Crosstalk of PmCBFs and PmDAMs Based on the Changes of Phytohormones under Seasonal Cold Stress in the Stem of Prunus mume

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
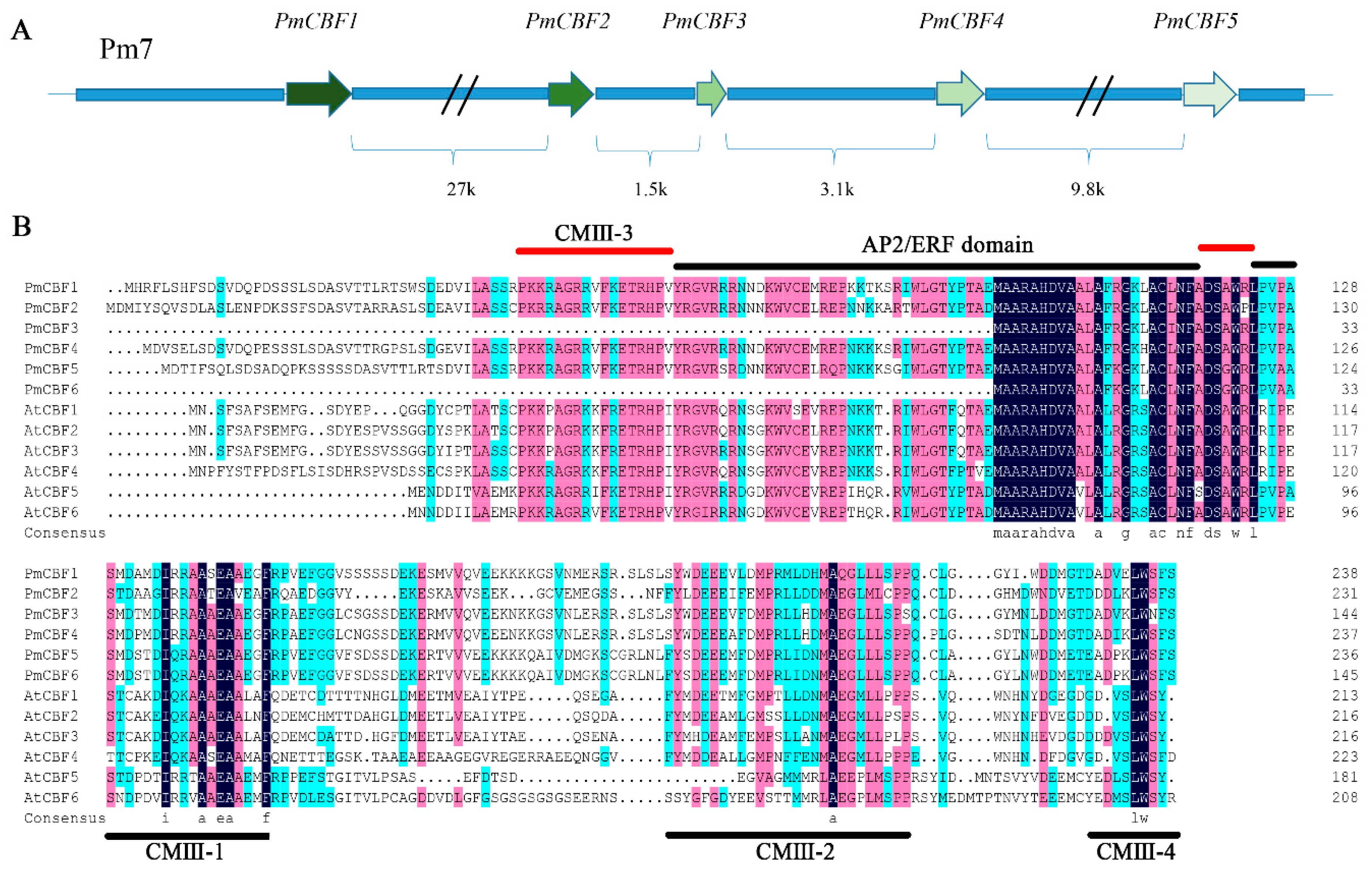
2.1. Identification and Cloning of CBF Genes in P. mume
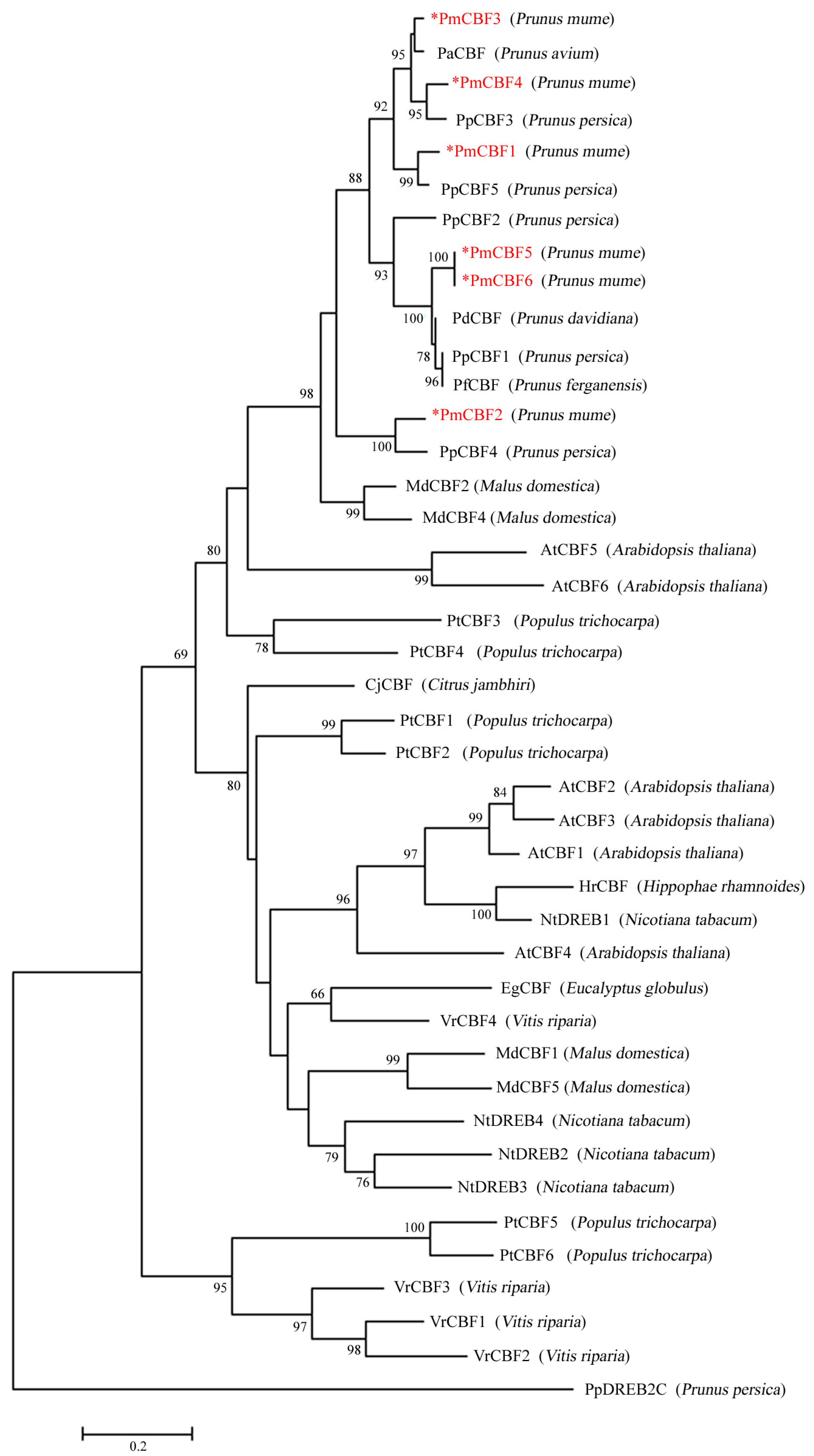
2.2. Multiple Sequence Alignments and Phylogenetic Analysis
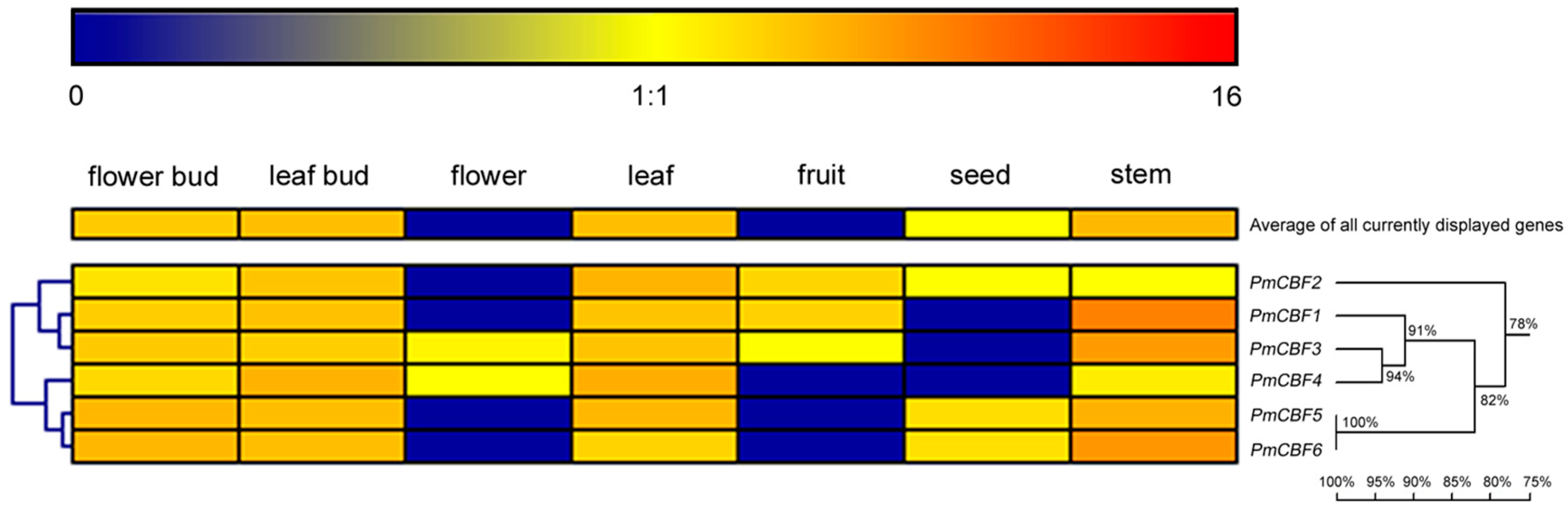
2.3. Expressions of PmCBFs in Different Organs
2.4. Sectionalization of PmCBFs and PmDAMs during the Expression in the Stem
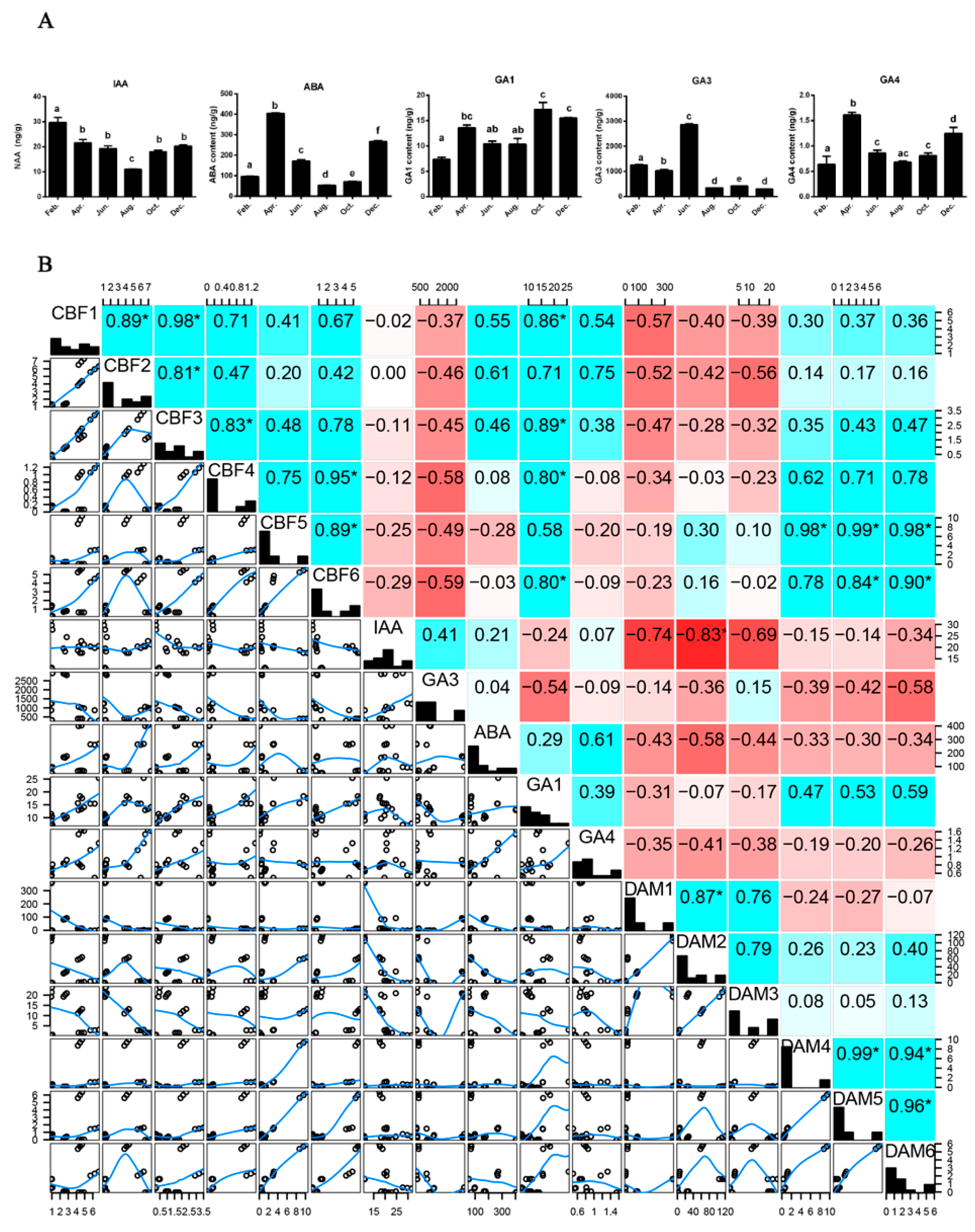
2.5. Seasonal Changes in Hormones Synthesis and Correlations with Gene Expression during Stem Development
2.6. Subcellular Localization Assessment
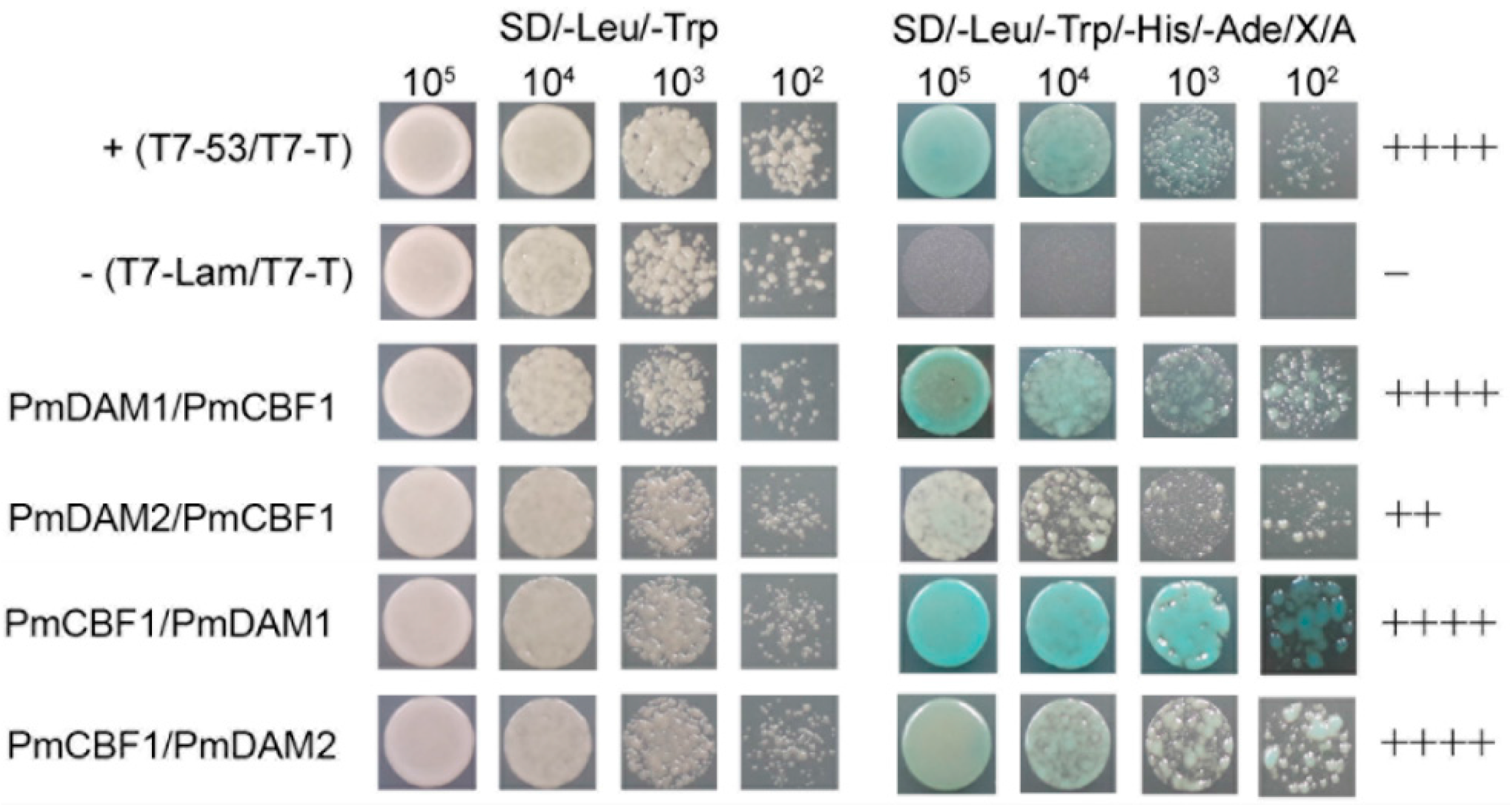
2.7. Protein-Protein Interactions between CBF and DAM Genes in P. mume
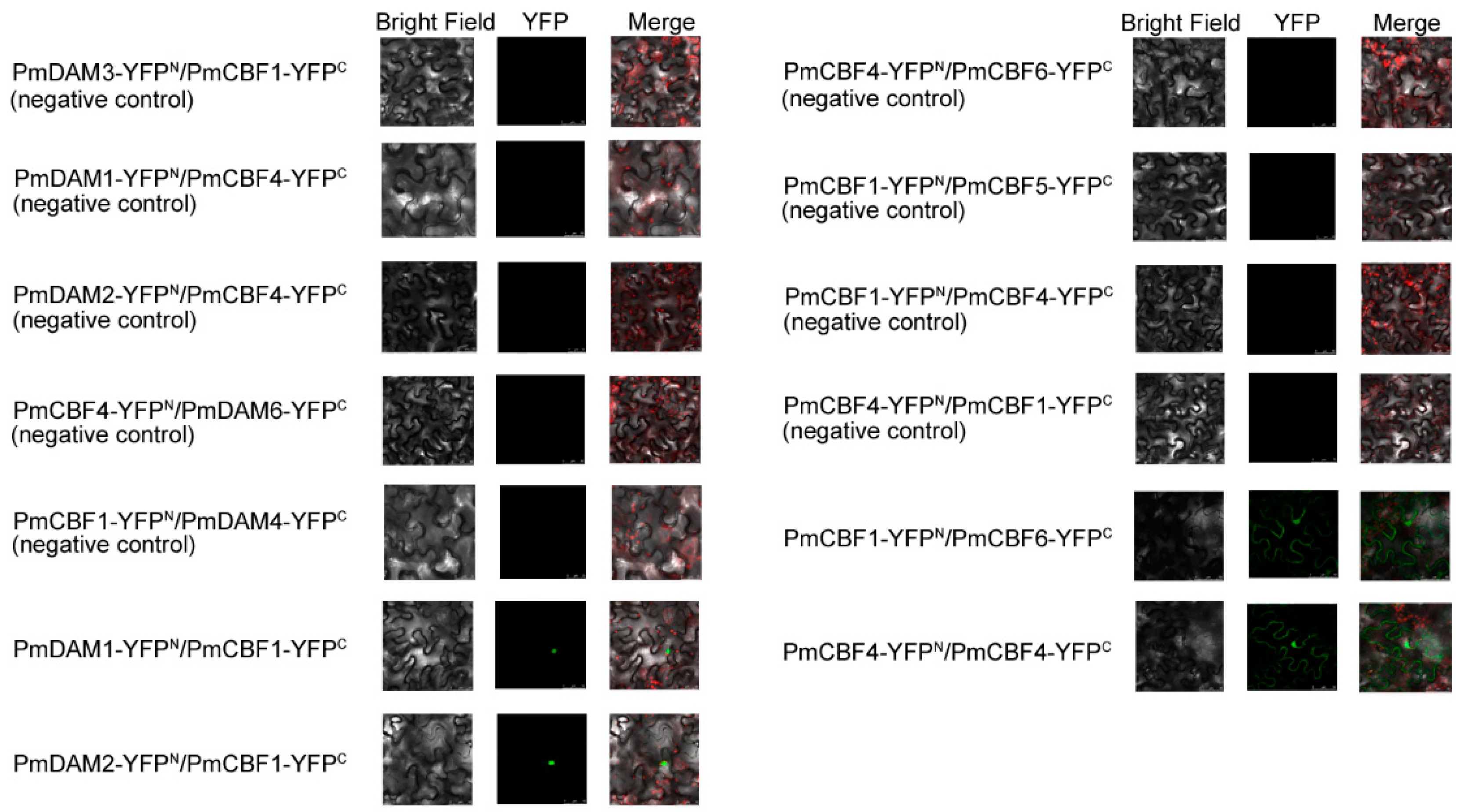
2.8. BiFC Assay
3. Discussion
3.1. CBF Genes in P. mume and Their Evolution
3.2. Positive Response to the Coldness by PmCBFs and PmDAM4–6
3.3. The Interactive Roles of Major Hormones
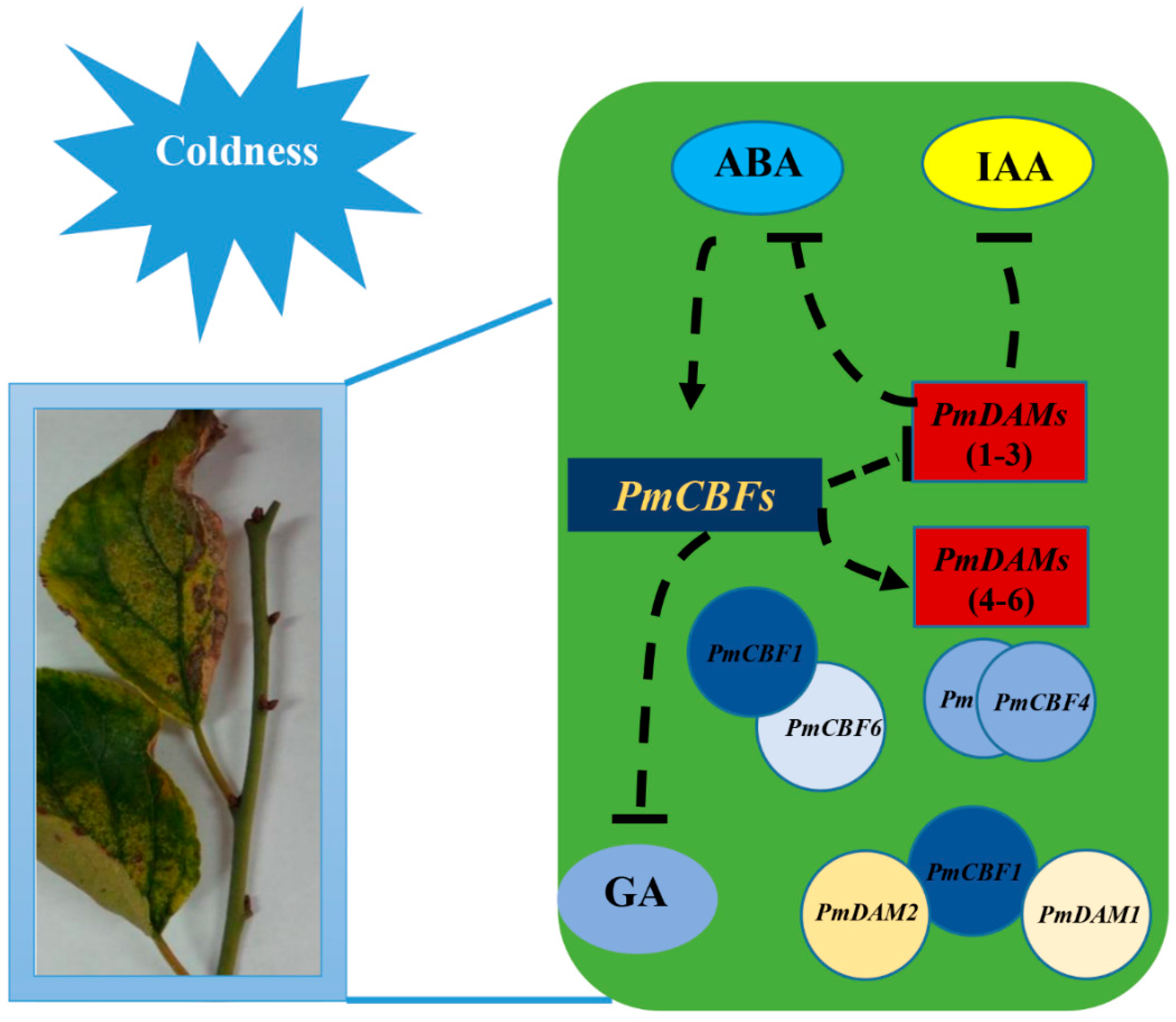
3.4. Proposed Regulation Network between Hormones and PmCBF and PmDAMs in Stem Cold Response
4. Material and Methods
4.1. Plant Materials
4.2. Identification and Cloning of CBF Genes
4.3. Phylogenetic Analyses
4.4. RT-qPCR
4.5. Subcellular Localization Assessments
4.6. Yeast Two-Hybrid Assays
4.7. BiFC Assays
4.8. Phytohormone Assays
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, G.; Li, W.; Zheng, P.; Xu, T.; Chen, L.; Liu, D.; Hussain, S.; Teng, Y. Transcriptomic analysis of ‘suli’ pear (Pyrus pyrifolia white pear group) buds during the dormancy by rna-seq. BMC Genom. 2012, 13, 700. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Saito, T.; Sakamoto, D.; Ito, A.; Fujii, H.; Moriguchi, T. Transcriptome analysis of japanese pear (Pyrus pyrifolia nakai) flower buds transitioning through endodormancy. Plant Cell Physiol. 2013, 54, 1132–1151. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Li, J.; Cai, D.; Qian, M.; Jia, H.; Bai, S.; Hussain, S.; Liu, G.; Teng, Y.; Zheng, X. Dormancy-associated mads-box genes and micrornas jointly control dormancy transition in pear (Pyrus pyrifolia white pear group) flower bud. J. Exp. Bot. 2016, 67, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.B.; Yang, Y.H.; Zhang, Z.J.; Chen, J.; Wang, X.C.; Huang, R.F. Expression of the ethylene response factor gene tsrf1 enhances abscisic acid responses during seedling development in tobacco. Planta 2008, 228, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Rashotte, A.M.; Goertzen, L.R. The crf domain defines cytokinin response factor proteins in plants. BMC Plant Biol. 2010, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.R.; Wang, M.L.; Lynch, T.J.; Rao, S.; Goodman, H.M. The arabidopsis abscisic acid response locus abi4 encodes an apetala 2 domain protein. Plant Cell 1998, 10, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Hirota, A.; Kato, T.; Fukaki, H.; Aida, M.; Tasaka, M. The auxin-regulated ap2/erebp gene puchi is required for morphogenesis in the early lateral root primordium of arabidopsis. Plant Cell 2007, 19, 2156–2168. [Google Scholar] [CrossRef] [PubMed]
- Horvath, D.P.; Chao, W.S.; Suttle, J.C.; Thimmapuram, J.; Anderson, J.V. Transcriptome analysis identifies novel responses and potential regulatory genes involved in seasonal dormancy transitions of leafy spurge (Euphorbia esula L.). BMC Genom. 2008, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Pre, M.; Atallah, M.; Champion, A.; De Vos, M.; Pieterse, C.M.; Memelink, J. The ap2/erf domain transcription factor ora59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol. 2008, 147, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; van Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. Hre1 and hre2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J. 2010, 62, 302–315. [Google Scholar] [CrossRef] [PubMed]
- McKhann, H.I.; Gery, C.; Berard, A.; Leveque, S.; Zuther, E.; Hincha, D.K.; De Mita, S.; Brunel, D.; Teoule, E. Natural variation in CBF gene sequence, gene expression and freezing tolerance in the versailles core collection of Arabidopsis thaliana. BMC Plant Biol. 2008, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, dreb1 and dreb2, with an erebp/ap2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the erf gene family in arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Osdreb genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Xu, Z.; Xia, L.; Li, L.; Cheng, X.; Dong, J.; Wang, Q.; Ma, Y. Cold-induced modulation and functional analyses of the dre-binding transcription factor gene, gmdreb3, in soybean (Glycine max L.). J. Exp. Bot. 2009, 60, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Hong, B.; Yang, Y.; Li, Q.; Ma, N.; Ma, C.; Gao, J. Overexpression of two chrysanthemum dgdreb1 group genes causing delayed flowering or dwarfism in arabidopsis. Plant Mol. Boil. 2009, 71, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.; Ayax, C.; Martinez, Y.; Laur, J.; El Kayal, W.; Marque, C.; Teulieres, C. Two egucbf1 genes overexpressed in Eucalyptus display a different impact on stress tolerance and plant development. Plant Biotechnol. J. 2011, 9, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Siddiqua, M.; Nassuth, A. Vitis cbf1 and vitis cbf4 differ in their effect on Arabidopsis abiotic stress tolerance, development and gene expression. Plant Cell Environ. 2011, 34, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, X.D.; Chi, X.J.; Wu, C.A.; Li, Y.Z.; Song, L.L.; Liu, X.M.; Wang, Y.F.; Wang, F.W.; Zhang, C.; et al. Dwarf apple mbdreb1 enhances plant tolerance to low temperature, drought, and salt stress via both aba-dependent and aba-independent pathways. Planta 2011, 233, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Stockinger, E.J.; Gilmour, S.J.; Thomashow, M.F. Arabidopsis thaliana cbf1 encodes an ap2 domain-containing transcriptional activator that binds to the c-repeat/dre, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 1997, 94, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Sakuma, Y.; Kasuga, M.; Ito, Y.; Seki, M.; Goda, H.; Shimada, Y.; Yoshida, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Identification of cold-inducible downstream genes of the Arabidopsis dreb1a/cbf3 transcriptional factor using two microarray systems. Plant J. 2004, 38, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Novillo, F.; Alonso, J.M.; Ecker, J.R.; Salinas, J. Cbf2/dreb1c is a negative regulator of cbf1/dreb1b and cbf3/dreb1a expression and plays a central role in stress tolerance in arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 3985–3990. [Google Scholar] [CrossRef] [PubMed]
- Novillo, F.; Medina, J.; Salinas, J. Arabidopsis cbf1 and cbf3 have a different function than cbf2 in cold acclimation and define different gene classes in the CBF regulon. Proc. Natl. Acad. Sci. USA 2007, 104, 21002–21007. [Google Scholar] [CrossRef] [PubMed]
- Haake, V.; Cook, D.; Riechmann, J.L.; Pineda, O.; Thomashow, M.F.; Zhang, J.Z. Transcription factor cbf4 is a regulator of drought adaptation in arabidopsis. Plant Physiol. 2002, 130, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. Dwarf and delayed-flowering 1, a novel arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative ap2 transcription factor. Plant J. 2004, 37, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. The ddf1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, ga2ox7, under high-salinity stress in arabidopsis. Plant J. 2008, 56, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Norelli, J.; Artlip, T. Overexpression of a peach CBF gene in apple: A model for understanding the integration of growth, dormancy, and cold hardiness in woody plants. Front. Plant Sci. 2015, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Artlip, T.S.; Wisniewski, M.E.; Arora, R.; Norelli, J.L. An apple rootstock overexpressing a peach CBF gene alters growth and flowering in the scion but does not impact cold hardiness or dormancy. Hortic. Res. 2016, 3, 16006. [Google Scholar] [CrossRef] [PubMed]
- Artlip, T.S.; Wisniewski, M.E.; Bassett, C.L.; Norelli, J.L. Cbf gene expression in peach leaf and bark tissues is gated by a circadian clock. Tree Physiol. 2013, 33, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Norelli, J.; Bassett, C.; Artlip, T.; Macarisin, D. Ectopic expression of a novel peach (Prunus persica) CBF transcription factor in apple (malus × domestica) results in short-day induced dormancy and increased cold hardiness. Planta 2011, 233, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.G.; Zhang, W.K.; He, S.J.; Zhang, J.S.; Liu, Q.; Chen, S.Y. An erebp/ap2-type protein in triticum aestivum was a dre-binding transcription factor induced by cold, dehydration and aba stress. Theor. Appl. Genet. 2003, 106, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, L.; Hancock, R.D.; Haupt, S.; Walker, P.G.; Pont, S.D.; McNicol, J.; Cardle, L.; Morris, J.; Viola, R.; Brennan, R.; et al. Co-ordinated gene expression during phases of dormancy release in raspberry (Rubus idaeus L.) buds. J. Exp. Bot. 2007, 58, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Arya, P.; Gupta, K.; Randhawa, V.; Acharya, V.; Singh, A.K. Comparative phylogenetic analysis and transcriptional profiling of mads-box gene family identified dam and flc-like genes in apple (Malusx domestica). Sci. Rep. 2016, 6, 20695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G.; et al. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, Q.; Sun, L.; Du, D.; Cheng, T.; Pan, H.; Yang, W.; Wang, J. Genome-wide identification, characterisation and expression analysis of the mads-box gene family in Prunus mume. Mol. Genet. Genom. 2014, 289, 903–920. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, R.; Yamane, H.; Ooka, T.; Jotatsu, H.; Kitamura, Y.; Akagi, T.; Tao, R. Functional and expressional analyses of pmdam genes associated with endodormancy in japanese apricot. Plant Physiol. 2011, 157, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Kashiwa, Y.; Ooka, T.; Tao, R.; Yonemori, K. Suppression subtractive hybridization and differential screening reveals endodormancy-associated expression of an svp/agl24-type mads-box gene in lateral vegetative buds of japanese apricot. J. Am. Soc. Hortic. Sci. 2008, 133, 708–716. [Google Scholar]
- Kitamura, Y.; Takeuchi, T.; Yamane, H.; Tao, R. Simultaneous down-regulation of dormancy-associated mads-box6 and soc1 during dormancy release in japanese apricot (Prunus mume) flower buds. J. Hortic. Sci. Biotechnol. 2016, 91, 1–7. [Google Scholar] [CrossRef]
- Li, Z.; Reighard, G.L.; Abbott, A.G.; Bielenberg, D.G. Dormancy-associated mads genes from the evg locus of peach [Prunus persica (L.) batsch] have distinct seasonal and photoperiodic expression patterns. J. Exp. Bot. 2009, 60, 3521–3530. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Bai, S.; Imai, T.; Ito, A.; Nakajima, I.; Moriguchi, T. Histone modification and signalling cascade of the dormancy-associated mads-box gene, ppmads13-1, in japanese pear (Pyrus pyrifolia) during endodormancy. Plant Cell Environ. 2015, 38, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Jaglo, K.R.; Thomashow, M.F. Components of the arabidopsis c-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in brassica napus and other plant species. Plant Physiol. 2001, 127, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Lee, H.; Jin, J.; Park, H.; Kim, J.; Noh, Y.S.; Lee, I. Crosstalk between cold response and flowering in arabidopsis is mediated through the flowering-time gene soc1 and its upstream negative regulator flc. Plant Cell 2009, 21, 3185–3197. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, C.; Kozarewa, I.; Johansson, M.; Ogren, E.; Rohde, A.; Eriksson, M.E. Circadian clock components regulate entry and affect exit of seasonal dormancy as well as winter hardiness in populus trees. Plant Physiol. 2010, 153, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Abdullah, S.N.; Abdul Aziz, M.; Namasivayam, P. Oil palm egcbf3 conferred stress tolerance in transgenic tomato plants through modulation of the ethylene signaling pathway. J. Plant Physiol. 2016, 202, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Nassuth, A.; Teulières, C.; Marque, C.; Rowland, J.; Cao, P.B.; Brown, A. Genomics of cold hardiness in woody plants. Crit. Rev. Plant Sci. 2014, 33, 92–124. [Google Scholar] [CrossRef]
- Porto, D.D.; Falavigna, V.D.S.; Arenhart, R.A.; Perini, P.; Buffon, V.; Anzanello, R.; Santos, H.P.D.; Fialho, F.B.; Oliveira, P.R.D.D.; Revers, L.F. Structural genomics and transcriptional characterization of the dormancy-associated mads-box genes during bud dormancy progression in apple. Tree Genet. Genom. 2016, 12, 46. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Dahal, K.P.; Savitch, L.V.; Singh, J.; Bode, R.; Ivanov, A.G.; Hurry, V.; Huner, N.P. Role of cbfs as integrators of chloroplast redox, phytochrome and plant hormone signaling during cold acclimation. Int. J. Mol. Sci. 2013, 14, 12729–12763. [Google Scholar] [CrossRef] [PubMed]
- Barrero-Gil, J.; Salinas, J. Cbfs at the crossroads of plant hormone signaling in cold stress response. Mol. Plant 2017, 10, 542–544. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Gao, Q.; Li, Z.; Chen, X.; Li, W. The role of gibberellin in the cbf1-mediated stress-response pathway. Plant Mol. Biol. Rep. 2014, 32, 852–863. [Google Scholar] [CrossRef]
- Achard, P.; Genschik, P. Releasing the brakes of plant growth: How gas shutdown della proteins. J. Exp. Bot. 2009, 60, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Gong, F.; Cheminant, S.; Alioua, M.; Hedden, P.; Genschik, P. The cold-inducible cbf1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing della proteins via its effect on gibberellin metabolism. Plant Cell 2008, 20, 2117–2129. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate regulates the inducer of CBF expression-c-repeat binding factor/dre binding factor1 cascade and freezing tolerance in arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Li, W.; Zhao, J.; Zhang, S.; Xu, H.; Liu, Y.; Guo, C. Transcriptome sequencing analysis of alfalfa reveals CBF genes potentially playing important roles in response to freezing stress. Genet. Mol. Biol. 2017, 40, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Knight, H.; Zarka, D.G.; Okamoto, H.; Thomashow, M.F.; Knight, M.R. Abscisic acid induces CBF gene transcription and subsequent induction of cold-regulated genes via the crt promoter element. Plant Physiol. 2004, 135, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, M.; Lee, J.H.; Lee, H.J.; Park, C.M. The unified ice-CBF pathway provides a transcriptional feedback control of freezing tolerance during cold acclimation in arabidopsis. Plant Mol. Biol. 2015, 89, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Tuan, P.A.; Bai, S.; Saito, T.; Ito, A.; Moriguchi, T. Dormancy-associated mads-box (dam) and the abscisic acid pathway regulate pear endodormancy through a feedback mechanism. Plant Cell Physiol. 2017, 58, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Etchells, J.P.; Provost, C.M.; Turner, S.R. Plant vascular cell division is maintained by an interaction between pxy and ethylene signalling. PLoS Genet. 2012, 8, e1002997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.; Segear, E.; Beers, L.; Knauber, D.; Suttle, J. Dormancy in potato tuber meristems: Chemically induced cessation in dormancy matches the natural process based on transcript profiles. Funct. Integr. Genom. 2008, 8, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Ruttink, T.; Arend, M.; Morreel, K.; Storme, V.; Rombauts, S.; Fromm, J.; Bhalerao, R.P.; Boerjan, W.; Rohde, A. A molecular timetable for apical bud formation and dormancy induction in poplar. Plant Cell 2007, 19, 2370–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidokoro, S.; Watanabe, K.; Ohori, T.; Moriwaki, T.; Maruyama, K.; Mizoi, J.; Myint Phyu Sin Htwe, N.; Fujita, Y.; Sekita, S.; Shinozaki, K.; et al. Soybean DREB1/CBF-type transcription factors function in heat and drought as well as cold stress-responsive gene expression. Plant J. 2015, 81, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; He, J.; Zhong, X.J.; Guo, H.D.; Jin, S.H.; Li, X.; Sun, L.X. Molecular cloning and characterization of a novel freezing-inducible dreb1/CBF transcription factor gene in boreal plant Iceland poppy (Papaver nudicaule). Genet. Mol. Boil. 2016, 39, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Yang, K.S.; Ryu, S.H.; Kim, K.Y.; Song, W.K.; Kwon, S.Y.; Lee, H.S.; Bang, J.W.; Kwak, S.S. Molecular characterization of a cdna encoding dre-binding transcription factor from dehydration-treated fibrous roots of sweetpotato. Plant Physiol. Biochem. 2008, 46, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhu, J.K. The broad roles of CBF genes: From development to abiotic stress. Plant Signal. Behav. 2016, 11, e1215794. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, Z.; Xie, S.; Si, T.; Li, Y.; Zhu, J.K. Mutational evidence for the critical role of CBF transcription factors in cold acclimation in arabidopsis. Plant Physiol. 2016, 171, 2744–2759. [Google Scholar] [PubMed]
- Wang, T.; Hao, R.; Pan, H.; Cheng, T.; Zhang, Q. Selection of suitable reference genes for quantitative real-time polymerase chain reaction in Prunus mume during flowering stages and under different abiotic stress conditions. J. Am. Soc. Hortic. Sci. 2014, 139, 113–122. [Google Scholar]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, K.; Zhou, Y.; Li, Y.; Zhuo, X.; Ahmad, S.; Han, Y.; Yong, X.; Zhang, Q. Crosstalk of PmCBFs and PmDAMs Based on the Changes of Phytohormones under Seasonal Cold Stress in the Stem of Prunus mume. Int. J. Mol. Sci. 2018, 19, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020015
Zhao K, Zhou Y, Li Y, Zhuo X, Ahmad S, Han Y, Yong X, Zhang Q. Crosstalk of PmCBFs and PmDAMs Based on the Changes of Phytohormones under Seasonal Cold Stress in the Stem of Prunus mume. International Journal of Molecular Sciences. 2018; 19(2):15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020015
Chicago/Turabian StyleZhao, Kai, Yuzhen Zhou, Yushu Li, Xiaokang Zhuo, Sagheer Ahmad, Yu Han, Xue Yong, and Qixiang Zhang. 2018. "Crosstalk of PmCBFs and PmDAMs Based on the Changes of Phytohormones under Seasonal Cold Stress in the Stem of Prunus mume" International Journal of Molecular Sciences 19, no. 2: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020015