Oridonin Enhances Radiation-Induced Cell Death by Promoting DNA Damage in Non-Small Cell Lung Cancer Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
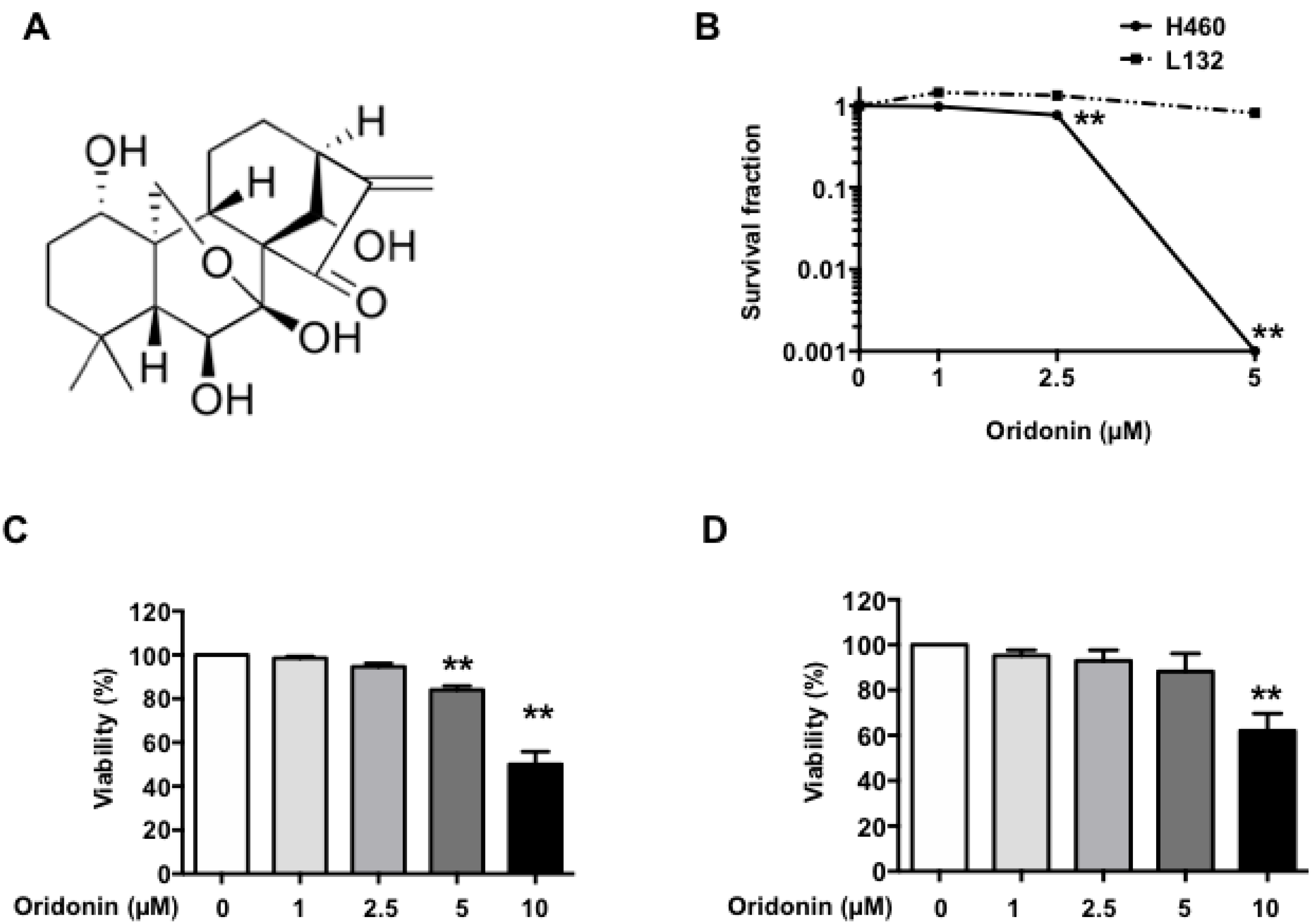
2.1. Oridonin Inhibits H460 Lung Cancer Cell Growth
2.2. Oridonin Enhances Radiation Effects on Lung Cancer Cells
2.3. Oridonin Enhances Radiation-Induced ROS Production
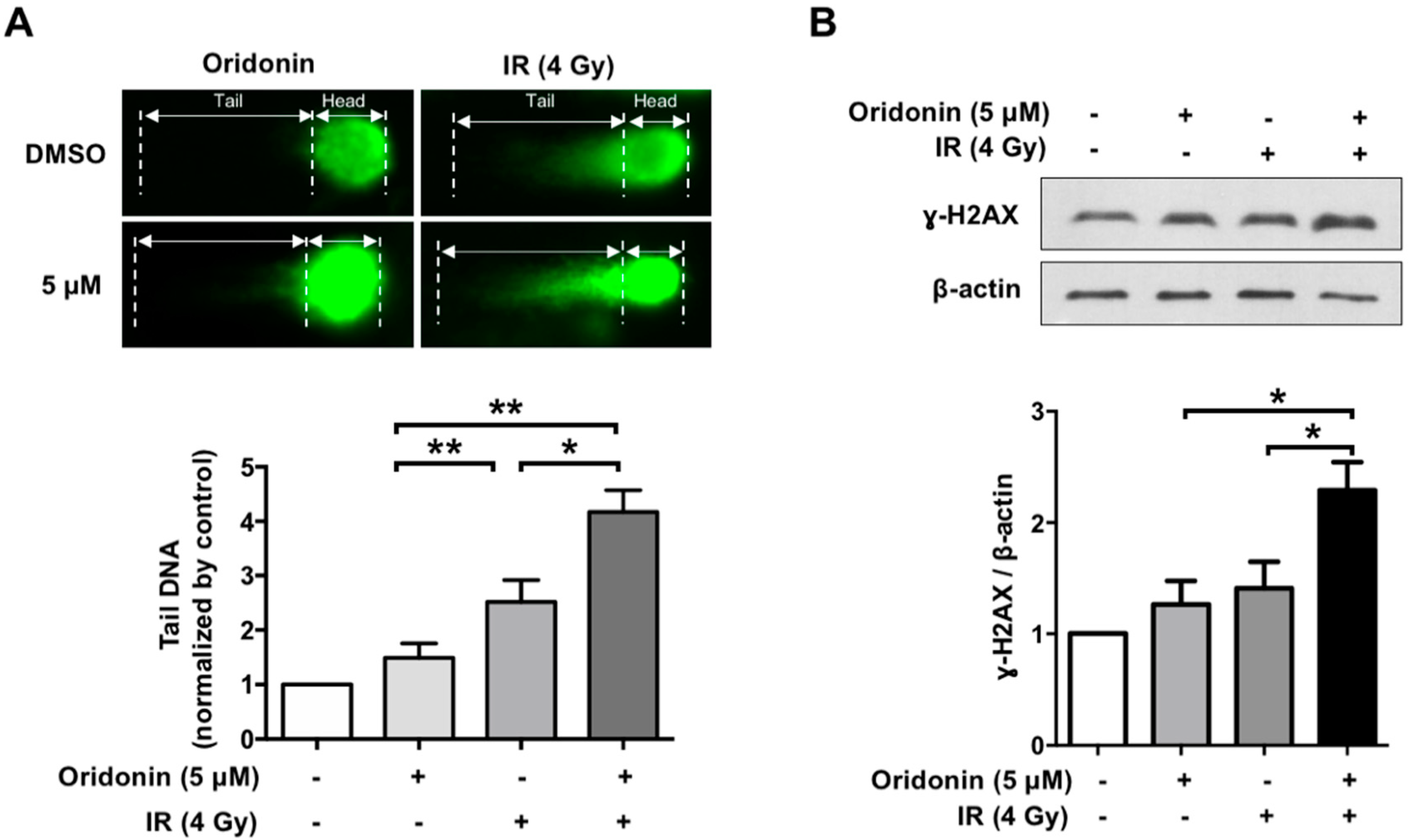
2.4. Oridonin Accelerates Radiation-Induced DNA Damage
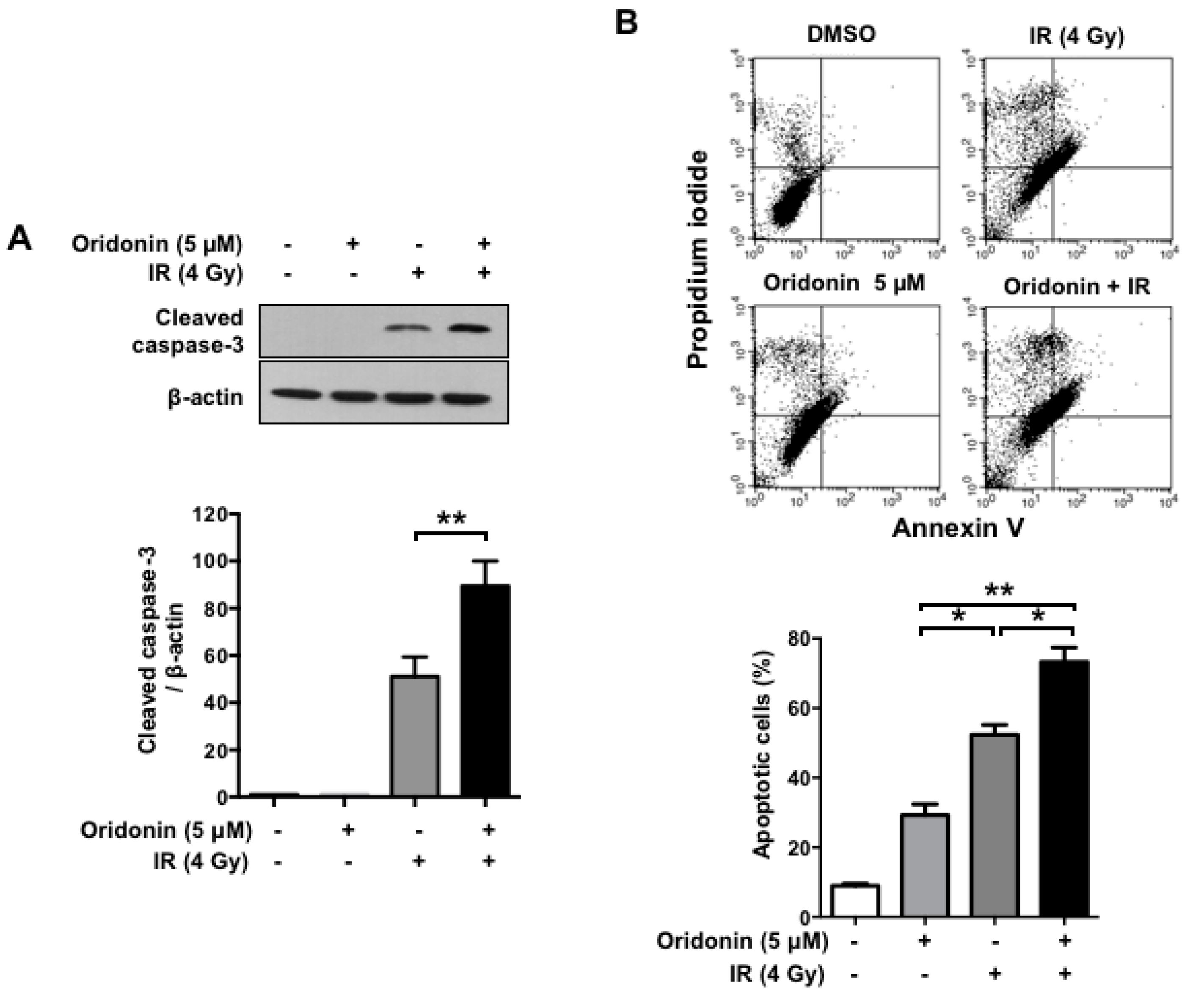
2.5. Oridonin Increases Radiation-Induced Apoptotic Cell Death
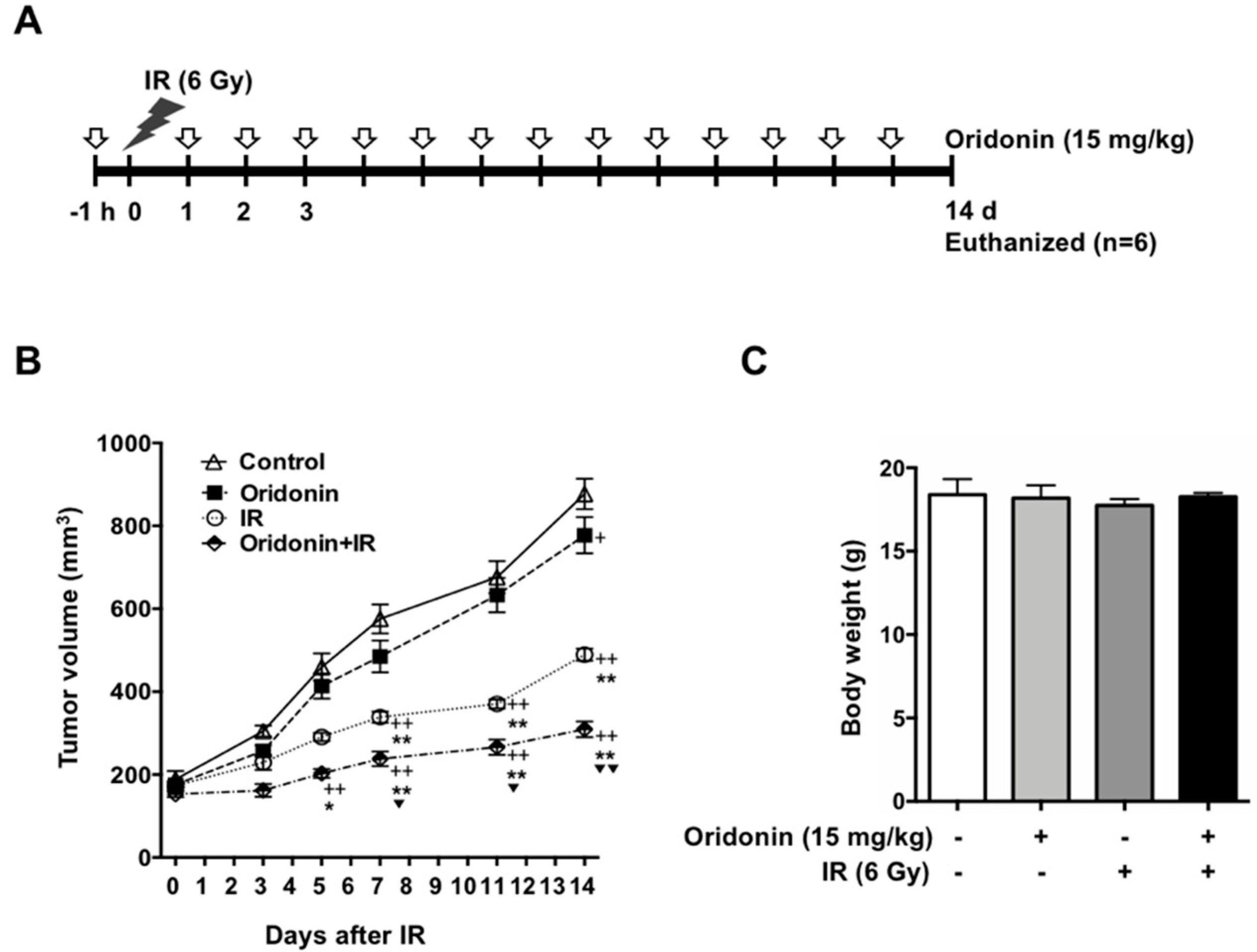
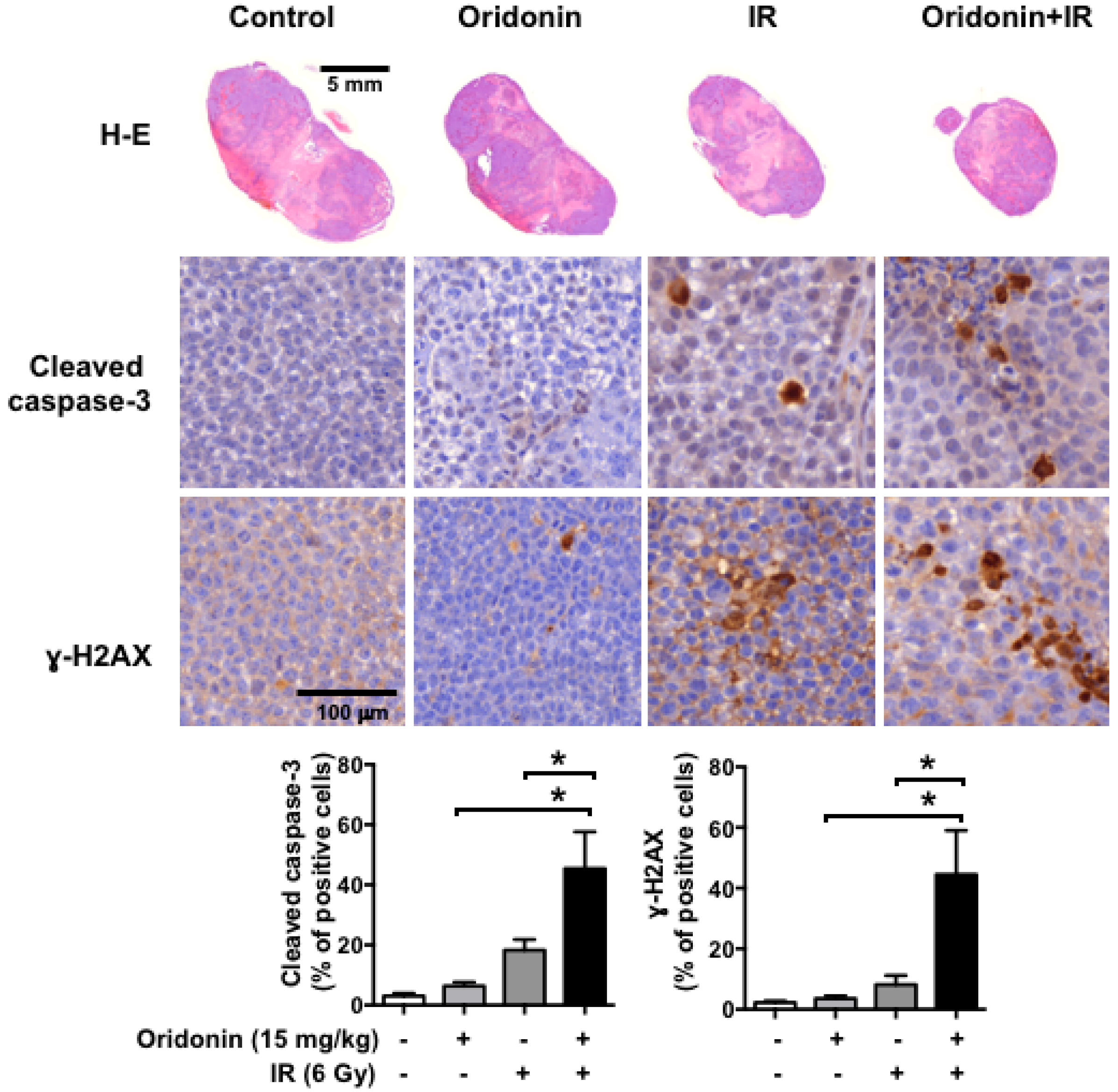
2.6. Oridonin Enhances Radiation-Induced Tumor Growth Inhibition
3. Discussion
4. Materials and Methods
4.1. Cells and Treatments
4.2. MTT Assay
4.3. Clonogenic Aassay
4.4. Determination of ROS Production
4.5. Comet Assay
4.6. Western Blots
4.7. Apoptotic Cell Death by Flow Cytometry
4.8. Animal Experiments
4.9. Immunohistochemistry and Quantification
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Maciejczyk, A.; Skrzypczyńska, I.; Janiszewska, M. Lung cancer. Radiotherapy in lung cancer: Actual methods and future trends. Rep. Pract. Oncol. Radiother. 2014, 19, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Delaney, G.; Barton, M.; Jacob, S.; Jalaludin, B. A model for decision making for the use of radiotherapy in lung cancer. Lancet Oncol. 2003, 4, 120–128. [Google Scholar] [CrossRef]
- Tyldesley, S.; Boyd, C.; Schulze, K.; Walker, H.; Mackillop, W.J. Estimating the need for radiotherapy for lung cancer: An evidence-based, epidemiologic approach. Int. J. Radiat. Oncol. Biol. Phys. 2001, 49, 973–985. [Google Scholar] [CrossRef]
- Zhang, Q.; Gao, M.; Luo, G.; Han, X.; Bao, W.; Cheng, Y.; Tian, W.; Yan, M.; Yang, G.; An, J. Enhancement of Radiation Sensitivity in Lung Cancer Cells by a Novel Small Molecule Inhibitor That Targets the β-Catenin/Tcf4 Interaction. PLoS ONE 2016, 11, e0152407. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Cox, J.D. Improving radiation conformality in the treatment of non-small cell lung cancer. Semin. Radiat. Oncol. 2010, 20, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Bogart, J.A. Fractionated radiotherapy for high-risk patients with early-stage non-small cell lung cancer. Semin. Thorac. Cardiovasc. Surg. 2010, 22, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Chougule, P.B.; Suk, S.; Chu, Q.D.; Leone, L.; Nigri, P.T.; McRae, R.; Lekas, M.; Barone, A.; Bhat, D.; Bellino, J. Cisplatin as a radiation sensitizer in the treatment of advanced head and neck cancers. Results of a phase II study. Cancer 1994, 74, 1927–1932. [Google Scholar] [CrossRef] [Green Version]
- Tishler, R.B.; Schiff, P.B.; Geard, C.R.; Hall, E.J. Taxol: A novel radiation sensitizer. Radiat. Oncol. Biol. 1992, 22, 613–617. [Google Scholar] [CrossRef]
- Yang, H.; Yoon, S.J.; Jin, J.; Choi, S.H.; Seol, H.J.; Lee, J.-I.; Nam, D.H.; Yoo, H.Y. Inhibition of checkpoint kinase 1 sensitizes lung cancer brain metastases to radiotherapy. Biochem. Biophys. Res. Commun. 2011, 406, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Takahashi, A.; Ohnishi, K.; Ota, I.; Ohnishi, T.; Tojo, T.; Taniguchi, S. Effect of rapamycin, an mTOR inhibitor, on radiation sensitivity of lung cancer cells having different p53 gene status. Int. J. Oncol. 2010, 37, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yan, Y.; Xiao, L.; Dai, S.; Zeng, S.; Qian, L.; Wang, L.; Yang, X.; Xiao, Y.; Gong, Z. Radiosensitizing effect of diosmetin on radioresistant lung cancer cells via Akt signaling pathway. PLoS ONE 2017, 12, e0175977. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, T.; Okamoto, I.; Suzuki, M.; Nakahara, T.; Yamanaka, K.; Hatashita, E.; Yamada, Y.; Fukuoka, M.; Ono, K.; Nakagawa, K. Radiosensitizing effect of YM155, a novel small-molecule survivin suppressant, in non-small cell lung cancer cell lines. Clin. Cancer Res. 2008, 14, 6496–6504. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Giacalone, N.J.; Lu, B. Terameprocol (tetra-O-methyl nordihydroguaiaretic acid), an inhibitor of Sp1-mediated survivin transcription, induces radiosensitization in non-small cell lung carcinoma. J. Thorac. Oncol. 2011, 6, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Fujita, E.; Nagao, Y.; Node, M.; Kaneko, K.; Nakazawa, S.; Kuroda, H. Antitumor activity of the Isodon diterpenoids: Structural requirements for the activity. Experientia 1976, 32, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Takeda, Y.; Sun, H.D.; Minami, Y.; Marunaka, T.; Takeda, S.; Yamada, Y.; Togo, T. Cytotoxic and antitumor activities of Rabdosia diterpenoids. Planta Med. 1988, 54, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lu, X.-J.; Zhang, J.; Diao, H.; Li, G.; Xu, L.; Wang, T.; Wei, J.; Meng, W.; Ma, J.L.; et al. Oridonin, a novel lysine acetyltransferases inhibitor, inhibits proliferation and induces apoptosis in gastric cancer cells through p53- and caspase-3-mediated mechanisms. Oncotarget 2016, 7, 22623–22631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.W.; Ma, Y.Y.; Guan, T.P.; Xia, Y.J.; Shao, C.M.; Chen, L.G.; Ren, Y.J.; Yao, H.B.; Yang, Q.; He, X.J. Oridonin induces apoptosis in gastric cancer through Apaf-1, cytochrome c and caspase-3 signaling pathway. World J. Gastroenterol. 2012, 18, 7166–7174. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Xie, F.; Li, M.; Liang, Z.; Xu, W.; Yang, J.; Liu, C.; Li, H.; Zhou, H.; Qu, L.H. Oridonin induces autophagy via inhibition of glucose metabolism in p53-mutated colorectal cancer cells. Cell Death Dis. 2017, 8, e2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhong, Z.; Wan, J.; Tan, W.; Wu, G.; Chen, M.; Wang, Y. Oridonin induces apoptosis, inhibits migration and invasion on highly-metastatic human breast cancer cells. Am. J. Chin. Med. 2013, 41, 177–196. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Dong, B.; Zhao, P.; Zhou, H.; Qu, L. Oridonin Triggers Chaperon- mediated Proteasomal Degradation of BCR-ABL in Leukemia. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Wu, L.J.; Tashiro, S.I.; Onodera, S.; Ikejima, T. Oridonin induces apoptosis of HeLa cells via altering expression of Bcl-2/Bax and activating caspase-3/ICAD pathway. Acta Pharmacol. Sin. 2004, 25, 691–698. [Google Scholar] [PubMed]
- Ye, Y.C.; Wang, H.J.; Xu, L.; Liu, W.W.; Liu, B.B.; Tashiro, S.I.; Onodera, S.; Ikejima, T. Oridonin induces apoptosis and autophagy in murine fibrosarcoma L929 cells partly via NO-ERK-p53 positive-feedback loop signaling pathway. Acta Pharmacol. Sin. 2012, 33, 1055–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikezoe, T.; Yang, Y.; Bandobashi, K.; Saito, T.; Takemoto, S.; Machida, H.; Togitani, K.; Koeffler, H.P.; Taguchi, H. Oridonin, a diterpenoid purified from Rabdosia rubescens, inhibits the proliferation of cells from lymphoid malignancies in association with blockade of the NF-κB signal pathways. Mol. Cancer Ther. 2005, 4, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wei, W.; Zhang, N.; Yu, Q.; Xu, W.B.; Yu, W.J.; Chen, G.Q.; Wu, Y.L.; Yan, H. Oridonin stabilizes retinoic acid receptor alpha through ROS-activated NF-κB signaling. BMC Cancer 2015, 15, 248. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Cai, H.; Jin, H.; Yang, F.; Jiang, J.; Wu, A.; Zhu, H.; Liu, J.; Su, X.; Yang, P.; et al. Qualitative and Quantitative Analysis of ROS-Mediated Oridonin-Induced Oesophageal Cancer KYSE-150 Cell Apoptosis by Atomic Force Microscopy. PLoS ONE 2015, 10, e0140935. [Google Scholar] [CrossRef] [PubMed]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulms, D.; Zeise, E.; Pöppelmann, B.; Schwarz, T. DNA damage, death receptor activation and reactive oxygen species contribute to ultraviolet radiation-induced apoptosis in an essential and independent way. Oncogene 2002, 21, 5844–5851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyori, B.M.; Venkatachalam, G.; Thiagarajan, P.S.; Hsu, D.; Clement, M.V. OpenComet: An automated tool for comet assay image analysis. Redox Biol. 2014, 2, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, L.; Schulte, B.A.; Yang, A.; Tang, S.; Wang, G.Y. Resveratrol enhances ionizing radiation-induced premature senescence in lung cancer cells. Int. J. Oncol. 2013, 43, 1999–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, E.J.; Wuu, C.S. Radiation-induced second cancers: The impact of 3D-CRT and IMRT. Int. J. Radiat. Oncol. Biol. 2003, 56, 83–88. [Google Scholar] [CrossRef]
- Kim, M.Y.; Park, S.J.; Shim, J.W.; Yang, K.; Kang, H.S.; Heo, K. Naphthazarin enhances ionizing radiation-induced cell cycle arrest and apoptosis in human breast cancer cells. Int. J. Oncol. 2015, 46, 1659–1666. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Negi, A.S.; Kumar, J.K.; Gupta, M.M.; Khanuja, S.P.S. Plant-based anticancer molecules: A chemical and biological profile of some important leads. Bioorg. Med. Chem. 2005, 13, 5892–5908. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Sultana, M.; Qazi, A.; Qazi, M.H.; Parveen, G.; Waquar, G.S.; Ashraf, A.B.; Rasool, M. Role of Natural Radiosensitizers and Cancer Cell Radioresistance: An Update. Anal. Cell Pathol. 2016, 2016, 6146595. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, X.; Li, X.; Wang, J.; Ye, Z.; Li, J.C. Oridonin Up-regulates Expression of P21 and Induces Autophagy and Apoptosis in Human Prostate Cancer Cells. Int. J. Biol. Sci. 2012, 8, 901–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Lv, Y.F.; Lu, L.; Cai, L. Oridonin inhibits mTOR signaling and the growth of lung cancer tumors. Anticancer Drug 2014, 25, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.H.; Chai, K.; Tashiro, S.I.; Onodera, S.; Ikejima, T. Inhibition of c-Met promoted apoptosis, autophagy and loss of the mitochondrial transmembrane potential in oridonin-induced A549 lung cancer cells. J. Pharm. Pharmacol. 2013, 65, 1622–1642. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Shan, Q.; Gong, Y.; Lin, J.; Yang, X.; Zhou, R. Oridonin in combination with imatinib exerts synergetic anti-leukemia effect in Ph+ acute lymphoblastic leukemia cells in vitro by inhibiting activation of LYN/mTOR signaling pathway. Cancer Biol. Ther. 2012, 13, 1244–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, L.; Zi, Y.; Zhang, L.; Guo, Y.; Huang, Y. Oridonin effectively reverses the drug resistance of cisplatin involving induction of cell apoptosis and inhibition of MMP expression in human acute myeloid leukemia cells. Saudi J. Biol. Sci. 2017, 24, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ding, Y.; Chen, C.H.; Zhou, Z.; Ding, C.; Chen, H.; Zhou, J.; Chen, C. A new oridonin analog suppresses triple-negative breast cancer cells and tumor growth via the induction of death receptor 5. Cancer Lett. 2016, 380, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Citrin, D.E.; Mitchell, J.B. Altering the response to radiation: Sensitizers and protectors. Semin. Oncol. 2014, 41, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Sun, Y.; Zhu, J.; Yu, L.; Jiang, X.; Zhang, J.; Dong, X.; Ma, B.; Zhang, Q. Oridonin exerts anticancer effect on osteosarcoma by activating PPAR-γ and inhibiting Nrf2 pathway. Cell Death Dis. 2018, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Qian, C.; Shen, Z. Anti-tumor activity of oridonin on SNU-5 subcutaneous xenograft model via regulation of c-Met pathway. Tumor Biol. 2014, 35, 9139–9146. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.Q.; Luo, J.; Chen, H.; Zhang, J.H.; Li, H.H.; Guo, H.C.; Wang, Z.H.; Lin, S.Z. Oridonin enhances antitumor activity of gemcitabine in pancreatic cancer through MAPK-p38 signaling pathway. Int. J. Oncol. 2012, 41, 949–958. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.; Jeong, Y.J.; Han, N.-K.; Kim, J.S.; Lee, H.-J. Oridonin Enhances Radiation-Induced Cell Death by Promoting DNA Damage in Non-Small Cell Lung Cancer Cells. Int. J. Mol. Sci. 2018, 19, 2378. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082378
Park H, Jeong YJ, Han N-K, Kim JS, Lee H-J. Oridonin Enhances Radiation-Induced Cell Death by Promoting DNA Damage in Non-Small Cell Lung Cancer Cells. International Journal of Molecular Sciences. 2018; 19(8):2378. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082378
Chicago/Turabian StylePark, Hyejin, Ye Ji Jeong, Na-Kyung Han, Joong Sun Kim, and Hae-June Lee. 2018. "Oridonin Enhances Radiation-Induced Cell Death by Promoting DNA Damage in Non-Small Cell Lung Cancer Cells" International Journal of Molecular Sciences 19, no. 8: 2378. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082378