Jatrorrhizine Hydrochloride Suppresses RANKL-Induced Osteoclastogenesis and Protects against Wear Particle-Induced Osteolysis

Abstract
:1. Introduction
2. Results
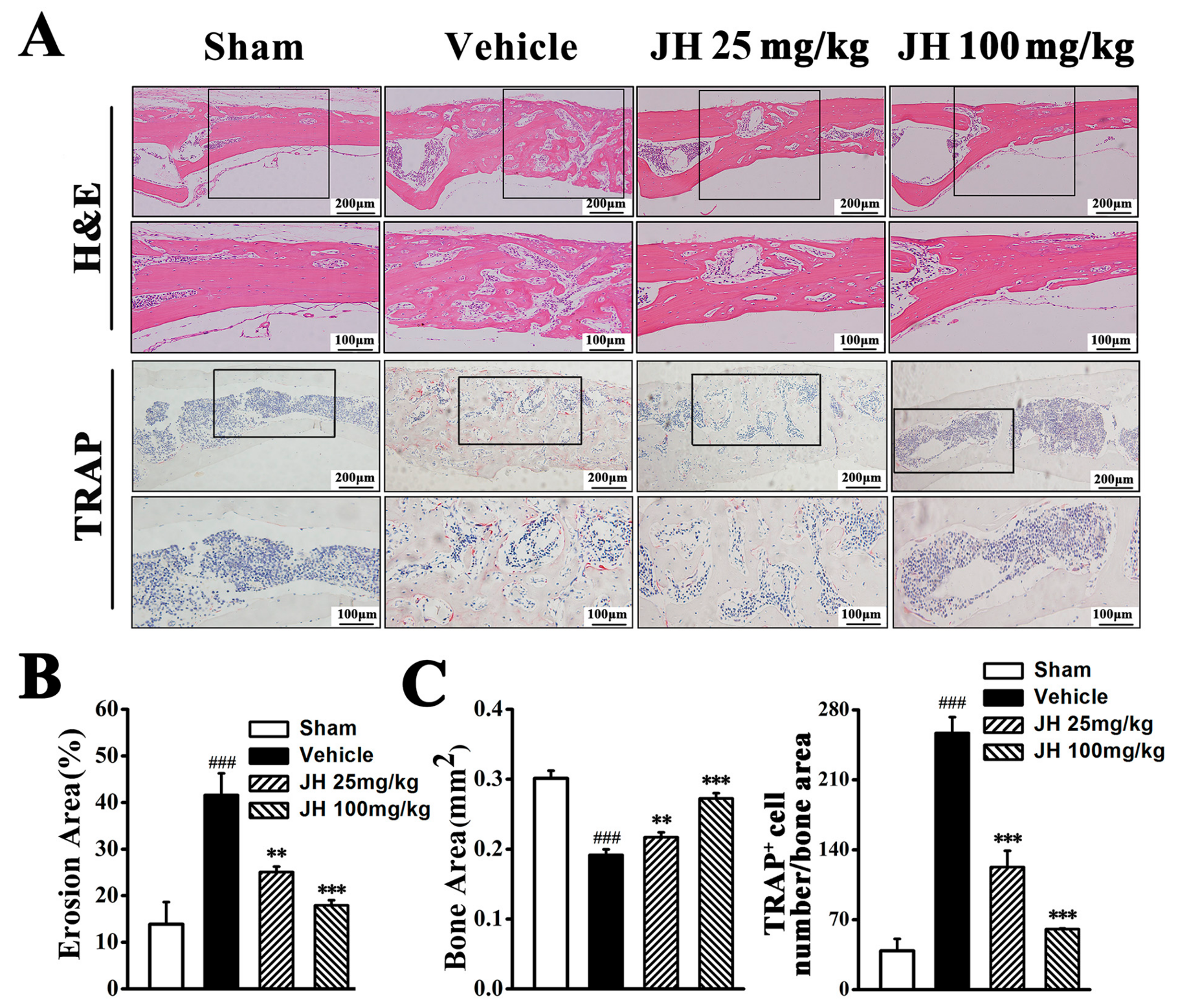
2.1. Therapeutic Effect of JH on Ti Particle-Induced Osteolysis In Vivo
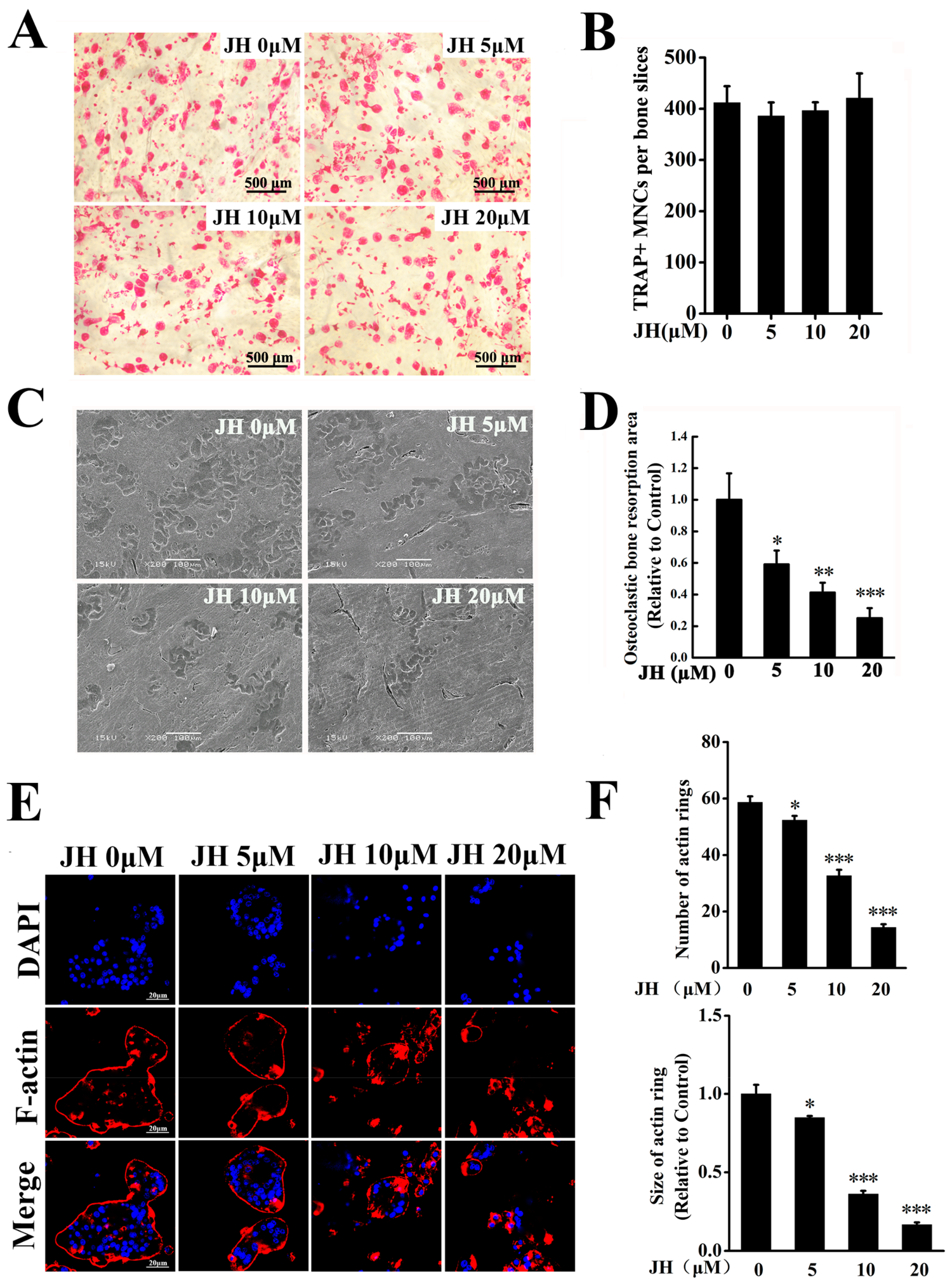
2.2. JH Inhibited RANKL-Induced Osteoclast Formation In Vitro
2.3. JH Impaired Ostoclastic Bone Resorption and F-Actin Ring Formation In Vitro
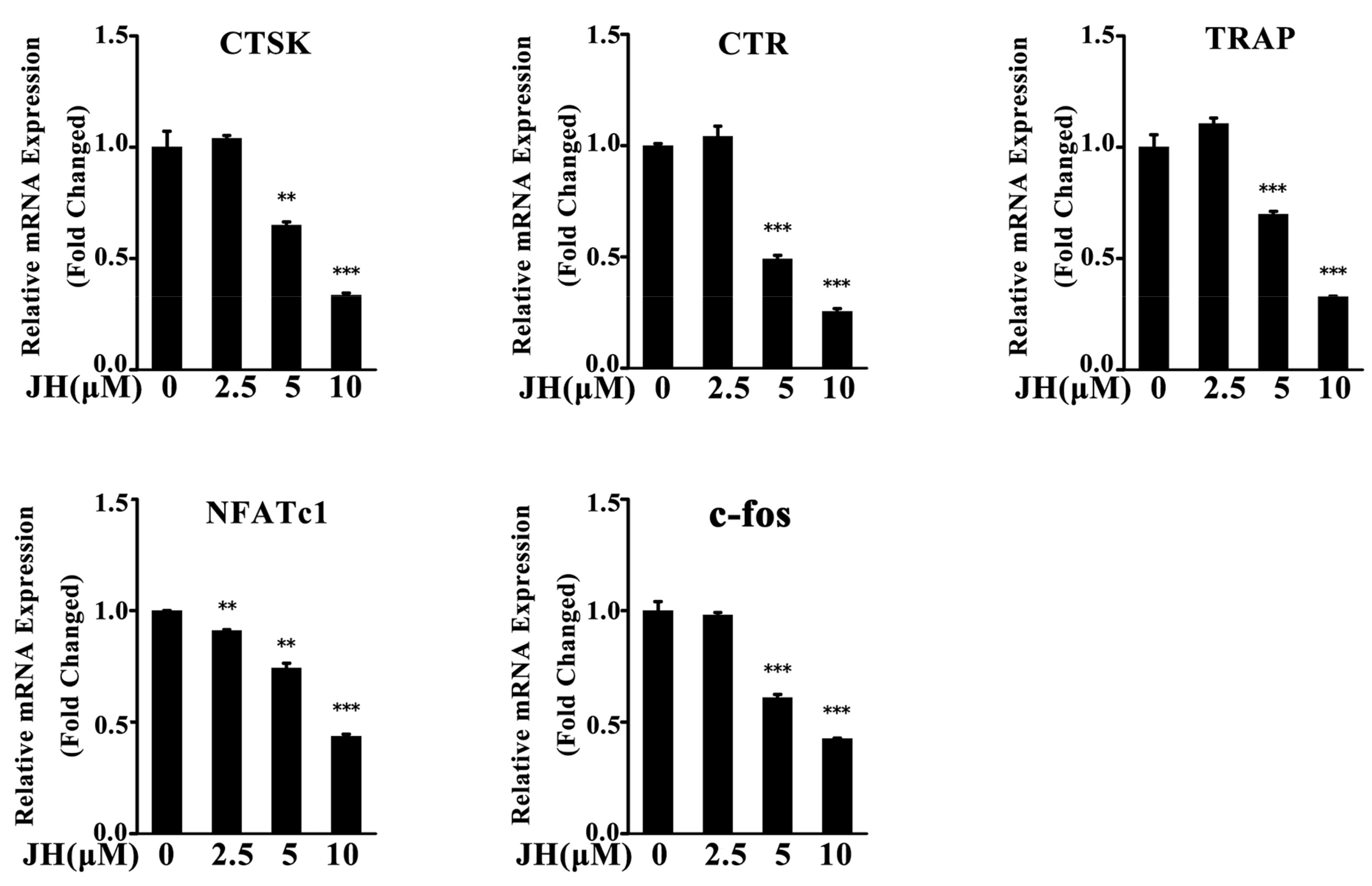
2.4. JH Inhibited RANKL-Induced Osteoclast-Specific Gene Expression
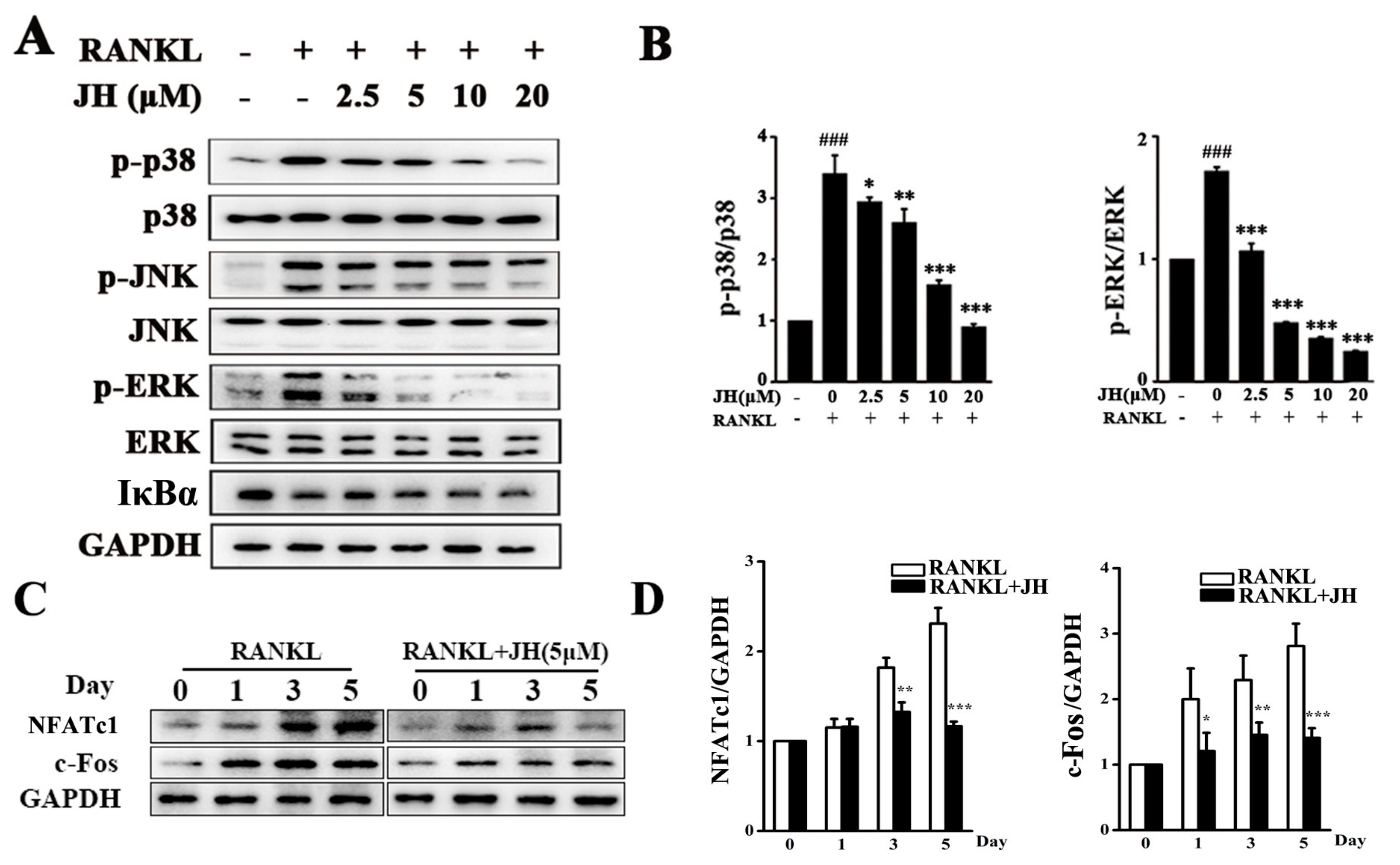
2.5. JH Inhibited RANKL-Induced MAPK and NFATc1 Activation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Media and Reagents
4.3. Titanium Particle-Induced Calvarial Osteolysis Model
4.4. Serum ALT and AST Measurement
4.5. Micro-CT Scanning
4.6. Histological and Histomorphometric Analysis
4.7. Cell Viability Assay
4.8. In Vitro Osteoclastogenesis Assay
4.9. Bone Resorption Pit Assay and F-Actin Ring Formation Assay
4.10. Apoptosis Analysis by Flow Cytometry
4.11. RNA Isolation and Real-Time PCR Analysis
4.12. Western Blot Analysis
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Harris, W.H. Wear and periprosthetic osteolysis: The problem. Clin. Orthop. 2001, 393, 66–70. [Google Scholar] [CrossRef]
- Yunpeng, J.; Tanghong, J.; Wooley, P.H.; Shang-You, Y. Current research in the pathogenesis of aseptic implant loosening associated with particulate wear debris. Acta Orthop. Belg. 2013, 79, 1–9. [Google Scholar]
- Gallo, J.; Vaculova, J.; Goodman, S.B.; Konttinen, Y.T.; Thyssen, J.P. Contributions of human tissue analysis to understanding the mechanisms of loosening and osteolysis in total hip replacement. Acta Biomater. 2014, 10, 2354–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundfeldt, M.; Carlsson, L.V.; Johansson, C.B.; Thomsen, P.; Gretzer, C. Aseptic loosening, not only a question of wear: A review of different theories. Acta Orthop. 2006, 77, 177–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, G.; Murnaghan, C.; Reilly, J.; Meek, R.M. The biology of aseptic osteolysis. Clin. Orthop. Relat. Res. 2007, 460, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Purdue, P.E.; Koulouvaris, P.; Potter, H.G.; Nestor, B.J.; Sculco, T.P. The cellular and molecular biology of periprosthetic osteolysis. Clin. Orthop. Relat. Res. 2007, 454, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Waddell, J.; Pritzker, K.P.; Boynton, E.L. Increased cytokine secretion in patients with failed implants compared with patients with primary implants. Clin. Orthop. Relat. Res. 2005, 434, 170–176. [Google Scholar] [CrossRef]
- Weinstein, R.S.; Roberson, P.K.; Manolagas, S.C. Giant osteoclast formation and long-term oral bisphosphonate therapy. N. Engl. J. Med. 2009, 360, 1677–1678. [Google Scholar] [CrossRef] [PubMed]
- Verron, E.; Bouler, J.M. Is bisphosphonate therapy compromised by the emergence of adverse bone disorders? Drug Discov. Today 2014, 19, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Gannon, S.C.; Cantley, M.D.; Haynes, D.R.; Hirsch, R.; Bartold, P.M. Azithromycin suppresses human osteoclast formation and activity in vitro. J. Cell Physiol. 2013, 228, 1098–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olof, S.; Agata, R.; Thomas, E.; Mats, S.; Henrik, B.; Olle, M. Denosumab for treating periprosthetic osteolysis; study protocol for a randomized, double-blind, placebo-controlled trial. BMC Musculoskelet. Disord. 2016, 17, 1–10. [Google Scholar]
- Yamazaki, T.; Yamori, M.; Yamamoto, K.; Saito, K.; Asai, K.; Sumi, E.; Goto, K.; Takahashi, K.; Nakayama, T.; Bessho, K. Risk of osteomyelitis of the jaw induced by oral bisphosphonates in patients taking medications for osteoporosis: A hospital-based cohort study in Japan. Bone 2012, 51, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Mcclung, M.; Harris, S.T.; Miller, P.D.; Bauer, D.C.; Davison, K.S.; Dian, L.; Hanley, D.A.; Kendler, D.L.; Yuen, C.K.; Lewiecki, E.M. Bisphosphonate therapy for osteoporosis: Benefits, risks, and drug holiday. Am. J. Med. 2013, 126, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Marx, R.E. The deception and fallacies of sponsored randomized prospective double-blinded clinical trials: The bisphosphonate research example. Int. J. Oral. Maxillofac. Implant. 2014, 29, e37–e44. [Google Scholar] [CrossRef] [PubMed]
- Hiligsmann, M.; Boonen, A.; Dirksen, C.D.; Sedrine, W.B.; Reginster, J.Y. Cost-effectiveness of denosumab in the treatment of postmenopausal osteoporotic women. Expert. Rev. Pharmacoecon. Outcomes Res. 2013, 13, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Slobodníková, L.; Kost’Álová, D.; Labudová, D.; Kotulová, D.; Kettmann, V. Antimicrobial activity of Mahonia aquifolium crude extract and its major isolated alkaloids. Phytother. Res. 2004, 18, 674–676. [Google Scholar] [CrossRef] [PubMed]
- Volleková, A.; Košt’Álová, D.; Kettmann, V.; Tóth, J. Antifungal activity of Mahonia aquifolium extract and its major protoberberine alkaloids. Phytother Res. 2003, 17, 834–837. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Liao, J.W.; Peng, W.H.; Lee, M.S.; Pao, L.H.; Cheng, H.Y. Antioxidant, Analgesic, Anti-Inflammatory, and Hepatoprotective Effects of the Ethanol Extract of Mahonia oiwakensis Stem. Int. J. Mol. Sci. 2013, 14, 2928–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Cao, Z.; Pan, Y.; Zhang, G.; Yang, P.; Guo, P.; Zhou, Q. Jatrorrhizine hydrochloride inhibits the proliferation and neovascularization of C8161 metastatic melanoma cells. Anticancer Drugs 2013, 24, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Ng, L.T.; Hsu, F.F.; Shieh, D.E.; Chiang, L.C. Cytotoxic effects of Coptis chinensis and Epimedium sagittatum extracts and their major constituents (berberine, coptisine and icariin) on hepatoma and leukaemia cell growth. Clin. Exp. Pharmacol. Physiol. 2004, 31, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.U.; Ben-Rong, H.U.; Tang, Q.; Qin, F.U.; Zhang, Q.Y.; Xiang, J.Z. Effect of jatrorrhizine, berberine, Huanglian Decoction and compound-mimic prescription on blood glucose in mice. Chin. Tradit. Herb. Drugs 2005, 36, 548–551. [Google Scholar]
- Yang, W.; She, L.; Yu, K.; Yan, S.; Zhang, X.; Tian, X.; Ma, S.; Zhang, X. Jatrorrhizine hydrochloride attenuates hyperlipidemia in a high-fat diet-induced obesity mouse model. Mol. Med. Rep. 2016, 14, 3277–3284. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Sun, S.; Ma, X.; Cui, C.; Chen, G.; Liu, Z.; Li, H.; Liu, M. Jatrorrhizine Hydrochloride Suppresses Proliferation, Migration, and Secretion of Synoviocytes In Vitro and Ameliorates Rat Models of Rheumatoid Arthritis In Vivo. Int. J. Mol. Sci. 2018, 19, 1514. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef]
- Matsuo, K.; Galson, D.L.; Zhao, C.; Peng, L.; Laplace, C.; Wang, K.Z.; Bachler, M.A.; Amano, H.; Aburatani, H.; Ishikawa, H.; et al. Nuclear factor of activated T-cells (NFAT) rescues osteoclastogenesis in precursors lacking c-Fos. J. Biol. Chem. 2004, 279, 26475–26480. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.P.; Na, K.L.; Lee, S.Y. Current Understanding of RANK Signaling in Osteoclast Differentiation and Maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar]
- David, J.P.; Sabapathy, K.; Behrens, A.; Berger, B.; Hochedlinger, K.; Hoffmann, O.; Wagner, E.F. c-Jun phosphorylation by JNK1 but not by JNK2 is necessary for efficient osteoclastogenesis in response to TRANCE stimulation. J. Bone Miner. Res. 1999, 14, S149. [Google Scholar]
- Ang, E.S.M.; Pavlos, N.J.; Chim, S.M.; Feng, H.T.; Scaife, R.M.; Steer, J.H.; Zheng, M.H.; Xu, J. Paclitaxel inhibits osteoclast formation and bone resorption via influencing mitotic cell cycle arrest and RANKL-induced activation of NF-κB and ERK. J. Cell. Biochem. 2012, 113, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Lim, H.; Jeong, J.E.; Yim, M. Attenuation of RANKL-induced Osteoclast Formation via p38-mediated NFATc1 Signaling Pathways by Extract ofEuphorbia LathyrisL. J. Bone Metab. 2016, 23, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Soysa, N.S.; Alles, N.; Aoki, K.; Ohya, K. Osteoclast formation and differentiation: An overview. J. Med. Dent. Sci. 2016, 59, 65–74. [Google Scholar]
- Xu, J.; Wu, H.; Ang, E.S.; Yip, K.; Woloszyn, M.; Zheng, M.; Tan, R. NF-κB modulators in osteolytic bone diseases. Cytokine Growth Factor Rev. 2009, 20, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, F.; Nishimura, R.; Matsubara, T.; Tanaka, S.; Inoue, J.-I.; Reddy, S.V.; Hata, K.; Yamashita, K.; Hiraga, T.; Watanabe, T.; et al. Critical roles of c-Jun signaling in regulation of NFAT family and RANKL-regulated osteoclast differentiation. J. Clin. Investig. 2004, 114, 475–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayanagi, H. New developments in osteoimmunology. Nat. Rev. Rheumatol. 2012, 8, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Ishisaki, A.; Kawashita, E.; Chosa, N.; Nakajima, K.; Nishihara, T.; Toyoshima, K.; Okada, K.; Ueshima, S.; Matsushita, K.; et al. Plasminogen/Plasmin Modulates Bone Metabolism by Regulating the Osteoblast and Osteoclast Function. J. Biol. Chem. 2014, 289, 8952–8960. [Google Scholar] [CrossRef]
- Wedemeyer, C.; Von, K.F.; Pingsmann, A.; Hilken, G.; Sprecher, C.; Saxler, G.; Henschke, F.; Löer, F.; Von, K.M. Stimulation of bone formation by zoledronic acid in particle-induced osteolysis. Biomaterials 2005, 26, 3719–3725. [Google Scholar] [CrossRef] [PubMed]
- Córdova, L.A.; Trichet, V.; Escriou, V.; Rosset, P.; Amiaud, J.; Battaglia, S.; Charrier, C.; Berreur, M.; Brion, R.; Gouin, F.; et al. Inhibition of osteolysis and increase of bone formation after local administration of siRNA-targeting RANK in a polyethylene particle-induced osteolysis model. Acta Biomater. 2015, 13, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Kaar, S.G.; Ragab, A.A.; Kaye, S.J.; Kilic, B.A.; Jinno, T.; Goldberg, V.M.; Bi, Y.; Stewart, M.C.; Carter, J.R.; Greenfield, E.M. Rapid repair of titanium particle-induced osteolysis is dramatically reduced in aged mice. J. Orthop. Res. 2001, 19, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, K.; Bauer, T.W.; Stulberg, B.N.; Wilde, A.H. Comparison and quantitation of wear debris of failed total hip and total knee arthroplasty. J. Biomed. Mater. Res. 1996, 31, 257–263. [Google Scholar] [CrossRef]
- Von, K.M.; Jewison, D.E.; Sibonga, J.D.; Sprecher, C.; Morrey, B.F.; Loer, F.; Berry, D.J.; Scully, S.P. The effectiveness of polyethylene versus titanium particles in inducing osteolysis in vivo. J. Orthop. Res. 2004, 22, 237–243. [Google Scholar] [Green Version]
- Taki, N.; Tatro, J.M.; Nalepka, J.L.; Togawa, D.; Goldberg, V.M.; Rimnac, C.M.; Greenfield, E.M. Polyethylene and titanium particles induce osteolysis by similar, lymphocyte-independent, mechanisms. J. Orthop. Res. 2005, 23, 376–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, F.; Zhai, Z.; Jiang, C.; Liu, X.; Li, H.; Qu, X.; Ouyang, Z.; Fan, Q.; Tang, T.; Qin, A.; et al. Geraniin suppresses RANKL-induced osteoclastogenesis in vitro and ameliorates wear particle-induced osteolysis in mouse model. Exp. Cell Res. 2015, 330, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wang, W.; Tian, B.; Liu, X.; Qu, X.; Zhai, Z.; Li, H.; Liu, F.; Fan, Q.; Tang, T.; et al. Myricetin prevents titanium particle-induced osteolysis in vivo and inhibits RANKL-induced osteoclastogenesis in vitro. Biochem. Pharmacol. 2015, 93, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xu, L.; Ma, X.; Xu, J.; Wang, J.; Xian, M.; Zhou, X.; Wang, M.; Wang, F.; Qin, A.; et al. MAGED1 Is a Negative Regulator of Bone Remodeling in Mice. Am. J. Pathol. 2015, 185, 2653–2667. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | Vehicle | JH (25 mg∙kg−1) | JH (100 mg∙kg−1) |
|---|---|---|---|---|
| Initial weight (g) | 22.02 ± 1.30 | 21.60 ± 0.61 | 21.99 ± 0.57 | 21.46 ± 0.41 |
| Final weight (g) | 22.29 ± 1.56 | 22.37 ± 0.42 | 22.45 ± 0.76 | 22.94 ± 0.73 |
| AST (U/L) | 27.88 ± 1.80 | 29.37 ± 2.48 | 22.96 ± 4.47 | 22.55 ± 3.31 |
| ALT (U/L) | 9.99 ± 1.83 | 8.68 ± 1.84 | 8.60 ± 0.91 | 9.06 ± 2.15 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Wang, J.; Sun, Q.; Chen, G.; Sun, S.; Ma, X.; Qiu, H.; Liu, X.; Xu, L.; Liu, M. Jatrorrhizine Hydrochloride Suppresses RANKL-Induced Osteoclastogenesis and Protects against Wear Particle-Induced Osteolysis. Int. J. Mol. Sci. 2018, 19, 3698. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113698
Li H, Wang J, Sun Q, Chen G, Sun S, Ma X, Qiu H, Liu X, Xu L, Liu M. Jatrorrhizine Hydrochloride Suppresses RANKL-Induced Osteoclastogenesis and Protects against Wear Particle-Induced Osteolysis. International Journal of Molecular Sciences. 2018; 19(11):3698. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113698
Chicago/Turabian StyleLi, Hui, Jing Wang, Qiwen Sun, Gang Chen, Shengnan Sun, Xuemei Ma, Haiwen Qiu, Xuerong Liu, Liangyi Xu, and Mei Liu. 2018. "Jatrorrhizine Hydrochloride Suppresses RANKL-Induced Osteoclastogenesis and Protects against Wear Particle-Induced Osteolysis" International Journal of Molecular Sciences 19, no. 11: 3698. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113698