Preferential Inhibition of Wnt/β-Catenin Signaling by Novel Benzimidazole Compounds in Triple-Negative Breast Cancer

 ,
,
Abstract
:1. Introduction
2. Results
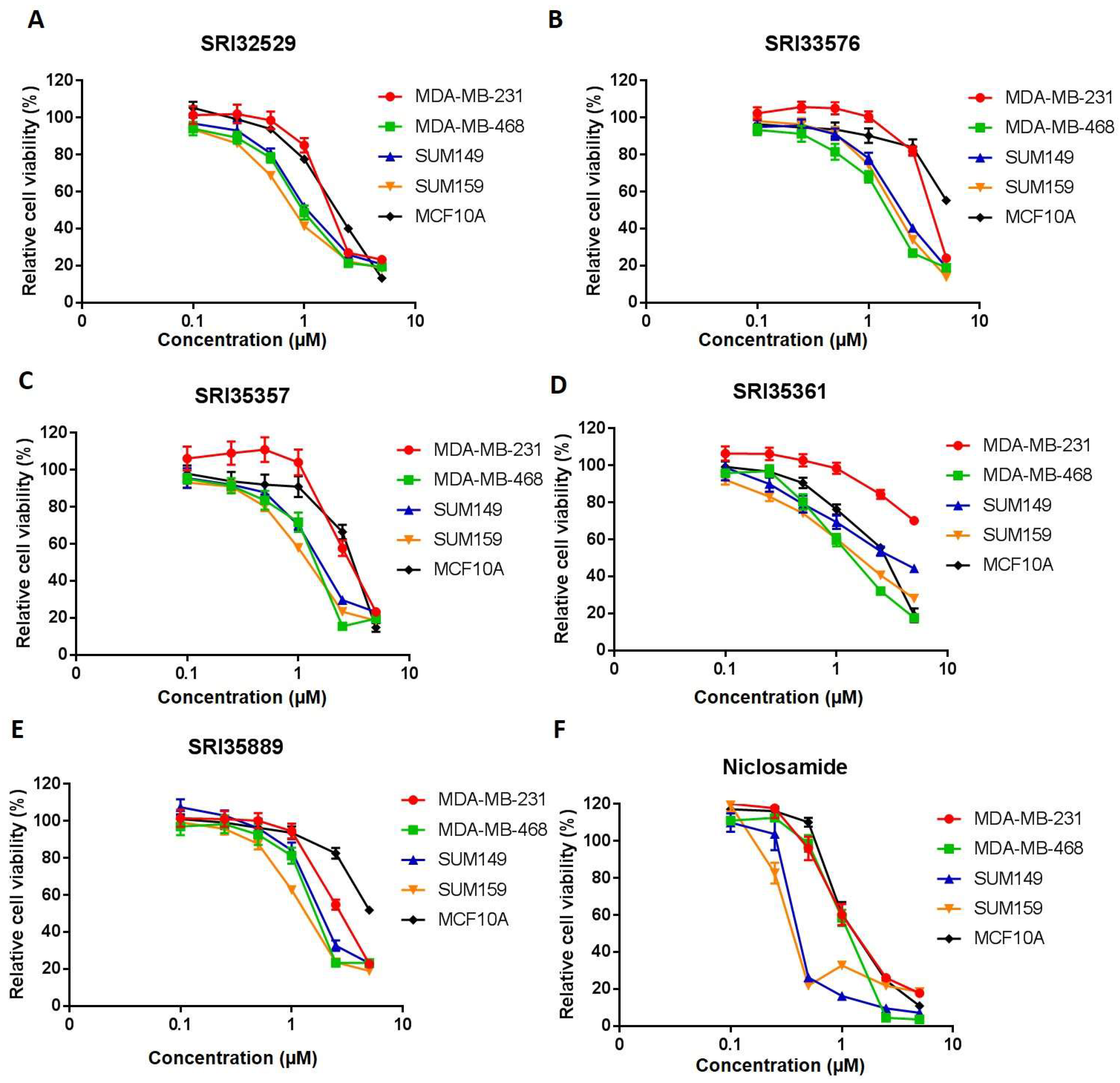
2.1. SRI33576 and SRI35889 are More Cytotoxic to TNBC Than to Noncancerous MCF10A Cells
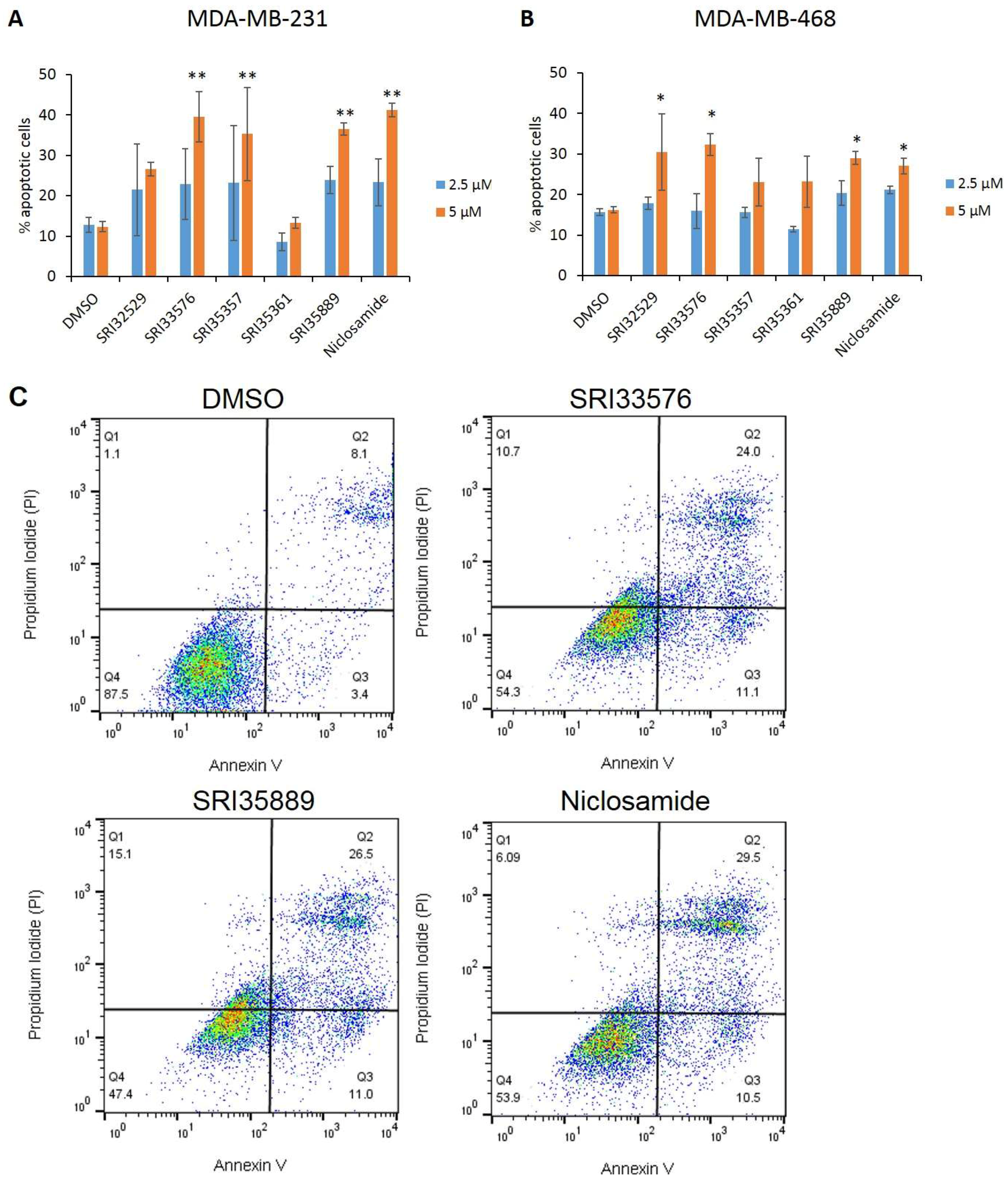
2.2. SRI33576 and SRI35889 Induce Apoptosis in TNBC
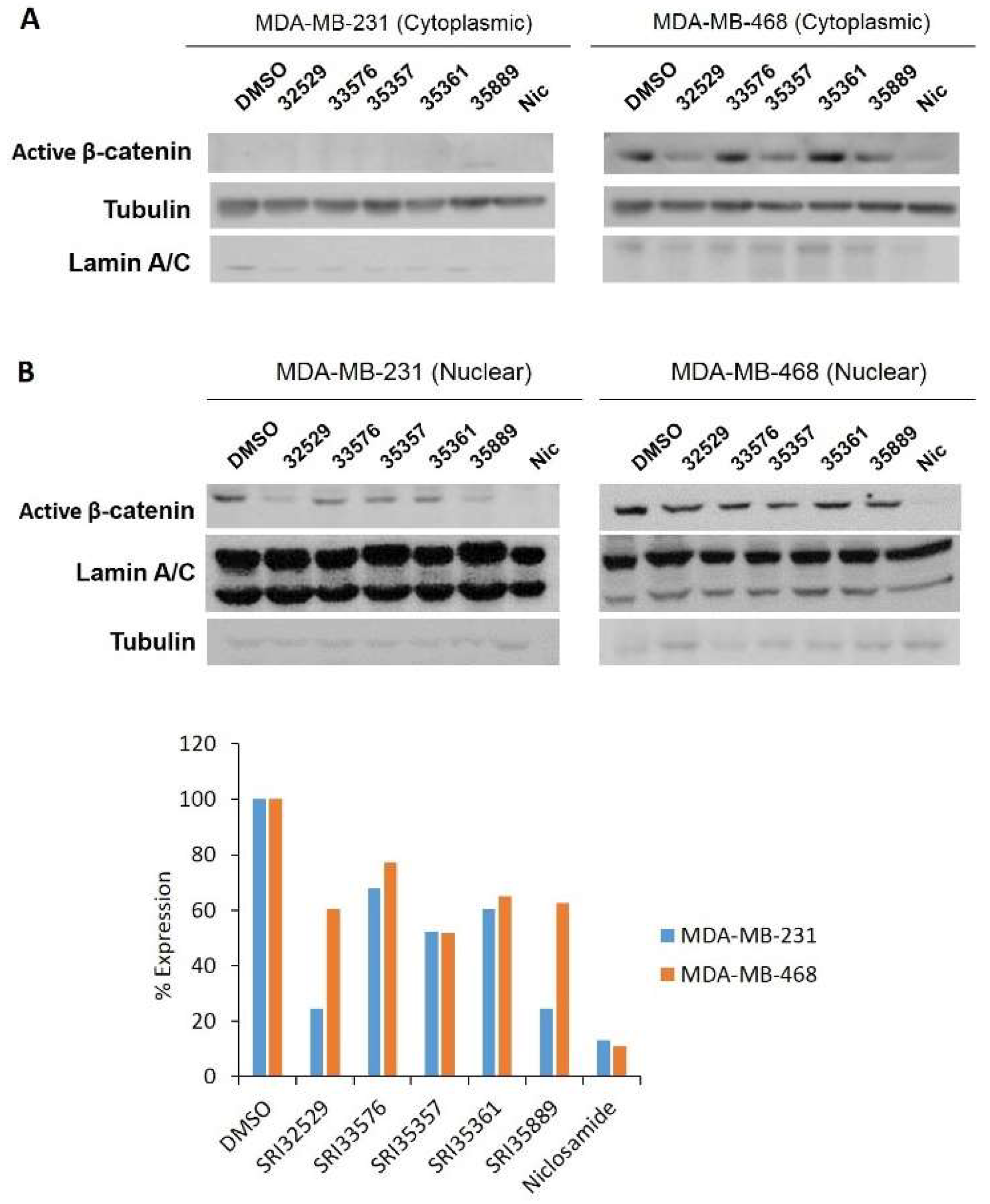
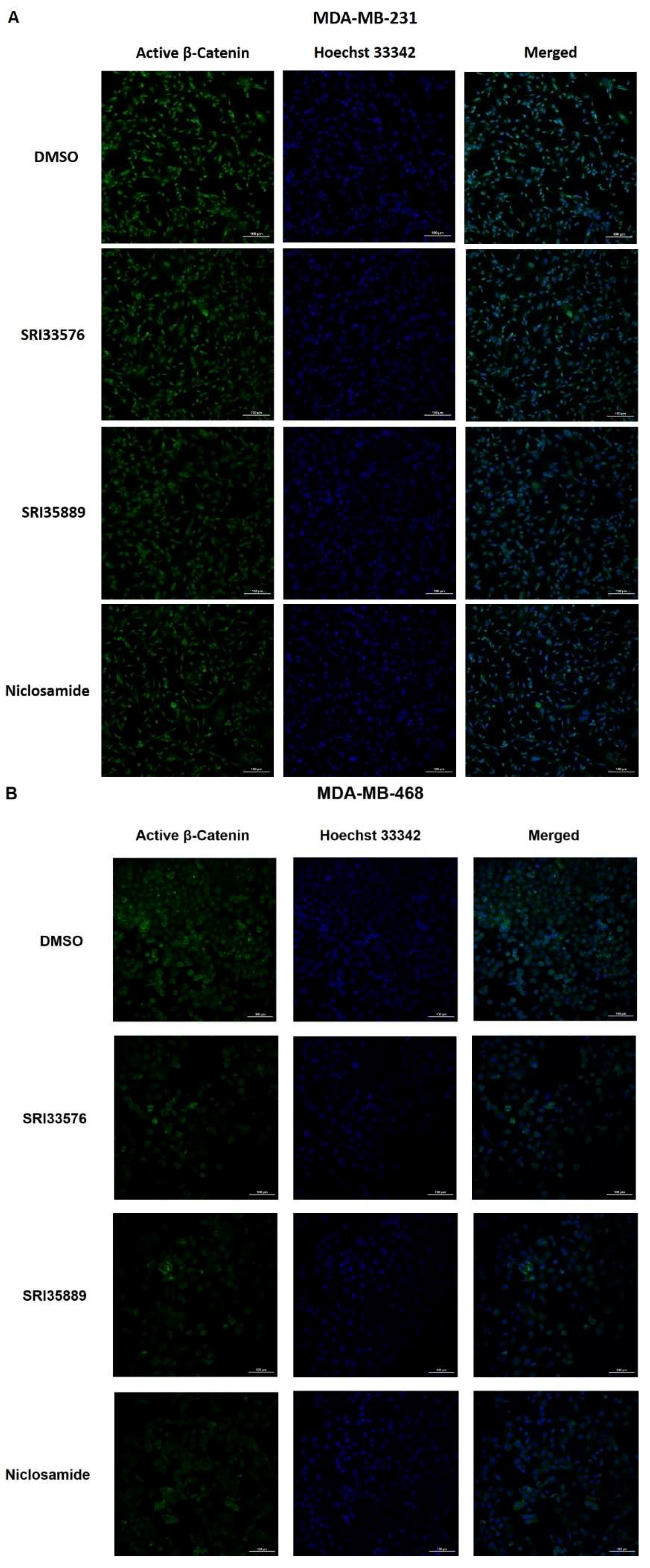
2.3. SRI33576 and SRI35889 Inhibit Wnt/β-Catenin Signaling in TNBC Cell Lines But Not Noncancerous Cells
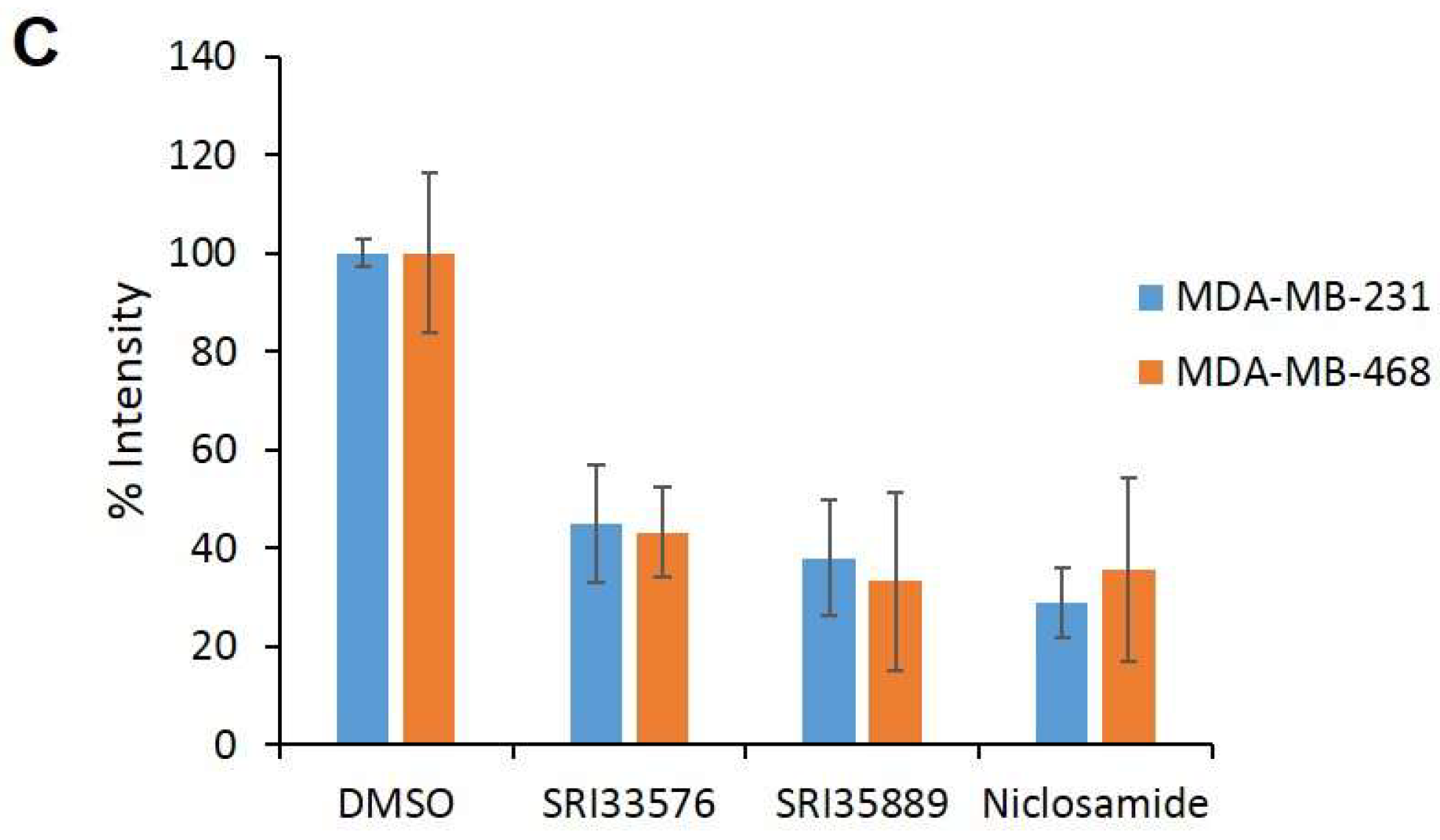
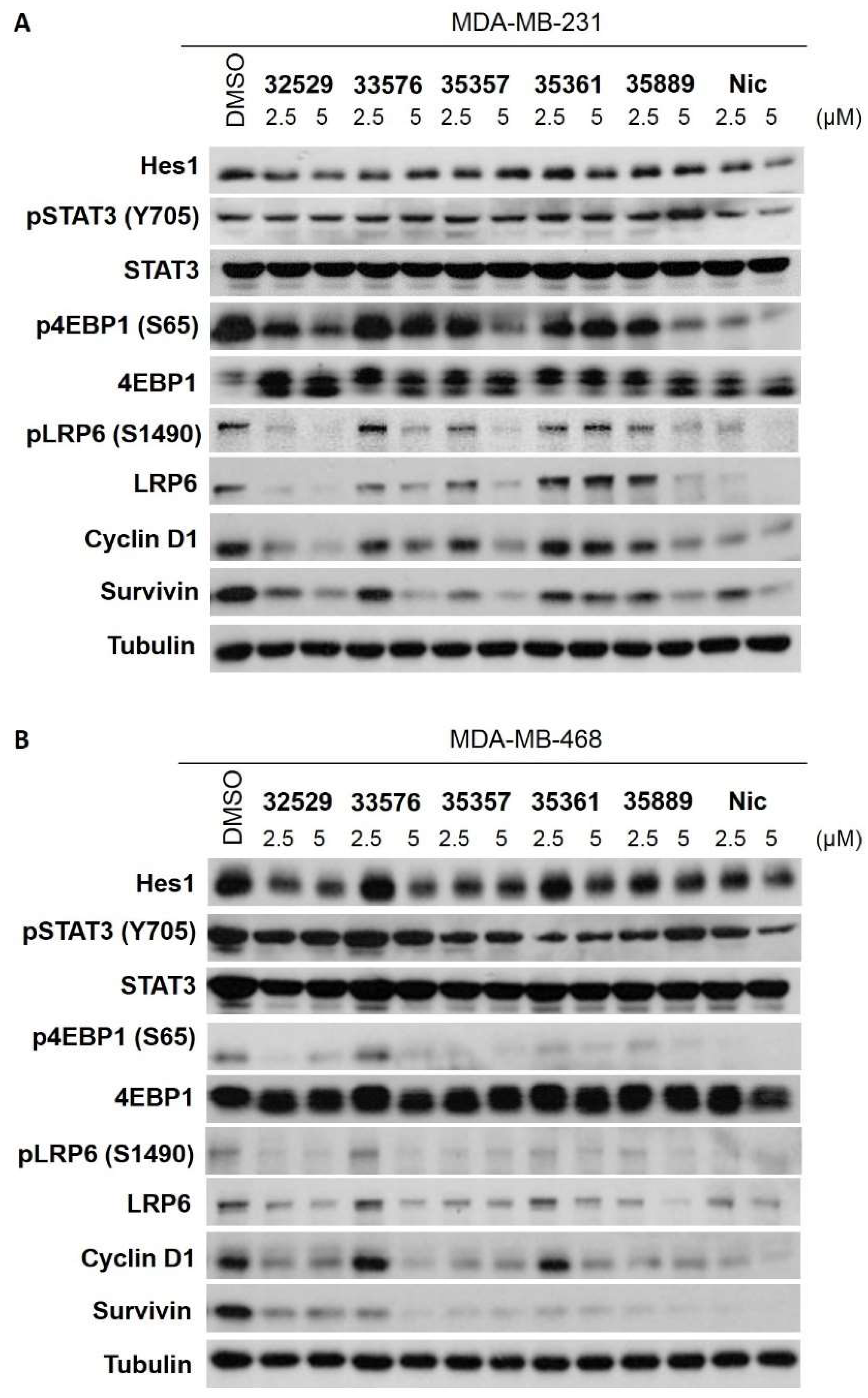
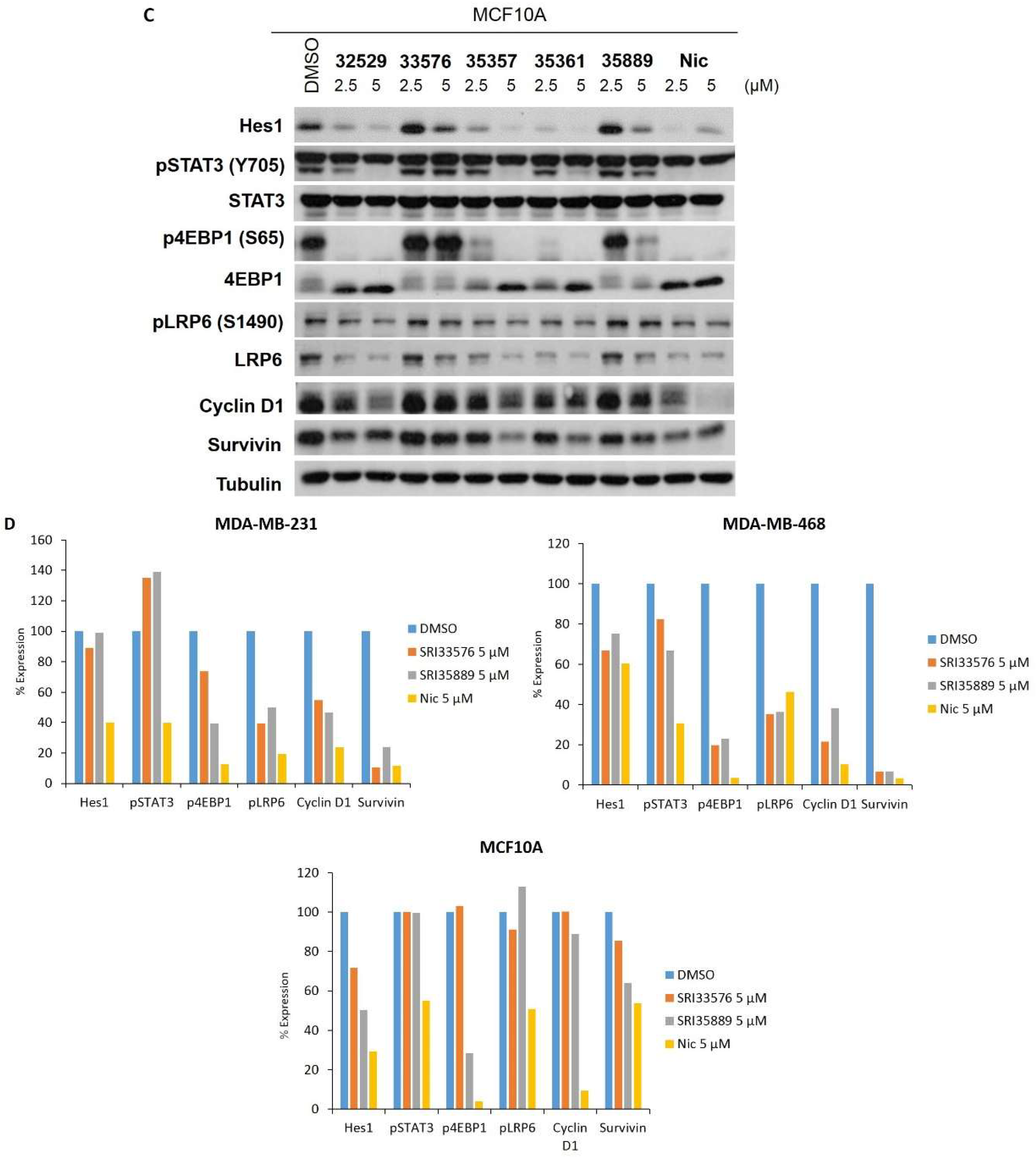
2.4. SRI33576 and SRI35889 Have Less Effect on mTOR, STAT3 and Notch Signaling Than Niclosamide
3. Discussion
4. Materials and Methods
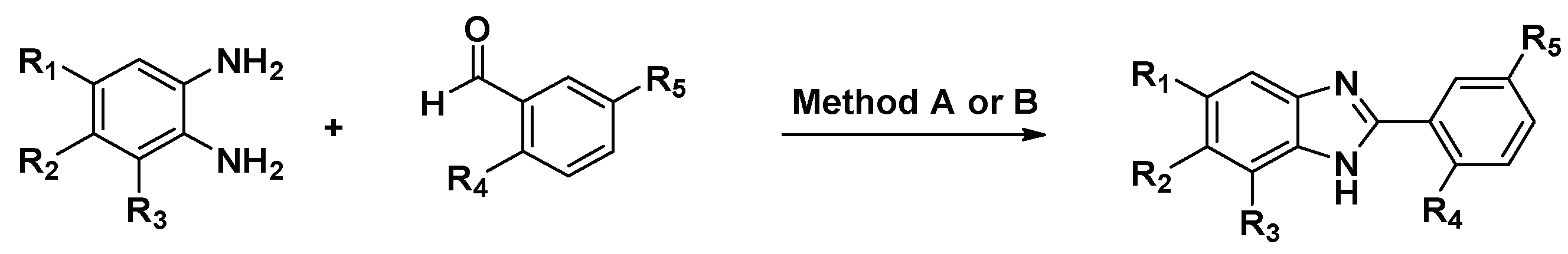
4.1. Compound Synthesis
4.1.1. Method A
4.1.2. Method B
4.1.3. 2-(5,7-Dichloro-3H-benzo[d]imidazol-2-yl)-4-fluorophenol (35357)
4.1.4. 2-(5-Chloro-6-fluoro-1H-benzo[d]imidazol-2-yl)-4-fluorophenol (35361).
4.1.5. 4-Chloro-2-(5,6-dichloro-1H-benzo[d]imidazol-2-yl)phenol (35889)
4.2. Materials
4.3. Cell Culture
4.4. In Vitro Cytotoxicity
4.5. Apoptosis Assay
4.6. Western Blot Analysis
4.7. Immunocytochemistry
4.8. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine Serum Albumin |
| CSC | Cancer Stem Cell |
| FBS | Fetal Bovine Serum |
| DMSO | Dimethyl Sulfoxide |
| ECL | Enhanced Chemiluminescence |
| ICC | Immunocytochemistry |
| LRP6 | Low-density Lipoprotein Receptor-Related Protein 6 |
| SDS-PAGE | Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis |
| TCF/LEF | T-cell factor/Lef-1 Family of Transcription Factors |
| TNBC | Triple-Negative Breast Cancer |
References
- American Cancer Society, Inc. Breast Cancer Facts & Figure 2017–2018; American Cancer Society, Inc.: Atlanta, GA, USA, 2017. [Google Scholar]
- Perou, C.M. Molecular Stratification of Triple-Negative Breast Cancers. Oncologist 2011, 16, 61–70. [Google Scholar] [CrossRef] [PubMed]
- O’toole, S.A.; Beith, J.M.; Millar, E.K.; West, R.; Mclean, A.; Cazet, A.; Swarbrick, A.; Oakes, S.R. Therapeutic Targets in Triple Negative Breast Cancer. J. Clin. Pathol. 2013, 66, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-Negative Breast Cancer: Clinical Features and Patterns of Recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [PubMed]
- Bilir, B.; Kucuk, O.; Moreno, C.S. Wnt Signaling Blockage inhibits Cell Proliferation and Migration and induces Apoptosis in Triple-Negative Breast Cancer Cells. J. Transl. Med. 2013, 11, 280. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.; Barwick, B.G.; Moreno, C.S.; Ordanic-Kodani, M.; Chen, Z.; Oprea-Ilies, G.; Tang, W.; Catzavelos, C.; Kerstann, K.F.; Sledge, G.W., Jr.; et al. Wnt Signaling in Triple Negative Breast Cancer Is Associated with Metastasis. BMC Cancer 2013, 13, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geyer, F.C.; Lacroix-Triki, M.; Savage, K.; Arnedos, M.; Lambros, M.B.; Mackay, A.; Natrajan, R.; Reis-Filho, J.S. Beta-Catenin Pathway Activation in Breast Cancer Is Associated with Triple-Negative Phenotype but Not with Ctnnb1 Mutation. Mod. Pathol. 2011, 24, 209–231. [Google Scholar] [CrossRef] [PubMed]
- Maubant, S.; Tesson, B.; Maire, V.; Ye, M.; Rigaill, G.; Gentien, D.; Cruzalegui, F.; Tucker, G.C.; Roman-Roman, S.; Dubois, T. Transcriptome Analysis of Wnt3a-Treated Triple-Negative Breast Cancer Cells. PLoS ONE 2015, 10, E0122333. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/Beta-Catenin Signaling, Disease and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Shtutman, M.; Zhurinsky, J.; Simcha, I.; Albanese, C.; D’amico, M.; Pestell, R.; Ben-Ze’ev, A. The Cyclin D1 Gene Is A Target of The Beta-Catenin/Lef-1 Pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 5522–5527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Otevrel, T.; Gao, Z.; Gao, Z.; Ehrlich, S.M.; Fields, J.Z.; Boman, B.M. Evidence That Apc Regulates Survivin Expression: A Possible Mechanism Contributing to the Stem Cell Origin of Colon Cancer. Cancer Res. 2001, 61, 8664–8667. [Google Scholar] [PubMed]
- Macdonald, B.T.; Tamai, K.; He, X. Wnt/Beta-Catenin Signaling: Components, Mechanisms and Diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Prosperi, J.R.; Choudhury, N.; Olopade, O.I.; Goss, K.H. Beta-Catenin Is Required for the Tumorigenic Behavior of Triple-Negative Breast Cancer Cells. PLoS ONE 2015, 10, E0117097. [Google Scholar]
- Yin, S.; Xu, L.; Bonfil, R.D.; Banerjee, S.; Sarkar, F.H.; Sethi, S.; Reddy, K.B. Tumor-initiating Cells and Fzd8 Play a Major Role in Drug Resistance in Triple-Negative Breast Cancer. Mol. Cancer Ther. 2013, 12, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, X.; Wang, Y.; Zhang, K.; Wu, J.; Yuan, Y.C.; Deng, X.; Chen, L.; Kim, C.C.; Lau, S.; et al. Fzd7 Has a Critical Role in Cell Proliferation in Triple Negative Breast Cancer. Oncogene 2011, 30, 4437–4446. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Prior, J.; Piwnica-Worms, D.; Bu, G. Lrp6 Overexpression Defines a Class of Breast Cancer Subtype and Is a Target for Therapy. Proc. Natl. Acad. Sci. USA 2010, 107, 5136–5141. [Google Scholar] [CrossRef] [PubMed]
- Bafico, A.; Liu, G.; Goldin, L.; Harris, V.; Aaronson, S.A. An Autocrine Mechanism for Constitutive Wnt Pathway Activation in Human Cancer Cells. Cancer Cell 2004, 6, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Schlange, T.; Matsuda, Y.; Lienhard, S.; Huber, A.; Hynes, N.E. Autocrine Wnt Signaling Contributes to Breast Cancer Cell Proliferation Via the Canonical Wnt Pathway and Egfr Transactivation. Breast Cancer Res. 2007, 9, R63. [Google Scholar] [CrossRef] [PubMed]
- Green, J.L.; La, J.; Yum, K.W.; Desai, P.; Rodewald, L.W.; Zhang, X.; Leblanc, M.; Nusse, R.; Lewis, M.T.; Wahl, G.M. Paracrine Wnt Signaling both Promotes and inhibits Human Breast Tumor Growth. Proc. Natl. Acad. Sci. USA 2013, 110, 6991–6996. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lu, W.; Chen, D.; Xu, B.; Li, Y. Role of Wnt Co-Receptor Lrp6 in Triple Negative Breast Cancer Cell Migration and invasion. J. Cell Biochem. 2017, 118, 2968–2976. [Google Scholar] [CrossRef] [PubMed]
- Kahn, M. Can We Safely Target the Wnt Pathway? Nat. Rev. Drug Discov. 2014, 13, 513–532. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sutter, C.; Parker, D.S.; Blauwkamp, T.; Fang, M.; Cadigan, K.M. Cbp/P300 Are Bimodal Regulators of Wnt Signaling. EMBO J. 2007, 26, 2284–2294. [Google Scholar] [CrossRef] [PubMed]
- Takada, R.; Satomi, Y.; Kurata, T.; Ueno, N.; Norioka, S.; Kondoh, H.; Takao, T.; Takada, S. Monounsaturated Fatty Acid Modification of Wnt Protein: Its Role in Wnt Secretion. Dev. Cell 2006, 11, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Takebe, N.; Miele, L.; Harris, P.J.; Jeong, W.; Bando, H.; Kahn, M.; Yang, S.X.; Ivy, S.P. Targeting Notch, Hedgehog and Wnt Pathways in Cancer Stem Cells: Clinical Update. Nat. Rev. Clin. Oncol. 2015, 12, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.B.; Hong, I.S.; Kim, R.J.; Lee, S.Y.; Park, S.J.; Lee, E.S.; Park, J.H.; Yun, C.H.; Chung, J.U.; Lee, K.J.; et al. Wnt/Beta-Catenin Small-Molecule Inhibitor Cwp232228 Preferentially Inhibits the Growth of Breast Cancer Stem-Like Cells. Cancer Res. 2015, 75, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase inhibition Stabilizes Axin and Antagonizes Wnt Signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Dodge, M.E.; Tang, W.; Lu, J.; Ma, Z.; Fan, C.W.; Wei, S.; Hao, W.; Kilgore, J.; Williams, N.S.; et al. Small Molecule-Mediated Disruption of Wnt-Dependent Signaling in Tissue Regeneration and Cancer. Nat. Chem. Biol. 2009, 5, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective Identification of Tumorigenic Breast Cancer Cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Ponti, D.; Costa, A.; Zaffaroni, N.; Pratesi, G.; Petrangolini, G.; Coradini, D.; Pilotti, S.; Pierotti, M.A.; Daidone, M.G. Isolation and in vitro Propagation of Tumorigenic Breast Cancer Cells with Stem/Progenitor Cell Properties. Cancer Res. 2005, 65, 5506–5511. [Google Scholar] [CrossRef] [PubMed]
- Khramtsov, A.I.; Khramtsova, G.F.; Tretiakova, M.; Huo, D.; Olopade, O.I.; Goss, K.H. Wnt/Beta-Catenin Pathway Activation Is Enriched in Basal-Like Breast Cancers and Predicts Poor Outcome. Am. J. Pathol. 2010, 176, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.B.; Kim, J.Y.; Cho, S.D.; Park, K.S.; Jung, J.Y.; Lee, H.Y.; Hong, I.S.; Nam, J.S. Blockade of Wnt/Beta-Catenin Signaling Suppresses Breast Cancer Metastasis by inhibiting Csc-Like Phenotype. Sci. Rep. 2015, 5, 12465. [Google Scholar] [CrossRef] [PubMed]
- Kim do, Y.; Park, E.Y.; Chang, E.; Kang, H.G.; Koo, Y.; Lee, E.J.; Ko, J.Y.; Kong, H.K.; Chun, K.H.; Park, J.H. A Novel Mir-34a Target, Protein Kinase D1, Stimulates Cancer Stemness and Drug Resistance through Gsk3/Beta-Catenin Signaling in Breast Cancer. Oncotarget 2016, 7, 14791–14802. [Google Scholar] [PubMed]
- Satoh, K.; Zhang, L.; Zhang, Y.; Chelluri, R.; Boufraqech, M.; Nilubol, N.; Patel, D.; Shen, M.; Kebebew, E. Identification of Niclosamide as a Novel Anticancer Agent for Adrenocortical Carcinoma. Clin. Cancer Res. 2016, 22, 3458–3466. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, J.; Lu, J.; Bond, M.C.; Ren, X.R.; Lyerly, H.K.; Barak, L.S.; Chen, W. The Anti-Helminthic Niclosamide inhibits Wnt/Frizzled1 Signaling. Biochemistry 2009, 48, 10267–10274. [Google Scholar] [CrossRef] [PubMed]
- Osada, T.; Chen, M.; Yang, X.Y.; Spasojevic, I.; Vandeusen, J.B.; Hsu, D.; Clary, B.M.; Clay, T.M.; Chen, W.; Morse, M.A.; et al. Antihelminth Compound Niclosamide Downregulates Wnt Signaling and Elicits Antitumor Responses in Tumors with Activating Apc Mutations. Cancer Res. 2011, 71, 4172–4182. [Google Scholar] [CrossRef] [PubMed]
- Sack, U.; Walther, W.; Scudiero, D.; Selby, M.; Kobelt, D.; Lemm, M.; Fichtner, I.; Schlag, P.M.; Shoemaker, R.H.; Stein, U. Novel Effect of Antihelminthic Niclosamide on S100a4-Mediated Metastatic Progression in Colon Cancer. J. Natl. Cancer Inst. 2011, 103, 1018–1036. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Lin, C.; Roberts, M.J.; Waud, W.R.; Piazza, G.A.; Li, Y. Niclosamide Suppresses Cancer Cell Growth by inducing Wnt Co-Receptor Lrp6 Degradation and inhibiting the Wnt/Beta-Catenin Pathway. PLoS ONE 2011, 6, e29290. [Google Scholar] [CrossRef] [PubMed]
- andrews, P.; Thyssen, J.; Lorke, D. The Biology and Toxicology of Molluscicides, Bayluscide. Pharmacol. Ther. 1982, 19, 245–295. [Google Scholar] [CrossRef]
- Fairweather, I.; Boray, J.C. Fasciolicides: Efficacy, Actions, Resistance and Its Management. Vet. J. 1999, 158, 81–112. [Google Scholar] [CrossRef] [PubMed]
- Mook, R.A., Jr.; Ren, X.R.; Wang, J.; Piao, H.; Barak, L.S.; Kim Lyerly, H.; Chen, W. Benzimidazole inhibitors from the Niclosamide Chemotype inhibit Wnt/Beta-Catenin Signaling with Selectivity over Effects on Atp Homeostasis. Bioorg. Med. Chem. 2017, 25, 1804–1816. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, B.D.; Diering, G.H.; Bidinosti, M.A.; Dalal, K.; Alain, T.; Balgi, A.D.; Forestieri, R.; Nodwell, M.; Rajadurai, C.V.; Gunaratnam, C.; et al. Structure-Activity Analysis of Niclosamide Reveals Potential Role for Cytoplasmic Ph in Control of Mammalian Target of Rapamycin Complex 1 (Mtorc1) Signaling. J. Biol. Chem. 2012, 287, 17530–17545. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Duan, L.; He, Q.; Zhang, Z.; Zhou, Y.; Wu, D.; Pan, J.; Pei, D.; Ding, K. Identification of Niclosamide As A New Small-Molecule inhibitor of the Stat3 Signaling Pathway. ACS Med. Chem. Lett. 2010, 1, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lou, W.; Armstrong, C.; Zhu, Y.; Evans, C.P.; Gao, A.C. Niclosamide Suppresses Cell Migration and invasion in Enzalutamide Resistant Prostate Cancer Cells via Stat3-Ar Axis Inhibition. Prostate 2015, 75, 1341–1353. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Li, R.; Park, D.; Xie, M.; Sica, G.L.; Cao, Y.; Xiao, Z.Q.; Deng, X. Disruption of Stat3 by Niclosamide Reverses Radioresistance of Human Lung Cancer. Mol. Cancer Ther. 2014, 13, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.M.; Ku, H.H.; Liang, Y.C.; Chen, Y.C.; Hwu, Y.M.; Yeh, T.S. The Autonomous Notch Signal Pathway Is Activated by Baicalin and Baicalein but Is Suppressed by Niclosamide in K562 Cells. J. Cell Biochem. 2009, 106, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Suliman, M.A.; Zhang, Z.; Na, H.; Ribeiro, A.L.; Zhang, Y.; Niang, B.; Hamid, A.S.; Zhang, H.; Xu, L.; Zuo, Y. Niclosamide inhibits Colon Cancer Progression through Downregulation of the Notch Pathway and Upregulation of the Tumor Suppressor Mir-200 Family. Int. J. Mol. Med. 2016, 38, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Londono-Joshi, A.I.; Arend, R.C.; Aristizabal, L.; Lu, W.; Samant, R.S.; Metge, B.J.; Hidalgo, B.; Grizzle, W.E.; Conner, M.; Forero-Torres, A.; et al. Effect of Niclosamide on Basal-Like Breast Cancers. Mol. Cancer Ther. 2014, 13, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.; Liu, F.; Li, L.; Ding, C.; Chen, K.; Sun, Q.; Shen, Z.; Tan, Y.; Tan, C.; Jiang, Y. A Benzimidazole Derivative Exhibiting Antitumor Activity Blocks Egfr and Her2 Activity and Upregulates Dr5 in Breast Cancer Cells. Cell Death Dis. 2015, 6, E1686. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Bouska, J.J.; Ellis, P.A.; Johnson, E.F.; Leverson, J.; Liu, X.; Marcotte, P.A.; Olson, A.M.; Osterling, D.J.; Przytulinska, M.; et al. Synthesis and Evaluation of a New Generation of Orally Efficacious Benzimidazole-Based Poly(Adp-Ribose) Polymerase-1 (Parp-1) inhibitors as Anticancer Agents. J. Med. Chem. 2009, 52, 6803–6813. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, B.; Fishel, M.L.; Hurley, T.D. Selective Aldh3a1 inhibition By Benzimidazole Analogues increase Mafosfamide Sensitivity in Cancer Cells. J. Med. Chem. 2014, 57, 449–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.M.; Shah, Z.; Ahmad, V.U.; Ambreen, N.; Khan, M.; Taha, M.; Rahim, F.; Noreen, S.; Perveen, S.; Choudhary, M.I.; et al. 6-Nitrobenzimidazole Derivatives: Potential Phosphodiesterase inhibitors: Synthesis and Structure-Activity Relationship. Bioorg. Med. Chem. 2012, 20, 1521–1526. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Knowles, E.; Zardawi, S.J.; Mcneil, C.M.; Millar, E.K.; Crea, P.; Musgrove, E.A.; Sutherland, R.L.; O’toole, S.A. Cytoplasmic Localization of Beta-Catenin Is a Marker of Poor Outcome in Breast Cancer Patients. Cancer Epidemiol. Biomark. Prev. 2010, 19, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Van Noort, M.; Meeldijk, J.; Van Der Zee, R.; Destree, O.; Clevers, H. Wnt Signaling Controls the Phosphorylation Status of Beta-Catenin. J. Biol. Chem. 2002, 277, 17901–17905. [Google Scholar] [CrossRef] [PubMed]
- Zammarchi, F.; De Stanchina, E.; Bournazou, E.; Supakorndej, T.; Martires, K.; Riedel, E.; Corben, A.D.; Bromberg, J.F.; Cartegni, L. Antitumorigenic Potential of Stat3 Alternative Splicing Modulation. Proc. Natl. Acad. Sci. USA 2011, 108, 17779–17784. [Google Scholar] [CrossRef] [PubMed]
- Helfman, D.M. Niclosamide: An Established Antihelminthic Drug as a Potential Therapy against S100a4-Mediated Metastatic Colon Tumors. J. Natl. Cancer Inst. 2011, 103, 991–992. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Chao, T.K.; Chang, C.C.; Yo, Y.T.; Yu, M.H.; Lai, H.C. Drug Screening Identifies Niclosamide as an Inhibitor of Breast Cancer Stem-Like Cells. PLoS ONE 2013, 8, E74538. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Xiong, Y.; Yan, Y.; Xia, Y.; Song, X.; Liu, L.; Li, D.; Wang, N.; Zhang, L.; Zhu, Y.; et al. The Anthelmintic Drug Niclosamide induces Apoptosis, Impairs Metastasis and Reduces Immunosuppressive Cells in Breast Cancer Model. PLoS ONE 2014, 9, E85887. [Google Scholar] [CrossRef] [PubMed]
- Khanim, F.L.; Merrick, B.A.; Giles, H.V.; Jankute, M.; Jackson, J.B.; Giles, L.J.; Birtwistle, J.; Bunce, C.M.; Drayson, M.T. Redeployment-Based Drug Screening Identifies the Anti-Helminthic Niclosamide as Anti-Myeloma Therapy That also Reduces Free Light Chain Production. Blood Cancer J. 2011, 1, E39. [Google Scholar] [CrossRef] [PubMed]
- Giri, A.K.; Adhikari, N.; Khan, K.A. Comparative Genotoxicity of Six Salicylic Acid Derivatives in Bone Marrow Cells of Mice. Mutat. Res. 1996, 370, 1–9. [Google Scholar] [CrossRef]
- Ostrosky-Wegman, P.; Garcia, G.; Montero, R.; Perez Romero, B.; Alvarez Chacon, R.; Cortinas De Nava, C. Susceptibility to Genotoxic Effects of Niclosamide in Human Peripheral Lymphocytes Exposed in vitro and in vivo. Mutat. Res. 1986, 173, 81–87. [Google Scholar] [CrossRef]
- Vega, S.G.; Guzman, P.; Garcia, L.; Espinosa, J.; Cortinas De Nava, C. Sperm Shape Abnormality and Urine Mutagenicity in Mice Treated with Niclosamide. Mutat. Res. 1988, 204, 269–276. [Google Scholar] [CrossRef]
- Lacasse, E.C.; Baird, S.; Korneluk, R.G.; Mackenzie, A.E. The inhibitors of Apoptosis (Iaps) and Their Emerging Role in Cancer. Oncogene 1998, 17, 3247–3259. [Google Scholar] [CrossRef] [PubMed]
- Deveraux, Q.L.; Reed, J.C. Iap Family Proteins--Suppressors of Apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Sung, B.J.; Cho, Y.S.; Kim, H.J.; Ha, N.C.; Hwang, J.I.; Chung, C.W.; Jung, Y.K.; Oh, B.H. An Anti-Apoptotic Protein Human Survivin Is a Direct inhibitor of Caspase-3 and -7. Biochemistry 2001, 40, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.S.; Grossman, D.; Plescia, J.; Li, F.; Zhang, H.; Villa, A.; Tognin, S.; Marchisio, P.C.; Altieri, D.C. Regulation of Apoptosis at Cell Division by P34cdc2 Phosphorylation of Survivin. Proc. Natl. Acad. Sci. USA 2000, 97, 13103–13107. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C. The Survivin Saga Goes in vivo. J. Clin. Investig. 2001, 108, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.; Papanikolaou, A. Cyclin D1 in Breast Cancer Pathogenesis. J. Clin. Oncol. 2005, 23, 4215–4224. [Google Scholar] [CrossRef] [PubMed]
- Walters Haygood, C.L.; Arend, R.C.; Gangrade, A.; Li, P.K.; Hidalgo, B.; Straughn, J.M., Jr.; Buchsbaum, D.J. Niclosamide Analogs for Treatment of Ovarian Cancer. Int. J. Gynecol. Cancer 2015. [Google Scholar] [CrossRef] [PubMed]
- Arend, R.C.; Londono-Joshi, A.I.; Gangrade, A.; Katre, A.A.; Kurpad, C.; Li, Y.; Samant, R.S.; Li, P.K.; Landen, C.N.; Yang, E.S.; et al. Niclosamide and Its Analogs Are Potent inhibitors of Wnt/Beta-Catenin, Mtor and Stat3 Signaling in Ovarian Cancer. Oncotarget 2016, 7, 86803–86815. [Google Scholar] [CrossRef] [PubMed]
- Solzak, J.P.; Atale, R.V.; Hancock, B.A.; Sinn, A.L.; Pollok, K.E.; Jones, D.R.; Radovich, M. Dual Pi3k and Wnt Pathway inhibition Is a Synergistic Combination against Triple Negative Breast Cancer. NPJ Breast Cancer 2017, 3, 17. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Choi, M.Y.; Yu, J.; Castro, J.E.; Kipps, T.J.; Carson, D.A. Salinomycin Inhibits Wnt Signaling and Selectively Induces Apoptosis in Chronic Lymphocytic Leukemia Cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13253–13257. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Clevers, H. Wnt Signalling in Stem Cells and Cancer. Nature 2005, 434, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Curtin, J.C.; Lorenzi, M.V. Drug Discovery Approaches to Target Wnt Signaling in Cancer Stem Cells. Oncotarget 2010, 1, 552–566. [Google Scholar]
- Gupta, P.B.; onder, T.T.; Jiang, G.; Tao, K.; Kuperwasser, C.; Weinberg, R.A.; Lander, E.S. Identification of Selective Inhibitors of Cancer Stem Cells by High-Throughput Screening. Cell 2009, 138, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Arend, R.C.; Londono-Joshi, A.I.; Samant, R.S.; Li, Y.; Conner, M.; Hidalgo, B.; Alvarez, R.D.; Landen, C.N.; Straughn, J.M.; Buchsbaum, D.J. Inhibition of Wnt/Beta-Catenin Pathway by Niclosamide: A Therapeutic Target for Ovarian Cancer. Gynecol. Oncol. 2014, 134, 112–120. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||
| R1 | R2 | R3 | R4 | R7 | |
| SRI32529 | H | NO2 | H | OH | Cl |
| SRI33576 | H | CF3 | H | OH | Cl |
| SRI35357 | Cl | H | Cl | OH | F |
| SRI35361 | F | Cl | H | OH | Cl |
| SRI35889 | Cl | Cl | H | OH | Cl |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gangrade, A.; Pathak, V.; Augelli-Szafran, C.E.; Wei, H.-X.; Oliver, P.; Suto, M.; Buchsbaum, D.J. Preferential Inhibition of Wnt/β-Catenin Signaling by Novel Benzimidazole Compounds in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2018, 19, 1524. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051524
Gangrade A, Pathak V, Augelli-Szafran CE, Wei H-X, Oliver P, Suto M, Buchsbaum DJ. Preferential Inhibition of Wnt/β-Catenin Signaling by Novel Benzimidazole Compounds in Triple-Negative Breast Cancer. International Journal of Molecular Sciences. 2018; 19(5):1524. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051524
Chicago/Turabian StyleGangrade, Abhishek, Vibha Pathak, Corinne E. Augelli-Szafran, Han-Xun Wei, Patsy Oliver, Mark Suto, and Donald J. Buchsbaum. 2018. "Preferential Inhibition of Wnt/β-Catenin Signaling by Novel Benzimidazole Compounds in Triple-Negative Breast Cancer" International Journal of Molecular Sciences 19, no. 5: 1524. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051524