0.83 | 4.98 ± 0.91 | 6.12 ± 0.82 | 5.78 ± 0.73 |
| CHOL (mmol/L) | 2.43 ± 0.42 | 2.38 ± 0.33 | 2.78 ± 0.62 | 2.74 ± 0.22 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katengua-Thamahane, E.; Szeiffova Bacova, B.; Bernatova, I.; Sykora, M.; Knezl, V.; Van Rooyen, J.; Tribulova, N. Effects of Red Palm Oil on Myocardial Antioxidant Enzymes, Nitric Oxide Synthase and Heart Function in Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2017, 18, 2476. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112476
Katengua-Thamahane E, Szeiffova Bacova B, Bernatova I, Sykora M, Knezl V, Van Rooyen J, Tribulova N. Effects of Red Palm Oil on Myocardial Antioxidant Enzymes, Nitric Oxide Synthase and Heart Function in Spontaneously Hypertensive Rats. International Journal of Molecular Sciences. 2017; 18(11):2476. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112476
Chicago/Turabian StyleKatengua-Thamahane, Emma, Barbara Szeiffova Bacova, Iveta Bernatova, Matus Sykora, Vladimir Knezl, Jacques Van Rooyen, and Narcis Tribulova. 2017. "Effects of Red Palm Oil on Myocardial Antioxidant Enzymes, Nitric Oxide Synthase and Heart Function in Spontaneously Hypertensive Rats" International Journal of Molecular Sciences 18, no. 11: 2476. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112476