1. Introduction
According to current statistics, more than 10% of insects in nature interact with symbiotic microorganisms [
1]. The interaction between symbiotic fungi and bark beetles has also been studied extensively [
2,
3,
4,
5,
6,
7]. The vast majority of bark beetles are closely related to symbiotic fungi at various stages of development, and some bark beetles directly use the fungal fruiting bodies or fungal hyphae colonized in the gallery as a food source [
2,
8]. A variety of symbiotic fungi, from the microecological point of view of the bark beetles, make the larvae more advantageous than other xylophagous insects. The bark beetles use intestinal microbiotas to improve the utilization of plant carbon and nitrogen nutrition, which is more difficult for insects to decompose, thereby improving the constraints of the bark beetles on overcoming the nutrient-poor factors of food sources and ensuring their development and reproduction [
9,
10,
11].
Trypophloeus klimeschi Eggers (Coleoptera, Curculionidae, Scolytinae) was first recorded in the Kyrgyz Republic, which borders Xinjiang Province in China [
12]. Following an outbreak in 2003 in Xinjiang Province,
T.
klimeschi spread rapidly to the adjacent areas. The widespread outbreak of this beetle has caused huge economic, ecological, and social losses in China’s northwest shelter forest. The insect is now found in Dunhuang, where it has been identified as
T.
klimeschi by morphology [
12,
13]. This is the first systematic survey of fungal communities across the life cycle of
T.
klimeschi. The previous research on the intestinal fungal diversity of insects was conducted mainly through traditional methods such as culture separation and morphological identification [
14]. This provides an important basis for the composition and species diversity of insect gut fungal, but it is inevitably incomplete in the description of insect gut microbes. According to statistics, approximately 99% of the microorganisms in nature are not culturable [
15], but molecular biology technology can make up for this limitation. Molecular biology methods allow for the subsequent sequencing and analysis of the DNA to characterize fungal species composition and abundance [
16].
There have been many reports on the research of intestinal micro-organisms with bark beetles, such as the intestinal microflora of some bark beetles showed differences in different geographic environment [
17,
18]. However, there are few studies on the differences in the entire development stage of the bark beetles, including difference in sexes. Recently, the symbiotic relationship between insects and their intestinal microbiota has attracted widespread attention from scholars around the world. Studies have shown that symbiotic microbes play a very important role in the invasion, settlement, spawning, development, reproduction, and other roles in developmental and life cycle stages of bark beetles [
19,
20,
21,
22]. Concurrently, a better understanding of the symbiosis formed by an insect and its colonizing microorganisms could be useful to improve insect control, use and development [
17,
23]. Through clarifying the composition of insect gut fungi, scientists can further study the role of gut fungi in the host physiology.
High throughput sequencing technology is used to study the fungal community structure and diversity dynamics at different developmental stages, different generations, and between T. klimeschi adult males and females. The results reveal the interaction between symbiotic microorganisms and T. klimeschi and provide a theoretical basis for the development of new biological control technologies.
3. Discussion
Although not often obvious to the naked eye, fungi are as deeply enmeshed in the evolutionary history and ecology of life as any other organism on Earth [
24]. Furthermore, many data are available for relevant ecological traits such as acting as decomposers by releasing extracellular enzymes to break down various plant biopolymers and using the resulting products [
25,
26], or communities of endophytic fungi containing wood-decomposer fungi that are present in a latent state prior to plant death [
24]. Investigation of the gut fungal community of bark-inhabiting insects is important to better understand the potential role of gut microorganisms in host nutrition, cellulose/hemicellulose degradation, nitrogen fixation, and detoxification processes. Additionally, microbes in the beetle’s intestine have proven to be an important source of enzymes for various industries [
27]. The authors not only conduct fungal inventories of
T. klimeschi across the full host life cycle, but also compare the differences in the community composition in different generations and each sex, which provides new insights into the metabolic potentials of Curculionidae-associated fungal communities. However, not all active fungi could be successfully detected and identified inside the host.
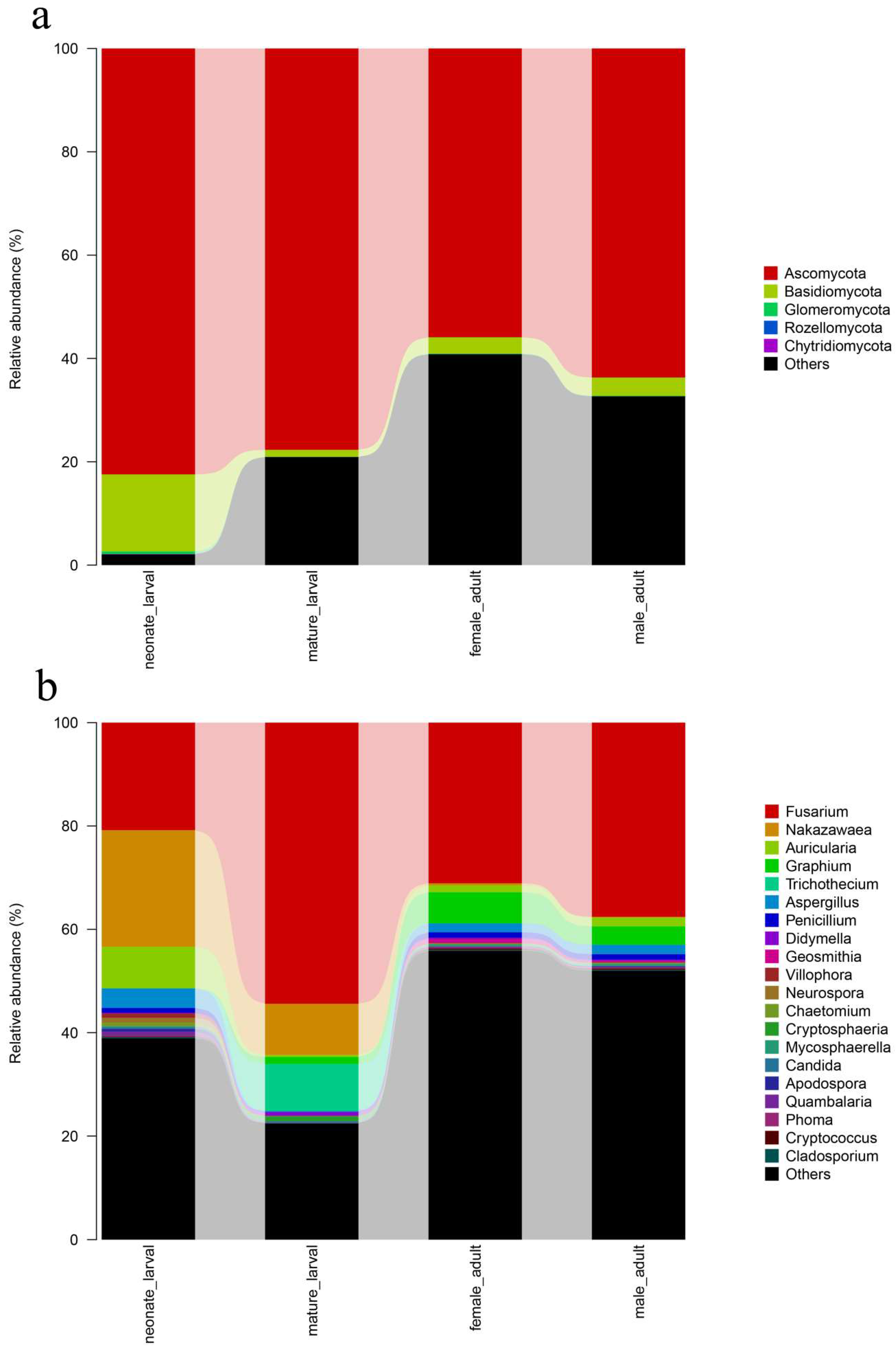
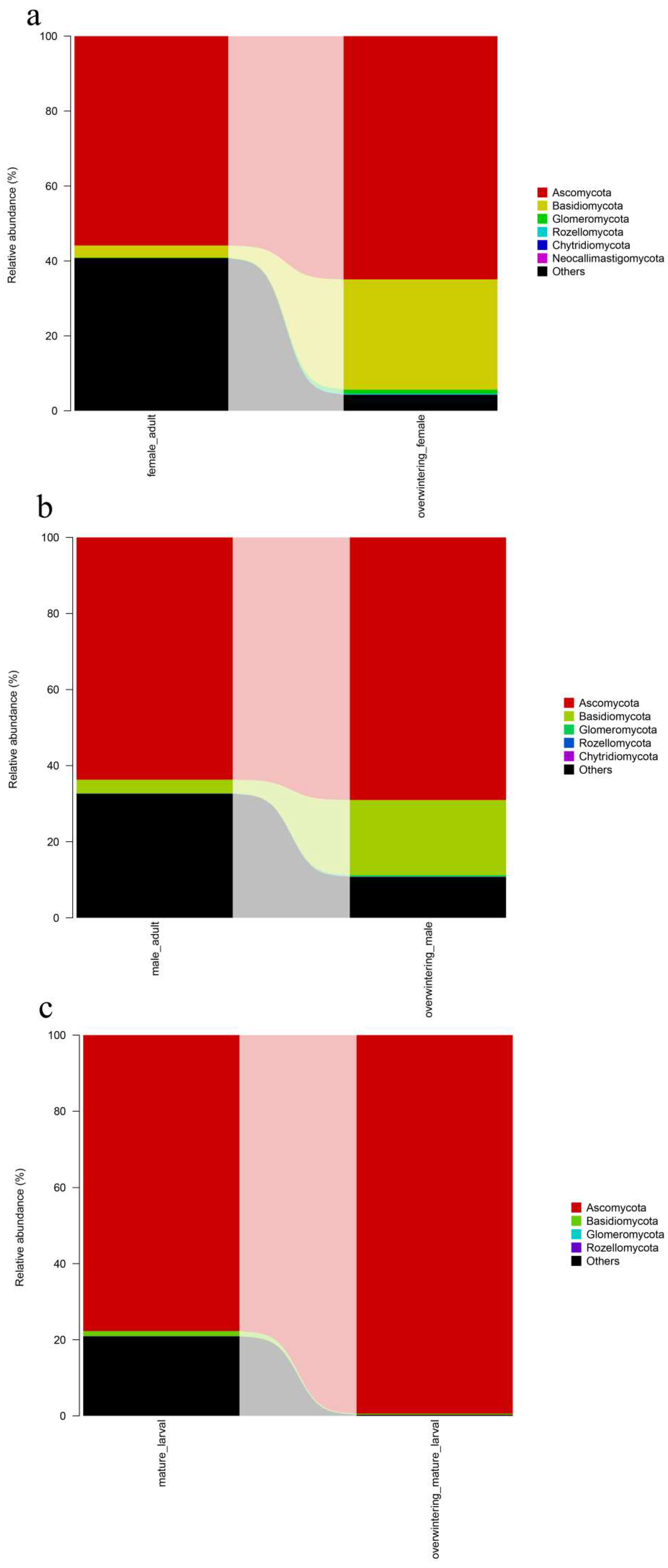
The predominant species found in the intestinal fungal communities of
T. klimeschi formed a group of low complexity, mainly belonging to the phyla Ascomycota and Basidiomycota. A low level of fungal community complexity is typical of the bark beetle gut discovered to date, except in the fungus-feeding beetles [
17,
28,
29]. The presence and high abundance of these fungal phyla have been previously reported in the gut of larvae from several Coleoptera [
30,
31,
32,
33].
However, the structure of the fungal community differed depending on the developmental stages. The fungal community in the neonate larvae was more diverse than the community identified in the mature larvae. Fungal community richness further increased in adults. Since the neonate larvae of
T. klimeschi feed on inner bark and the gallery contain almost entirely excrement, the living habits were similar to
Trypophloeus striatulus [
34]. Such behavior might contribute to maintaining gut fungal community in the neonate larvae stage. It has been documented that the prepupal larvae of
T. striatulus evacuate their gut [
34]. This phenomenon implies that gut fungal community might be re-structured in the subsequent developmental stages of the life cycle. Furthermore, micro-environments were different between larvae and adults. Moreover, this difference in taxonomic membership might reflect different functional roles across certain life stages. Some fungal taxa guide the entrance point for gallery construction, such as
Trypophloeus striatulus possibly attraction to odor emitted through lenticels that overlie susceptible
Cytospora-infested phloem [
34].
Fusarium was the most abundant genus and was conserved in all development stages. This result indicates that the conserved fungal community of shared fungal taxa should be well adapted to
T. klimeschi. It was interesting that Fusarium species (Ascomycota, Nectriaceae) are among the most diverse and widespread plant-infecting fungi, and numerous metabolites produced by Fusarium spp. are toxic to insects [
35,
36]. The
Tenebrio molitor larvae were able to use the wheat kernels that were colonized by
Fusarium proliferatumor and
Fusarium poae which produced fumonisins, enniatins, and beauvericin during kernel colonization without exhibiting increased mortality. The result suggests that
Tenebrio molitor can tolerate or metabolize those toxins. Some insect species appear to benefit from the presence of aflatoxin producers [
37,
38] or mycotoxin produced by Fusarium spp. fungi [
39]. The authors have not investigated induced reactions of Fusarium fungi to the presence or feeding of
T. klimeschi, this will be the subject of subsequent studies.
The genera Nakazawaea, Trichothecium, Aspergillus, Didymella, and Villophora which belonged to the Ascomycota; and Auricularia which belonged to Basidiomycota were more prevalent in the larvae samples.
Nakazawaea is the ascomycete yeast genus, derived from the genus Pichia [
40]. Yeasts are frequently isolated from the larvae of bark beetles [
41,
42]. Additionally,
Nakazawaea is widely distributed in nature and common to insects that bore into forest trees [
43,
44,
45]. Yeasts are commonly associated with bark beetles and might be an important nutritional source for the insect host [
28,
46]. Moreover, yeasts such as the
Candida species can assimilate nutrients such as nitrate, xylose, and cellobiose [
47]. Nutritional needs should be different over the developmental stages of the beetle, as the beetle needs nutritional benefits to accomplish different steps in its life style, such as development and ovogenesis [
48]. The high prevalence of yeasts associated with the larvae supports the hypothesis that yeasts are essential nutritional elements for the development of the
T.
klimeschi. Trichothecium has shown potential for biotransformation and enzyme production [
49]. Found in several wood-feeding Coleptera larvae, Trichothecium was the most abundant genus as well [
33]. There are few reports about Aspergillus interaction with insects; perhaps it plays the same role as the genus Fusarium. Auricularia are typical wood-inhabiting fungi in the forest ecosystem. They can degrade cellulose, hemicelluloses, and ligini of wood [
50]. Some wood-inhabiting fungi provide foods and breeding grounds for some beetles [
51,
52]. Similar to termites [
53],
T. klimeschi also feeds on plant tissue lacking nitrogen nutrients. Some studies have reported that the mycelium of Auricularia could improve the quality of termite foods and increase egg production by termites [
54].
According to this study’s results, the fungal community in the neonate larvae was more diverse than the community identified in other development stages. Considering that the digestive system of the neonate larvae was just maturing, ingesting large amounts of carbon and nitrogen nutrition associated with their symbiotic microorganisms explains why the guts of neonate larvae contained more diverse fungal communities. Overall, these fungi are all relevant to nutritional metabolism. The authors speculate that these fungal symbionts might play important roles in nutrition in T. klimeschi.
Adults harbored high proportions of Graphium, which belongs to Ascomycota. The genus Graphium is known as ‘blue stain fungi’ [
55]. When bark beetles invade conifers, the fungus taps into the sapwood nitrogen and transports it to the phloem where the beetle feeds, increasing the nitrogen content by up to 40% [
56]; this is critical for bark beetle development and survival [
2,
9,
56]. Moreover, one blue stain fungus
Leptographium qinlingensis is the pathogenic fungus carried by adults of
Dendroctonus armandi, which develops in the tissues and cells of xylem and phloem of
Pinus armandi after
D. armandi attacks healthy host trees, decomposes the secretory resin cells in resin ducts and the parenchyma cell sapwood tissues, then affects the metabolism of resin [
57]. The presence of Graphium was also observed in larvae and adult beetles of
Euwallacea fronicatus as well as in the galleries of several tree species [
33,
58]. According to the result, the authors speculate that the physiology and biochemistry resistance of
P. alba var.
pyramidalis was weakened and nutrient degradation was accelerated by the attacking of genus
Graphium. Moreover, some fungal taxa guide the entrance point for gallery construction, such as
Trypophloeus striatulus possibly attraction to odor emitted through lenticels that overlie susceptible
Cytospora-infested phloem [
34]. We speculated that
Graphium plays an important role in the invasion and colonization of
T. klimeschi.
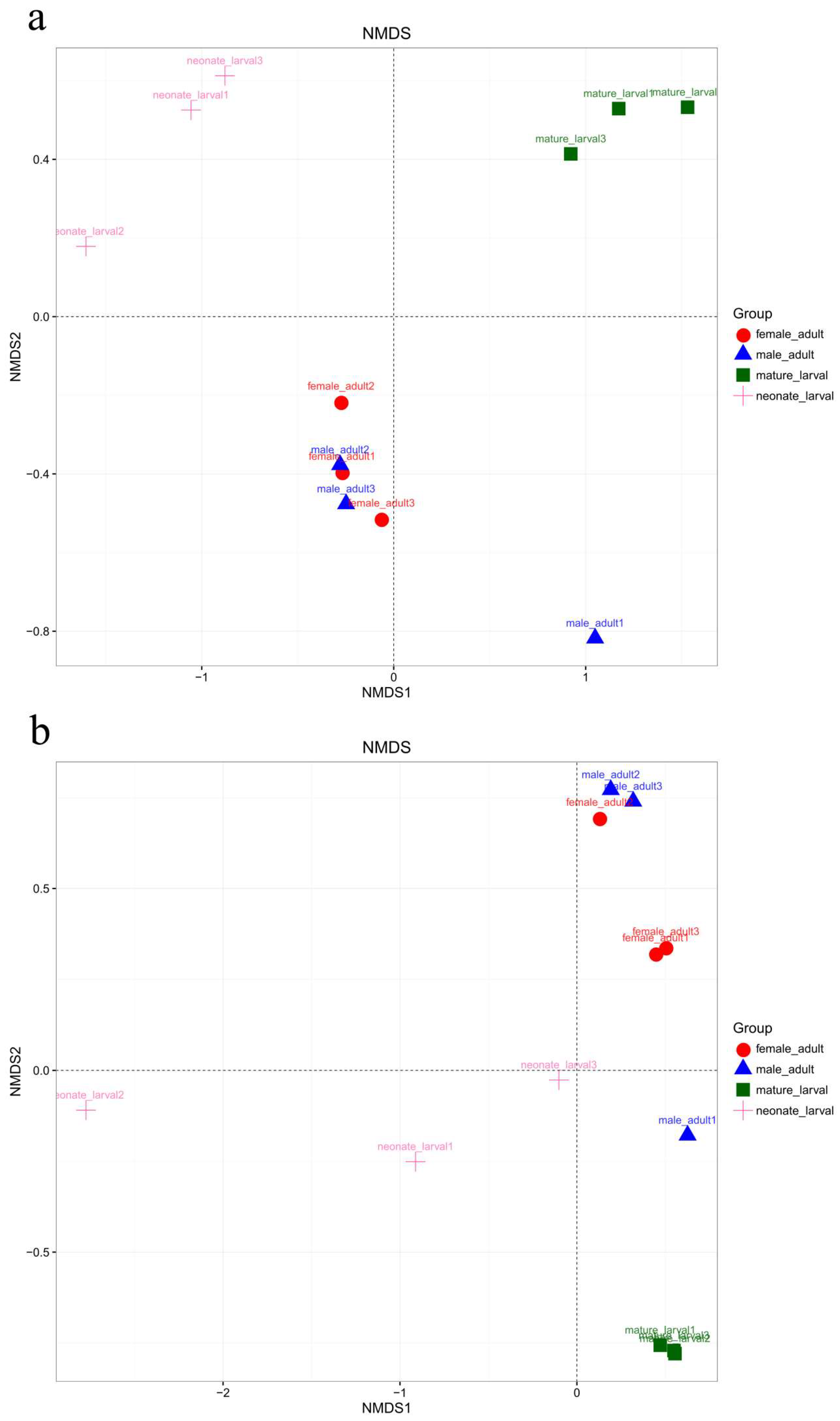
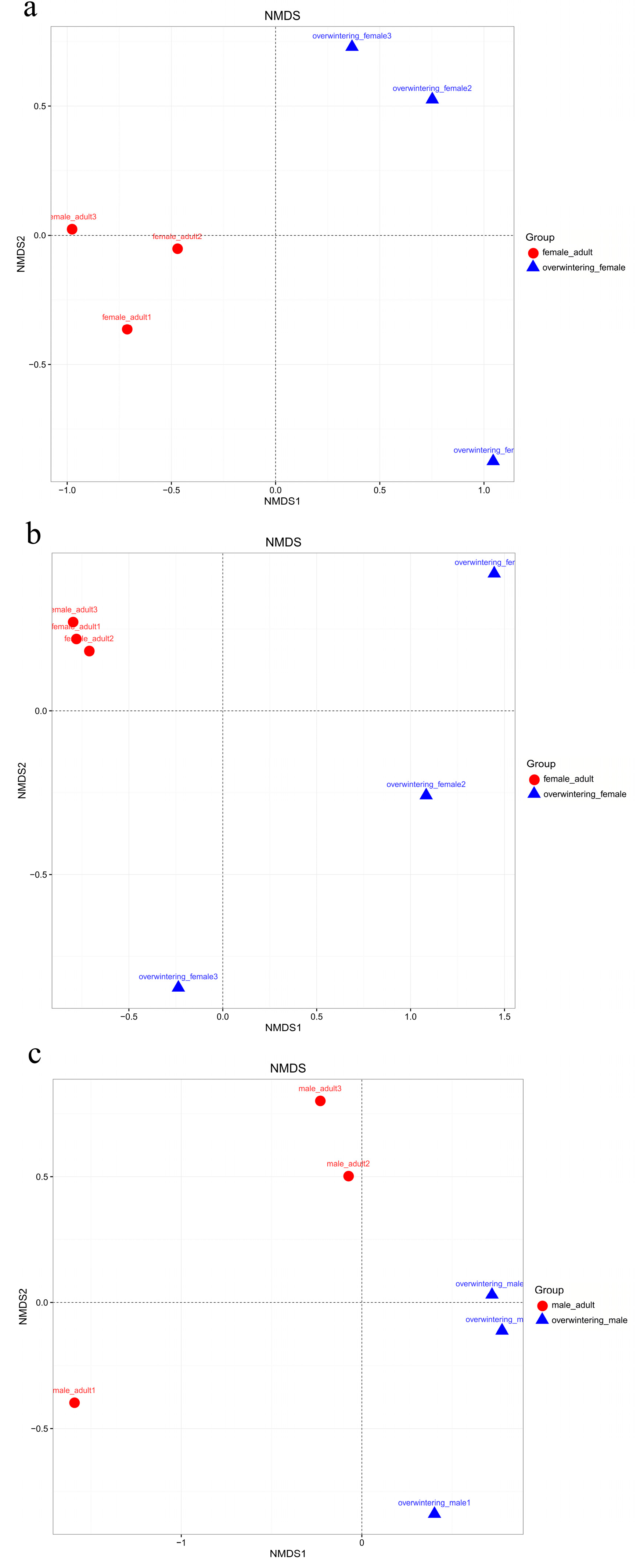
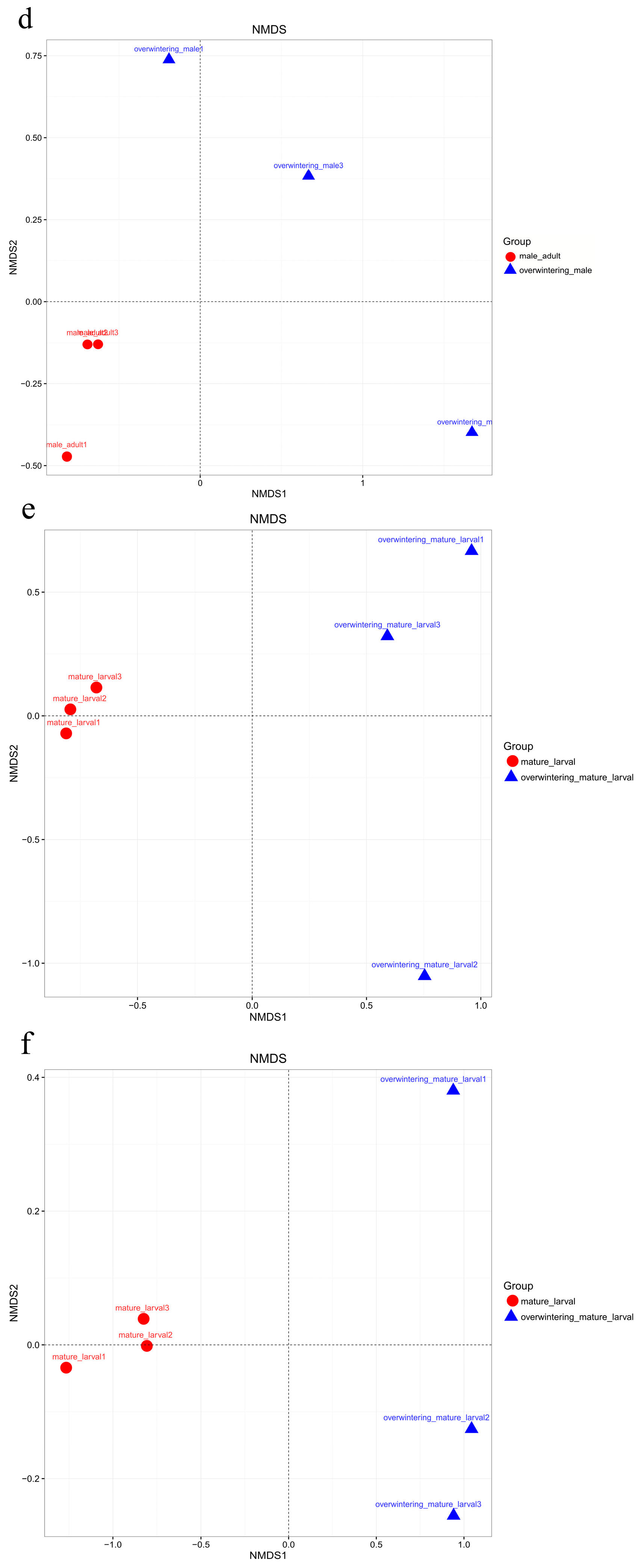
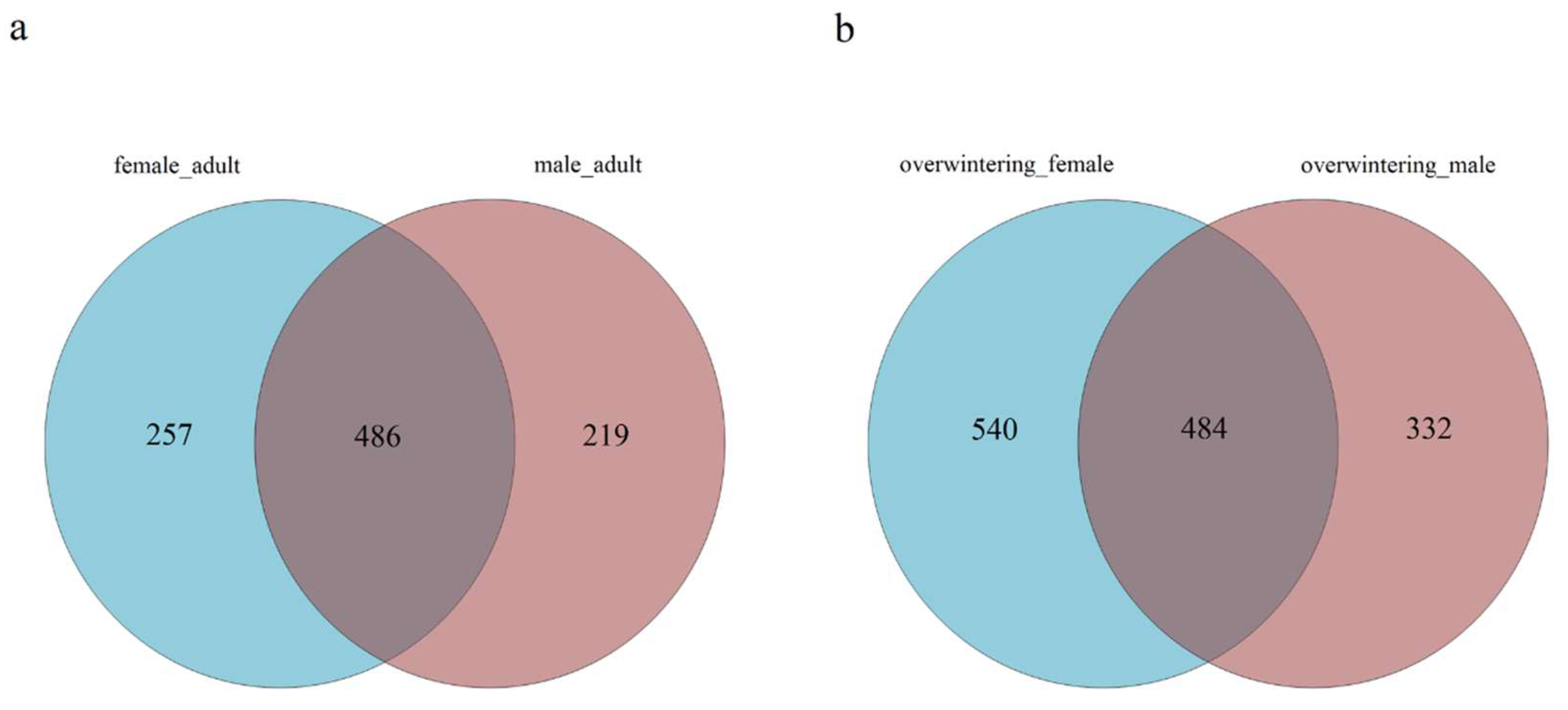
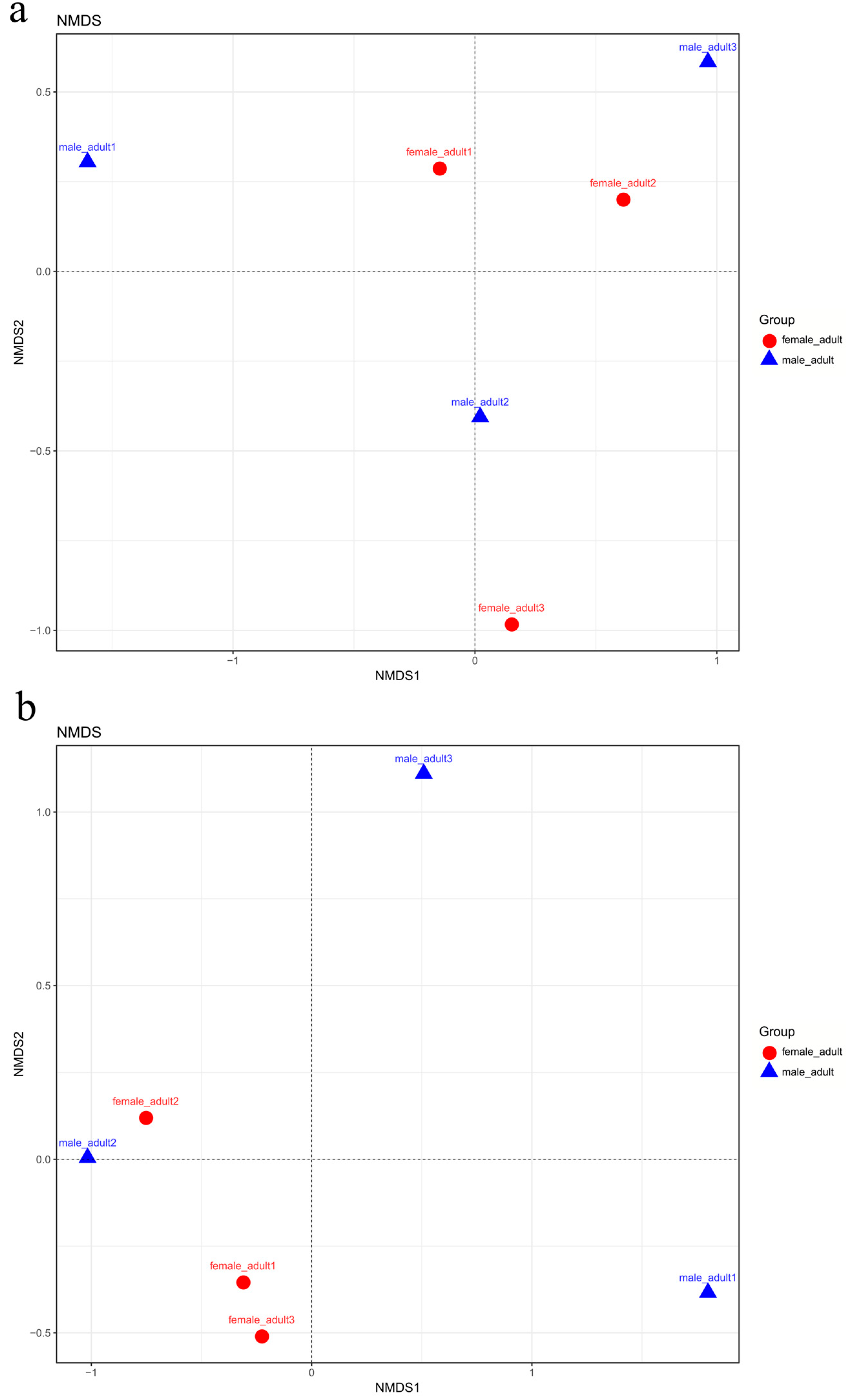
The fungal community structures found in the guts of adult females and adult males were similar, suggesting that the fungal community structure in T. klimeschi adult is conserved.
Interestingly, there was a significant difference in fungal community structure between the mature larvae and overwintering mature larvae. The pest overwintered in the form of mature larvae. Insects face great challenges in surviving at low temperatures in frigid and temperate zones [
59,
60,
61]. Insects’ cold-tolerance capacity is a dominant factor that affects their adaption to the geographical environment [
62,
63]. The present results suggested that gut fungal community compositions differed during the overwintering period. Fusarium, which belongs to the Ascomycota, were significantly increased in overwintering mature larvae. Nectriaceae only appeared and was abundant in overwintering mature larvae. We hypothesized that the two genera are associated with the insect overwintering process and resistance to low temperatures.
4. Materials and Methods
4.1. Collection Site Description
The T. klimeschi were collected from the bark of infested Populus alba var. pyramidalis at the shelter belt of Dunhuang City (40°06′50.61″ N, 94°36′10.24″ E), Gansu Province, China. Dunhuang is located in Northwestern Gansu Province, which has a temperate continental, dry climate with low rainfall, high evaporation, large temperature differences between day and night, long sunshine duration, an annual average temperature of 9.4 °C, monthly average maximum temperature of 24.9 °C (July), and monthly average minimum temperature of −9.3% °C (January), extreme maximum temperature of 43.6 °C, and minimum temperature of −28.5 °C.
4.2. Life-Cycle of T. klimeschi Description
There were two generations of
T. klimeschi per year and the pest overwintered in the form of mature larvae. There were two peak periods in a year. Mature larvae began to pupate in early May. Adults started to emerge beginning in mid-May, with a peak from late-May until mid-June. Second generation larvae pupated in mid-July. Adult emergence peaked in August.
T. klimeschi began wintering in October (see
Table A6 in
Appendix A).
4.3. Insect Collection and Dissection
According to the life history of T. klimeschi, larvae and adult females and males were collected from January 2017 to August 2017. The pest overwintered in the form of mature larvae from October to May of the following year. Due to the lowest average temperature being in January, overwintering mature larvae were collected in January 2017. Adults began to emerge in mid-May with a peak from late-May until mid-June. Overwintering adults were collected in May 2017. The neonate larvae of the first generation were collected in June 2017. The mature larvae of the first generation were collected in July 2017. The adults of the first generation were collected in August 2017. To identify gut fungal community structure succession in different development stages, the authors compared neonate larvae, mature larvae, adult females, and adult males. To identify gut fungal community structure succession in different generations, the authors compared adult females with overwintering adult females, adult males with overwintering adult males, and mature larvae with overwintering mature larvae. To identify gut fungal community structure succession in different sexes, the authors compared adult females with adult males and overwintering adult females with overwintering adult males.
The samples were collected at the laboratory in sterile vials. To investigate the influence of sex on gut-associated fungi, adult females and males were separated according to morphology (according to the morphological observation, the salient features distinguishing adult males and females are in the granules of the elytron: the male has three sharp corners on interstria 5 near the tail at the declivity of the left and right elytrons, and the females do not have this feature (relevant data have not been published). A total of 180 insect samples in each life stage were gathered for high-throughput sequencing analysis.
Insect samples were rinsed in sterile water, surface sterilized with 70% ethanol for 3 min, and then rinsed twice in sterile water. Following being placed in 10 mM sterilized phosphate-buffered saline (138 mM NaCl and 2.7 mM KCl, pH 7.4) the insects were dissected under a stereomicroscope with the aid of insect pins to excise the mid-guts and hindguts [
64]. Sixty guts were excised from each sample. The treatment in each sample was repeated three times.
4.4. DNA Extraction
The E.Z.N.A. Fungal DNA Kit (Omega Biotech, Doraville, GA, USA) was used to extract T. klimeschi samples guts fungal DNA following the instruction booklet. The gut fungal DNA was stored at −20 °C before using. DNA samples were mixed in equal concentrations, and the mixed DNA specimens were sent to Personal Biotechnology Co., Ltd. (Shanghai, China) for analysis by high throughput sequencing.
4.5. Bioinformatics and Statistical Analysis
Following sequencing, all reads were processed and analyzed using the QIIME package release v1.8.0 [
65]. Sequences were clustered by the open-reference OTU clustering using the default settings at a 97% identity threshold. The representative ITS sequences were assigned to taxonomy using the UNITE database [
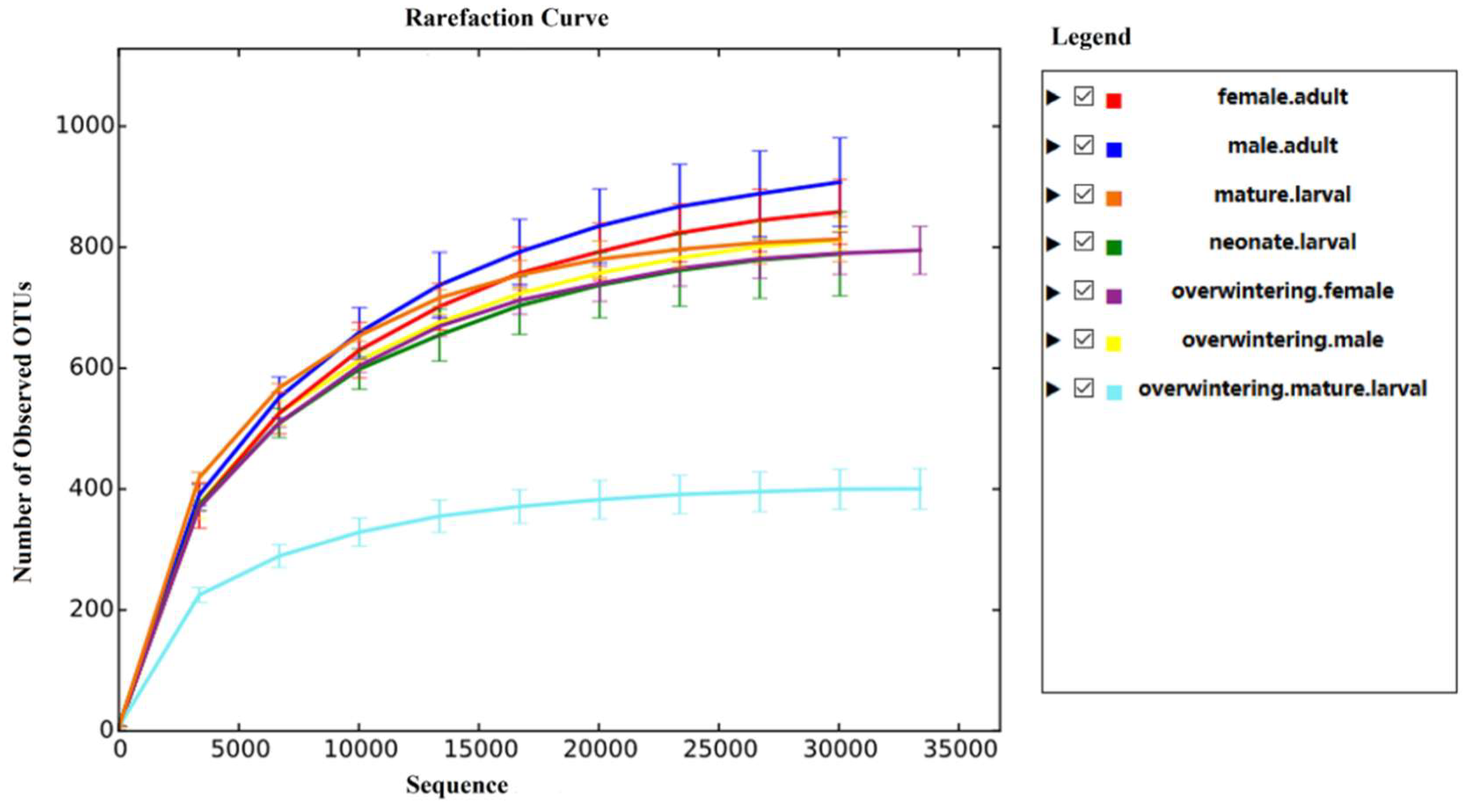
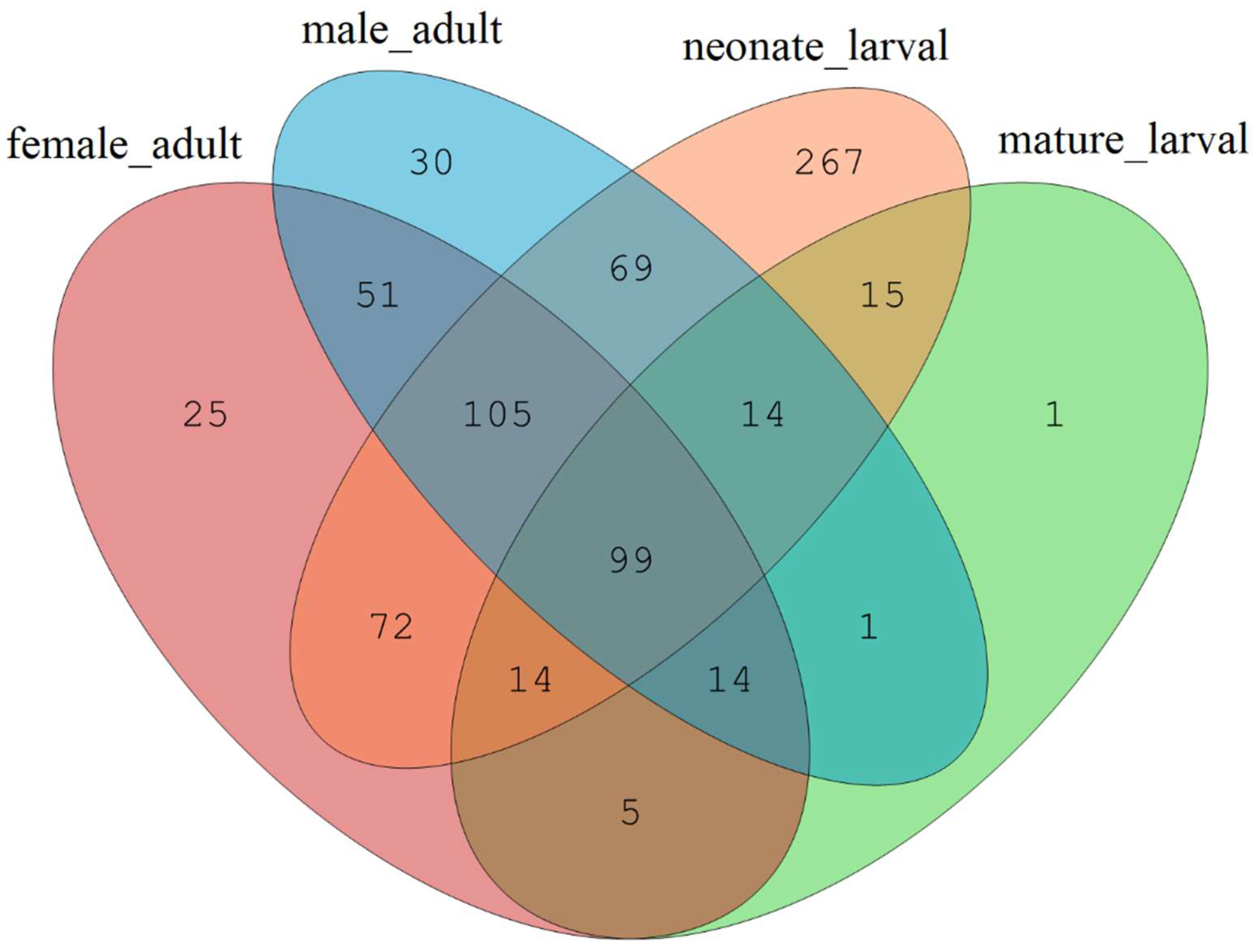
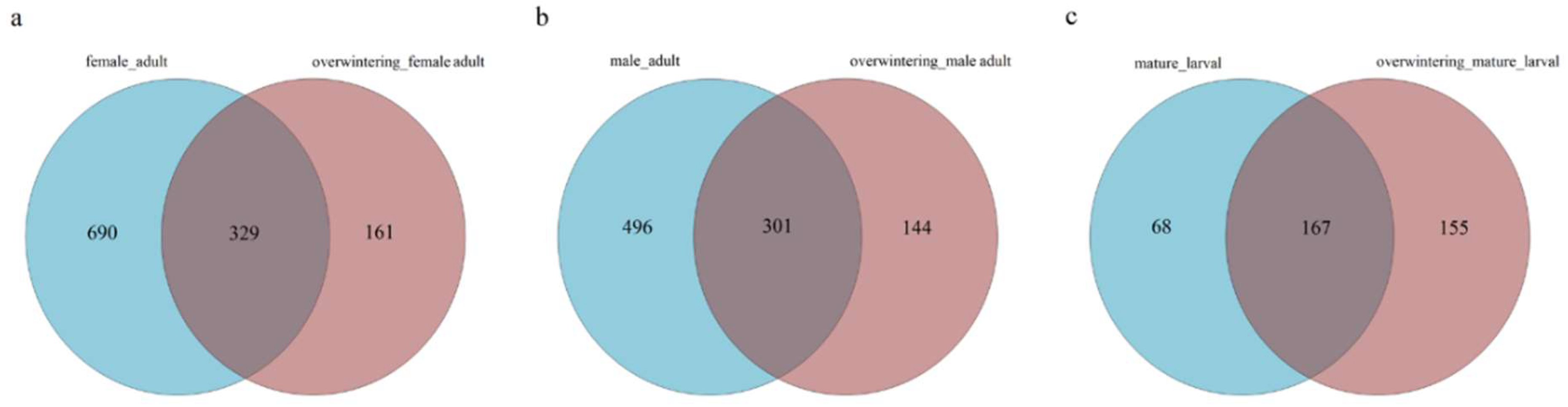
66]. The alpha diversity analysis included observed species, ACE and Chao estimators, Simpson and Shannon diversity indices estimate of coverage. Rarefaction curves were generated based on observed species. According to the OTU classification and classification status identification results, the specific composition of each sample at each classification level was obtained. Nonmetric multidimensional scaling (NMDS) analysis was conducted on the sample-OTU matrix using the Bray–Curtis distances. Additionally, Venn diagrams were also created to observe the partition of the OTUs across different samples. The data differences were analyzed by SPSS (SPSS version 20.0; SPSS, Chicago, IL, USA) software.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












