Genome-Wide Identification and Analysis of HAK/KUP/KT Potassium Transporters Gene Family in Wheat (Triticum aestivum L.)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification of the TaHAKs in Wheat
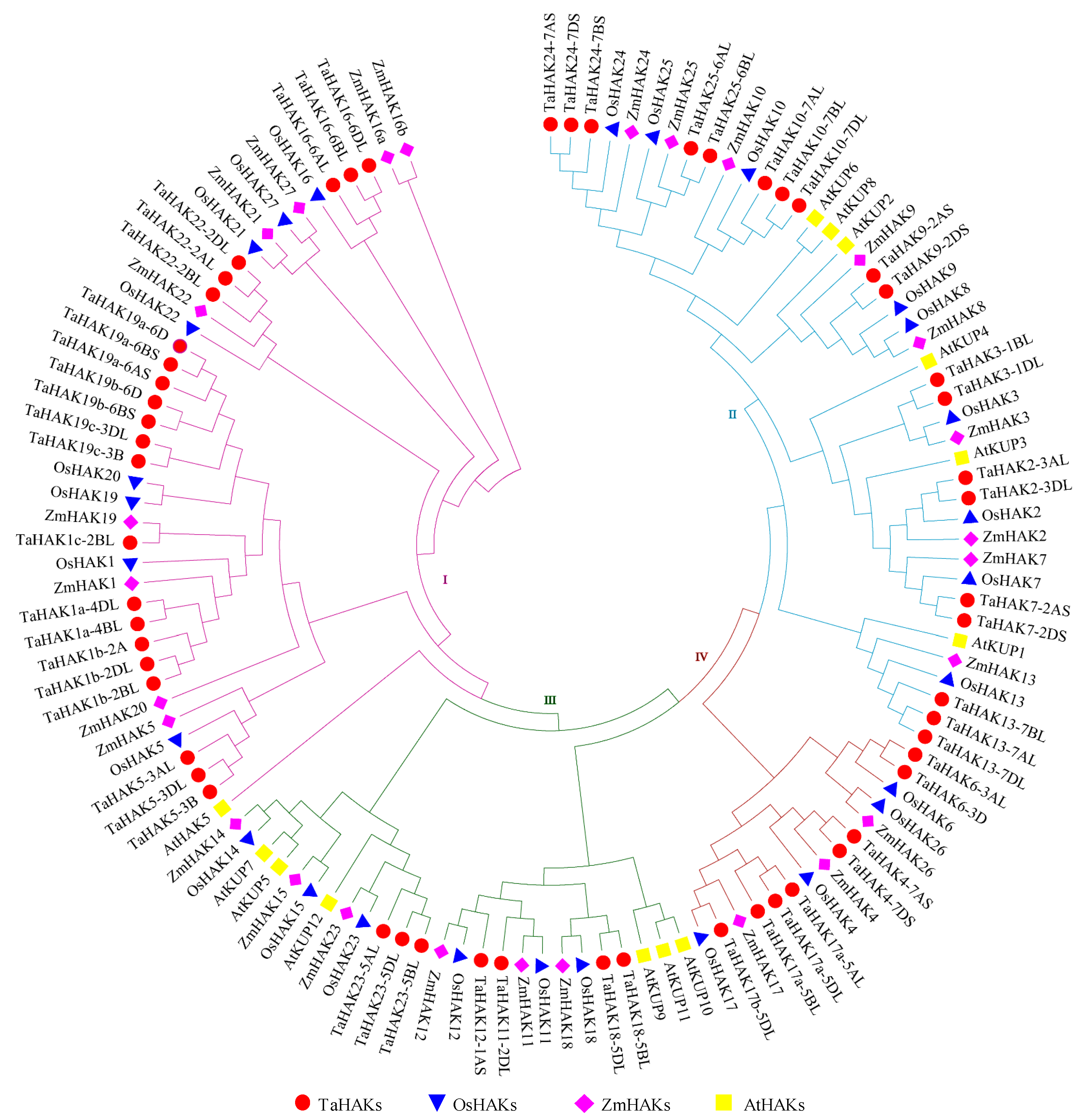
2.2. Phylogenetic Analysis of TaHAK Proteins
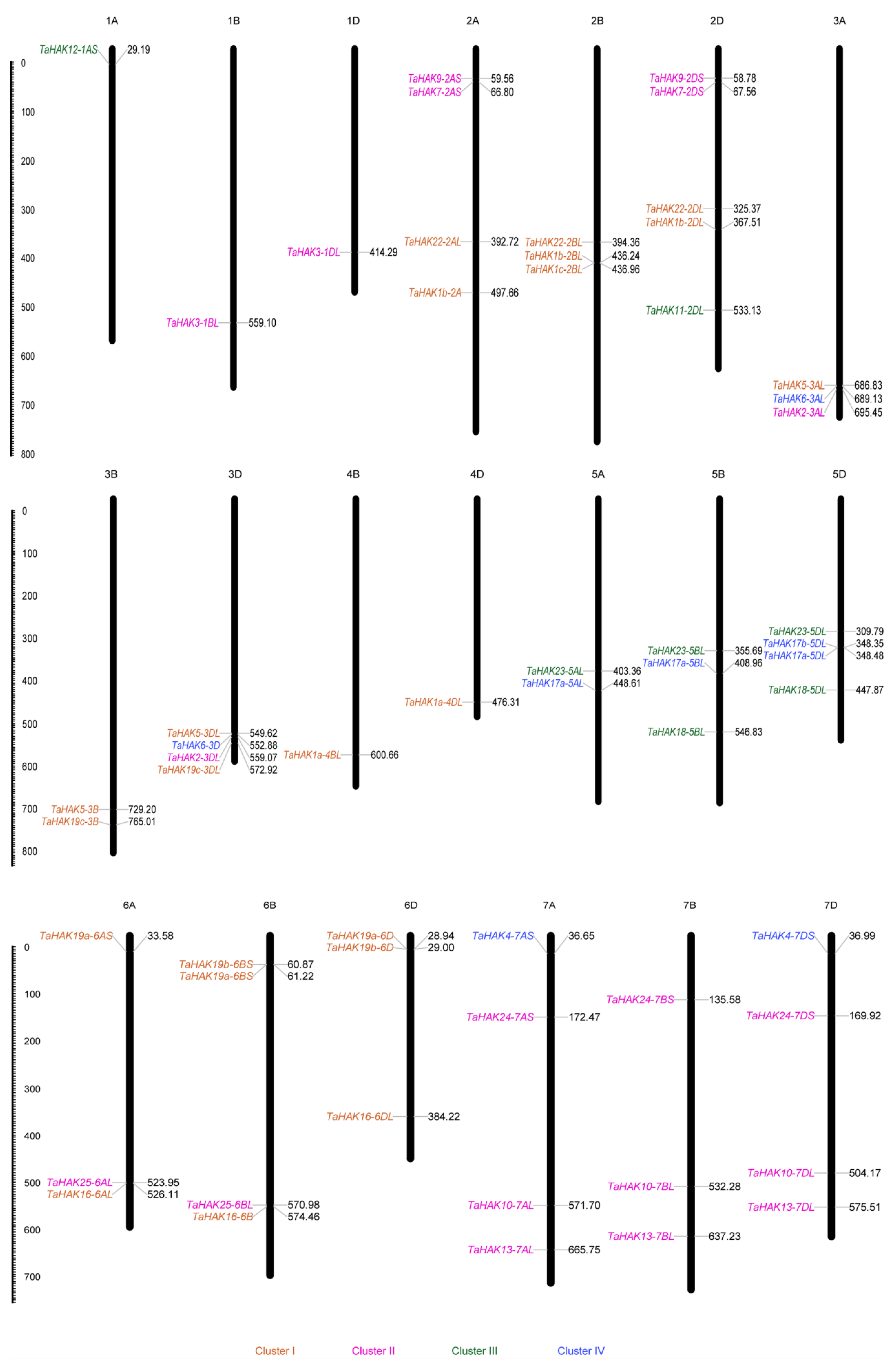
2.3. Genomic Distribution of TaHAKs
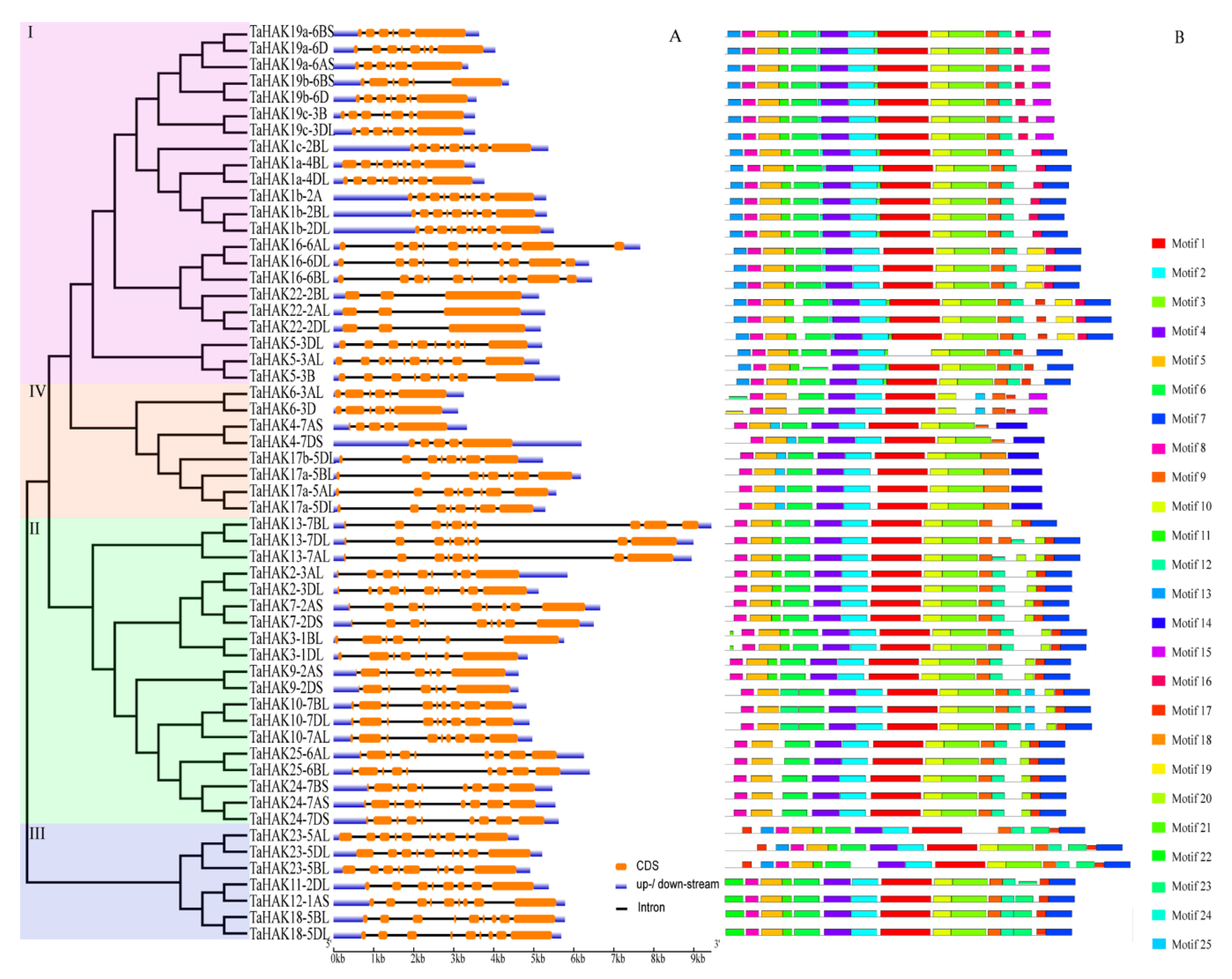
2.4. Gene Structure and Conserved Motif Analyses of TaHAKs
2.5. Expression Analysis of TaHAK Genes in Various Wheat Tissues
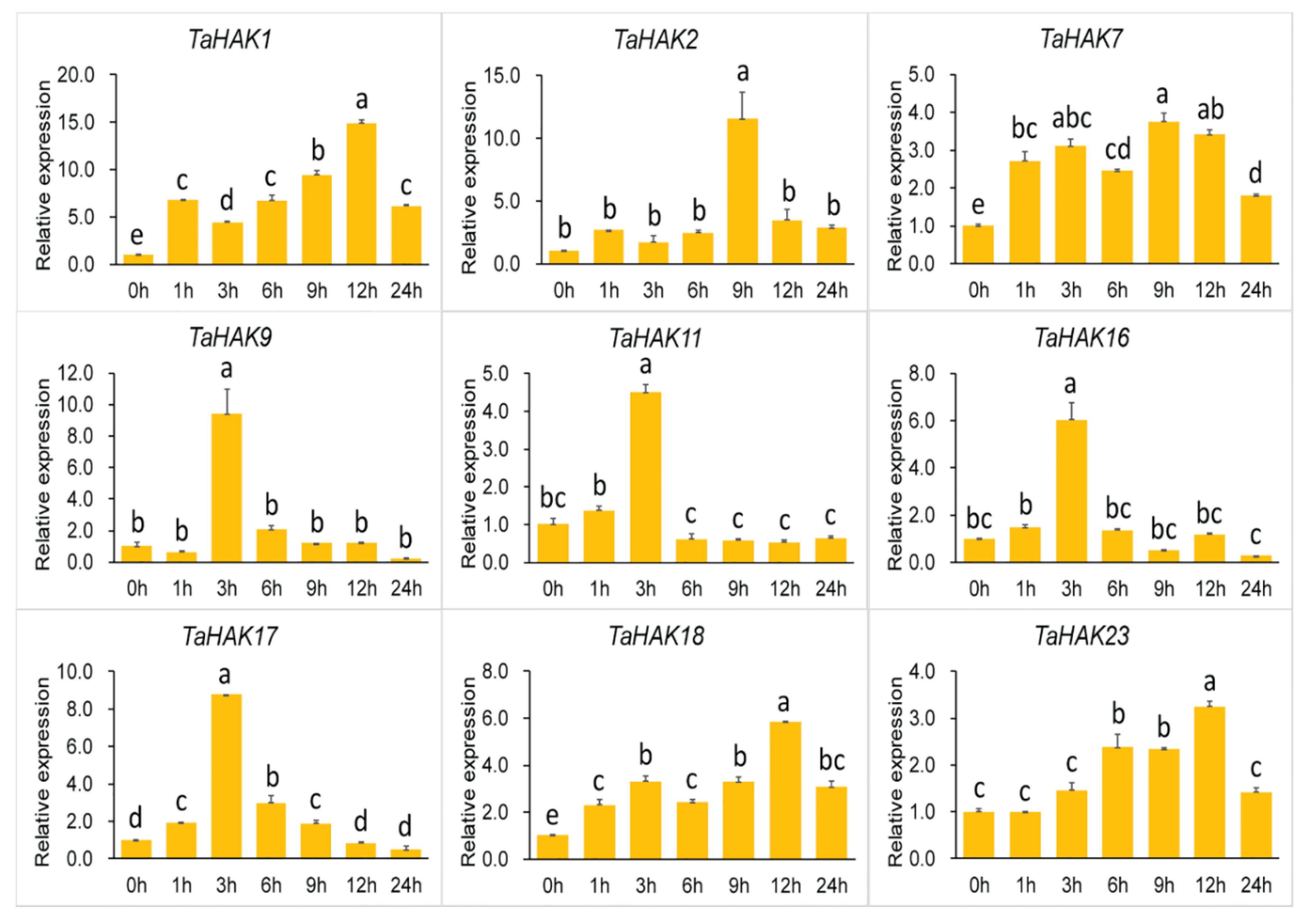
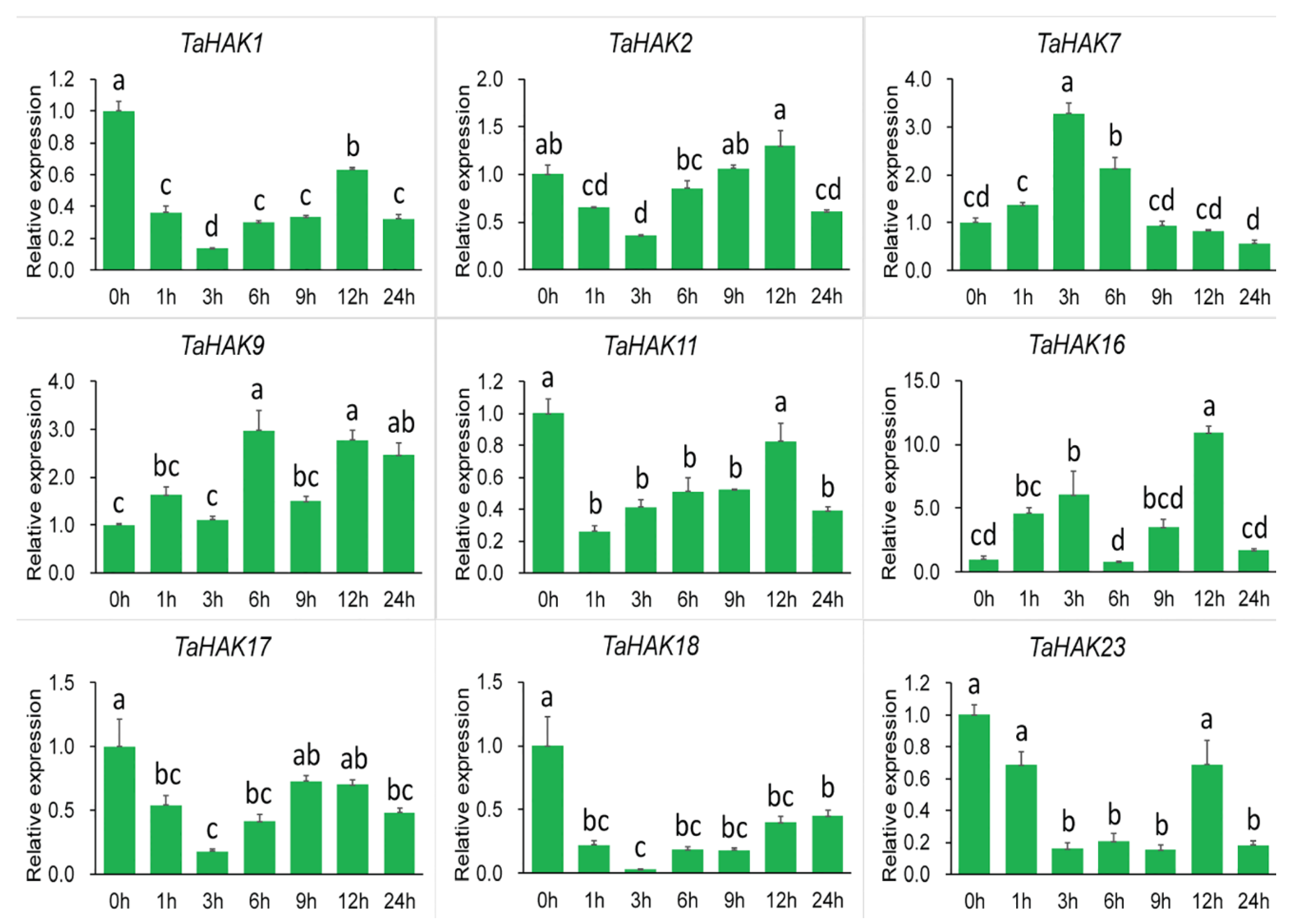
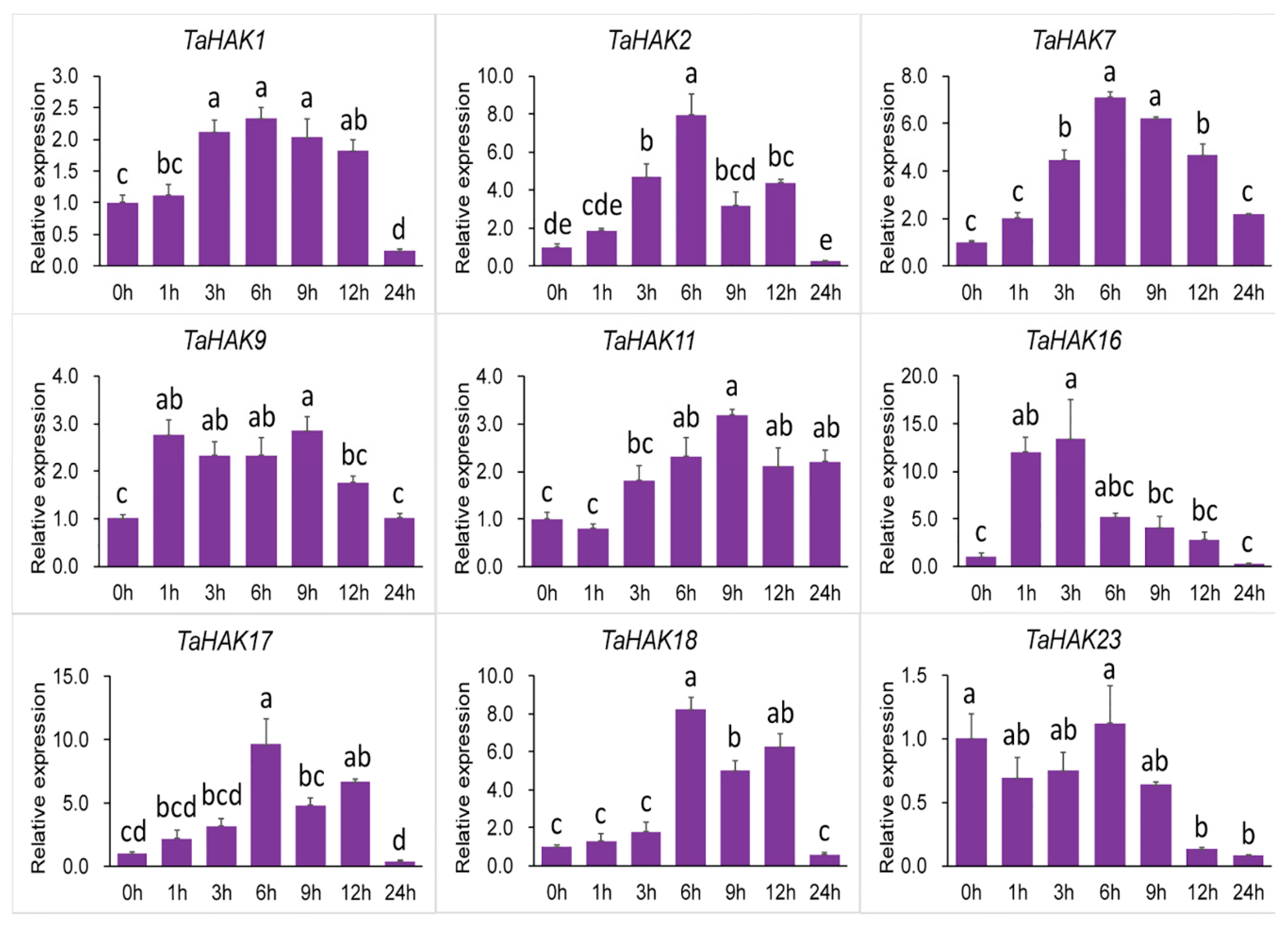
2.6. Expression Analysis of TaHAK Genes under Various Stresses
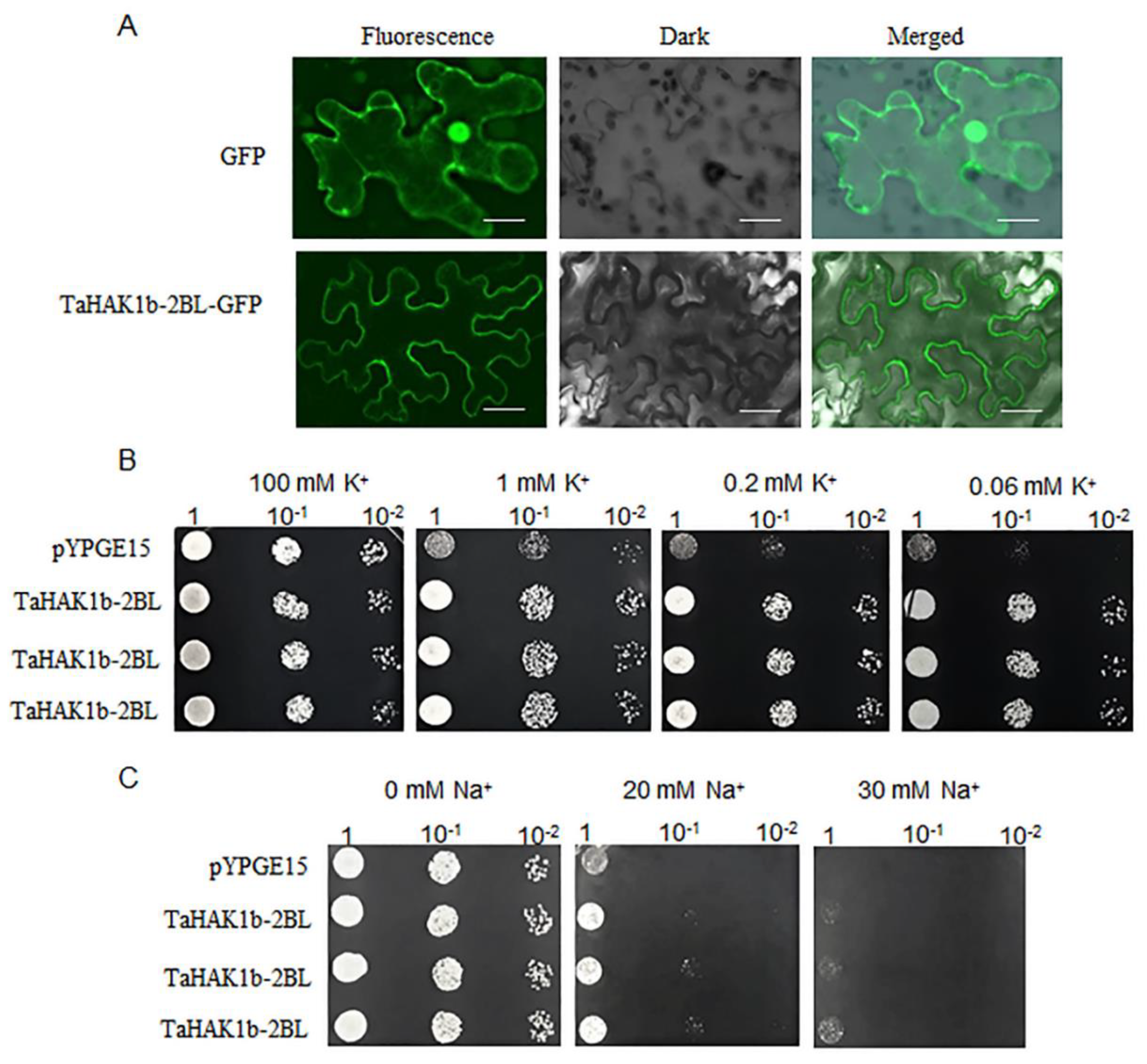
2.7. TaHAK1b-2BL Functions in Potassium Uptake and Salt Tolerance in Yeast
3. Discussion
4. Materials and Methods
4.1. Identification of TaHAK Genes in Wheat
4.2. Phylogenetic Analysis of TaHAKs
4.3. Physical Localization of TaHAK Genes
4.4. Analysis of Gene Structures and Protein Motifs
4.5. Plant Materials, Growth Conditions, and Experimental Treatments
4.6. RNA Isolation and qRT-PCR
4.7. Subcellular Location of the TaHAK1b-2BL Protein
4.8. Functional Complementation Assay of TaHAK1b-2BL in Yeast
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leigh, R.; Jones, R.W. A hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant-cell. New Phytol. 1984, 97, 1–13. [Google Scholar] [CrossRef]
- Wang, X.; Mohamed, I.; Ali, M.; Abbas, M.H.H.; Shah, G.M.; Chen, F. Potassium distribution in root and non-root zones of two cotton genotypes and its accumulation in their organs as affected by drought and potassium stress conditions. J. Plant Nutr. Soil Sci. 2018. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Nahar, K.; Hossain, M.; Mahmud, J.; Hossen, M.; Masud, A.; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, W.H. Regulation of potassium transport and signaling in plants. Curr. Opin. Plant Biol. 2017, 39, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed]
- Very, A.A.; Nieves-Cordones, M.; Daly, M.; Khan, I.; Fizames, C.; Sentenac, H. Molecular biology of K+ transport across the plant cell membrane: What do we learn from comparison between plant species? J. Plant Physiol. 2014, 171, 748–769. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef]
- Epstein, E.; Rains, D.; Elzam, O. Resolution of dual mechanisms of potassium absorption by barley roots. Proc. Natl. Acad. Sci. USA 1963, 49, 684–692. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Li, M.; Oh, S.; Kronzucker, H.J. Capacity and plasticity of potassium channels and high-affinity transporters in roots of barley and Arabidopsis. Plant Physiol. 2013, 162, 496–511. [Google Scholar] [CrossRef]
- Jordi, S.; Josep, P. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef]
- Gupta, M.; Qiu, X.; Wang, L.; Xie, W.; Zhang, C.; Xiong, L.; Lian, X.; Zhang, Q. KT/HAK/KUP potassium transporters gene family and their whole-life cycle expression profile in rice (Oryza sativa). Mol. Genet. Genom. 2008, 280, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Maser, P.; Thomine, S.; Julian, S.; Ward, J.M.; Hirschi, K. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef] [PubMed]
- Schleyer, M.; Bakker, E. Nucleotide sequence and 3’-end deletion studies indicate that the K+-uptake protein KUP from Escherichia coli is composed of a hydrophobic core linked to a large and partially essential hydrophilic C terminus. J. Bacteriol. 1993, 175, 6925–6931. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, M.; Klein, R.; Alexander-Bowman, S.; Rodríguez-Navarro, A. A potassium transporter of the yeast Schwanniomyces occidentalis homologous to the Kup system of Escherichia coli has a high concentrative capacity. EMBO J. 1995, 14, 3021–3027. [Google Scholar] [CrossRef] [PubMed]
- Quintero, F.; Blatt, M. A new family of K+ transporters from Arabidopsis that are conserved across phyla. FEBS Lett. 1997, 415, 206–211. [Google Scholar] [CrossRef]
- Li, Y.; Peng, L.; Xie, C.; Shi, X.; Dong, C.; Shen, Q.; Xu, Y. Genome-wide identification, characterization, and expression analyses of the HAK/KUP/KT potassium transporter gene family reveals their involvement in K+ deficient and abiotic stress responses in pear rootstock seedlings. Plant Growth Regul. 2018, 85, 187–198. [Google Scholar] [CrossRef]
- Li, W.; Xu, G.; Alli, A.; Yu, L. Plant HAK/KUP/KT K+ transporters: Function and regulation. Semin. Cell Dev. Biol. 2018, 74, 133–141. [Google Scholar] [CrossRef]
- Grabov, A. Plant KT/KUP/HAK potassium transporters: Single family—Multiple functions. Ann. Bot. 2007, 99, 1035–1041. [Google Scholar] [CrossRef]
- Gierth, M.; Maser, P. Potassium transporters in plants—Involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett. 2007, 581, 2348–2356. [Google Scholar] [CrossRef]
- Banuelos, M.A.; Garciadeblas, B.; Cubero, B.; Rodriguez-Navarro, A. Inventory and functional characterization of the HAK potassium transporters of rice. Plant Physiol. 2002, 130, 784–795. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Martinez, V.; Benito, B.; Rubio, F. Comparison between Arabidopsis and rice for main pathways of K+ and Na+ uptake by roots. Front. Plant Sci. 2016, 7, 992. [Google Scholar] [CrossRef] [PubMed]
- Rubio, F.; Esanta-Marita, G.; Rodrıíguez-Navarro, A. Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells. Physiol. Plant 2000, 109, 34–43. [Google Scholar] [CrossRef]
- Han, M.; Wu, W.; Wu, W.-H.; Wang, Y. Potassium transporter KUP7 is involved in K+ acquisition and translocation in Arabidopsis root under K+ limited conditions. Mol. Plant 2016, 9, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Hu, Q.; Luo, L.; Yang, T.; Zhang, S.; Hu, Y.; Yu, L.; Xu, G. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell Environ. 2015, 38, 2747–2765. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhang, S.; Hu, Y.; Wu, F.; Hu, Q.; Chen, G.; Cai, J.; Wu, T.; Moran, N.; Yu, L.; et al. The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol. 2014, 166, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Shen, L.; Shen, Z.; Jing, W.; Ge, H.; Zhao, J.; Zhang, W. The potassium transporter OsHAK21 functions in the maintenance of ion homeostasis and tolerance to salt stress in rice. Plant Cell Environ. 2015, 38, 2766–2779. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, C.; Gao, Z.; Zhang, Y.; Jiang, H.; Zhu, L.; Ren, D.; Yu, L.; Xu, G.; Qian, Q. OsHAK1, a high-affinity potassium transporter, positively regulates responses to drought stress in rice. Front. Plant Sci. 2017, 8, 1885. [Google Scholar] [CrossRef]
- Ensembl Plant Database. Available online: http://plants.ensembl.org (accessed on 9 July 2017).
- GSDS2.0: Gene Structure Display Server. Available online: http://gsds.cbi.pku.edu.cn/ (accessed on 10 June 2018).
- Zhang, Z.; Zhang, J.; Chen, Y.; Li, R.; Wang, H.; Wei, J. Genome-wide analysis and identification of HAK potassium transporter gene family in maize (Zea mays L.). Mol. Biol. Rep. 2012, 39, 8465–8473. [Google Scholar] [CrossRef]
- Wheat Exp: An Expression Database for Polyploid Wheat. Available online: https://wheat.pw.usda.gov/WheatExp/ (accessed on 7 November 2017).
- Ahn, S.J. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake. Plant Physiol. 2004, 134, 1135–1145. [Google Scholar] [CrossRef]
- Amrutha, R.; Sekhar, P.; Varshney, R.; Pbk, K. Genome-wide analysis and identification of genes related to potassium transpoter families in rice (Oryza sativa L.). Plant Sci. 2007, 172, 708–721. [Google Scholar] [CrossRef]
- Song, Z.Z.; Ma, R.J.; Yu, M.L. Genome-wide analysis and identification of KT/HAK/KUP potassium transporter gene family in peach (Prunus persica). Genet. Mol. Res. 2015, 14, 774–787. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qiao, L.; Bai, J.; Wang, P.; Duan, W.; Yuan, S.; Yuan, G.; Zhang, F.; Zhang, L.; Zhao, C. Genome-wide characterization of JASMONATE-ZIM DOMAIN transcription repressors in wheat (Triticum aestivum L.). BMC Genom. 2017, 18, 152. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Yang, Y.; Luo, W.; Yang, C.; Ding, P.; Liu, Y.; Qiao, L.; Chang, Z.; Geng, H.; Wang, P.; et al. Genome-wide identification and analysis of the MADS-box gene family in bread wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0181443. [Google Scholar] [CrossRef] [PubMed]
- Rodrígueznavarro, A. Potassium transport in fungi and plants. BBA Rev. Biomembr. 2000, 1469, 1–30. [Google Scholar] [CrossRef]
- Yang, Z.; Gao, Q.; Sun, C.; Li, W.; Gu, S.; Xu, C. Molecular evolution and functional divergence of HAK potassium transporter gene family in rice (Oryza sativa L.). J. Genet. Genom. 2009, 36, 161–172. [Google Scholar] [CrossRef]
- Rigas, S.; Debrosses, G.; Haralampidis, K.; Vicente-Agullo, F.; Feldmann, K.A.; Grabov, A.; Dolan, L.; Hatzopoulos, P. TRH1 encodes a potassium transporter required for tip growth in Arabidopsis root hairs. Plant Cell 2001, 13, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Elumalai, R.P. A mutation in the Arabidopsis KT2/KUP2 potassium transporter gene affects shoot cell expansion. Plant Cell Online 2002, 14, 119–131. [Google Scholar] [CrossRef]
- Maathuis, F.J. The role of monovalent cation transporters in plant responses to salinity. J. Exp. Bot. 2006, 57, 1137–1147. [Google Scholar] [CrossRef]
- Gierth, M.; Maser, P.; Schroeder, J.I. The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol. 2005, 137, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Nakayama, H.; Shinmyo, A.; Yoshida, K. Expression of OsHAK genes encoding potassium ion transporters in rice. Plant Biotechnol. 2008, 25, 241–245. [Google Scholar] [CrossRef]
- Cuin, T.A.; Bose, J.; Stefano, G.; Jha, D.; Tester, M.; Mancuso, S.; Shabala, S. Assessing the role of root plasma membrane and tonoplast Na+/H+ exchangers in salinity tolerance in wheat: In planta quantification methods. Plant Cell Environ. 2011, 34, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagamachi, K.; Tanaka, H.; Ohiraki, H.; Yamada, K.; Seo, S.U.; Abo, M.; et al. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell 2013, 25, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhou, Y.; Hong, S.; Xia, Z.; Cui, D.; Guo, J.; Xu, H.; Jiang, X. Functional characterization of a wheat NHX antiporter gene TaNHX2 that encodes a K+/H+ exchanger. PLoS ONE 2013, 8, e78098. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Conserved Domain Database (CDD) in NCBI. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/cdd (accessed on 11 June 2018).
- WoLF PSORT: Protein Subcellular Localization Prediction. Available online: https://wolfpsort.hgc.jp/ (accessed on 19 October 2017).
- TMHMM Server v. 2.0: Prediction of Transmembrane Helices in Proteins. Available online: http://www.cbs.dtu.dk/services/TMHMM/ (accessed on 18 October 2017).
- National Center for Biotechnology Information (NCBI). Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/ (accessed on 11 July 2017).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- URGI Wheat Genome Database. Available online: https://urgi.versailles.inra.fr/blast_iwgsc/blast.php (accessed on 3 November 2017).
- IWGSC CS RefSeq v1.0 Database. Available online: https://urgi.versailles.inra.fr/jbrowseiwgsc/gmod_jbrowse/?data=myData%2FIWGSC_RefSeq_v1.0 (accessed on 3 November 2017).
- MapInspect 1.0: Displays and Analyzes Genetic Linkage Maps. Available online: https://mapinspect.software.informer.com/ (accessed on 1 November 2017).
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- MEME: Multiple Em for Motif Elicitation, Version 4.11.4. Available online: http://memesuite.org/tools/meme (accessed on 20 June 2018).
- Anderson, J.; Huprikar, S.; Kochian, L.; Lucas, W.; Gaber, R. Functional expression of a probable Arabidopsis thaliana potassium channel in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1992, 89, 3736–3740. [Google Scholar] [CrossRef]
- Quintero, F.J.; Ohta, M.; Shi, H.; Zhu, J.K.; Pardo, J.M. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 9061–9066. [Google Scholar] [CrossRef]
- Zhou, Y.; Lai, Z.; Yin, X.; Yu, S.; Xu, Y.; Wang, X.; Cong, X.; Luo, Y.; Xu, H.; Jiang, X. Hyperactive mutant of a wheat plasma membrane Na+/H+ antiporter improves the growth and salt tolerance of transgenic tobacco. Plant Sci. 2016, 253, 176–186. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Ensembl ID | Amino Acid Length | TMS | Intron No. | Exon No. | S.L. | Gene Location (bp) | |
|---|---|---|---|---|---|---|---|---|
| Start | End | |||||||
| TaHAK1a-4BL | TRIAE_CS42_4BL_TGACv1_321685_AA1064100.1 | 784 | 11 | 7 | 8 | PM | 600657723 | 600654569 |
| TaHAK1a-4DL | TRIAE_CS42_4DL_TGACv1_343636_AA1137540.1 | 778 | 11 | 8 | 9 | PM | 476306655 | 476302887 |
| TaHAK1b-2BL | TRIAE_CS42_2BL_TGACv1_129472_AA0385230.2 | 768 | 11 | 8 | 9 | PM | 436243088 | 436237762 |
| TaHAK1b-2DL | TRIAE_CS42_2DL_TGACv1_158352_AA0516430.1 | 776 | 11 | 8 | 9 | PM | 367507729 | 367502225 |
| TaHAK1b-2A | TRIAE_CS42_U_TGACv1_641460_AA2095620.1 | 772 | 11 | 8 | 9 | PM | 497657505 | 497652193 |
| TaHAK1c-2BL | TRIAE_CS42_2BL_TGACv1_129437_AA0383630.1 | 774 | 10 | 8 | 9 | PM | 436956667 | 436962029 |
| TaHAK2-3AL | TRIAE_CS42_3AL_TGACv1_194420_AA0632700.2 | 784 | 13 | 8 | 9 | PM | 695496006 | 695500670 |
| TaHAK2-3DL | TRIAE_CS42_3DL_TGACv1_250243_AA0864920.1 | 788 | 13 | 9 | 10 | PM | 559072825 | 559076602 |
| TaHAK3-1BL | TRIAE_CS42_1BL_TGACv1_033126_AA0137190.1 | 818 | 11 | 6 | 7 | PM | 559097952 | 559092195 |
| TaHAK3-1DL | TRIAE_CS42_1DL_TGACv1_062044_AA0207990.1 | 817 | 11 | 6 | 7 | PM | 414287464 | 414282613 |
| TaHAK4-7AS | TRIAE_CS42_7AS_TGACv1_570480_AA1836390.3 | 686 | 11 | 4 | 5 | PM | 36648278 | 36651604 |
| TaHAK4-7DS | TRIAE_CS42_7DS_TGACv1_622074_AA2032320.3 | 725 | 11 | 4 | 5 | PM | 36992201 | 36998396 |
| TaHAK5-3AL | TRIAE_CS42_3AL_TGACv1_193587_AA0613950.2 | 788 | 11 | 9 | 10 | PM | 686834648 | 686837566 |
| TaHAK5-3B | TRIAE_CS42_3B_TGACv1_222217_AA0760010.1 | 782 | 12 | 8 | 9 | PM | 729204607 | 729207978 |
| TaHAK5-3DL | TRIAE_CS42_3DL_TGACv1_249200_AA0841080.1 | 764 | 11 | 9 | 10 | PM | 549623301 | 549628488 |
| TaHAK6-3AL | TRIAE_CS42_3AL_TGACv1_195362_AA0648570.1 | 732 | 11 | 5 | 6 | PM | 689132246 | 689135499 |
| TaHAK6-3D | TRIAE_CS42_U_TGACv1_646122_AA2146080.1 | 732 | 10 | 5 | 6 | PM | 552878679 | 552881787 |
| TaHAK7-2AS | TRIAE_CS42_2AS_TGACv1_114941_AA0370050.1 | 778 | 13 | 8 | 9 | PM | 66798807 | 66805468 |
| TaHAK7-2DS | TRIAE_CS42_2DS_TGACv1_179048_AA0603870.1 | 778 | 13 | 8 | 9 | PM | 67562036 | 67555857 |
| TaHAK9-2AS | TRIAE_CS42_2AS_TGACv1_112740_AA0344400.1 | 782 | 11 | 6 | 7 | PM | 59558656 | 59554035 |
| TaHAK9-2DS | TRIAE_CS42_2DS_TGACv1_177265_AA0571140.1 | 780 | 11 | 6 | 7 | PM | 58776915 | 58781534 |
| TaHAK10-7AL | TRIAE_CS42_7AL_TGACv1_556774_AA1770580.1 | 829 | 11 | 8 | 9 | PM | 571701040 | 571706004 |
| TaHAK10-7BL | TRIAE_CS42_7BL_TGACv1_578444_AA1895080.2 | 825 | 11 | 8 | 9 | PM | 532275080 | 532279899 |
| TaHAK10-7DL | TRIAE_CS42_7DL_TGACv1_604392_AA1997720.1 | 827 | 11 | 8 | 9 | PM | 504174129 | 504179023 |
| TaHAK11-2DL | TRIAE_CS42_2DL_TGACv1_159022_AA0530870.1 | 792 | 14 | 7 | 8 | PM | 533132266 | 533137640 |
| TaHAK12-1AS | TRIAE_CS42_1AS_TGACv1_020203_AA0075600.3 | 790 | 14 | 8 | 9 | PM | 29186381 | 29192163 |
| TaHAK13-7AL | TRIAE_CS42_7AL_TGACv1_557049_AA1775770.1 | 803 | 11 | 8 | 9 | PM | 665751743 | 665743564 |
| TaHAK13-7BL | TRIAE_CS42_7BL_TGACv1_577019_AA1862890.1 | 750 | 11 | 9 | 10 | PM | 637233552 | 637228550 |
| TaHAK13-7DL | TRIAE_CS42_7DL_TGACv1_603280_AA1979940.1 | 803 | 11 | 8 | 9 | PM | 575510919 | 575502903 |
| TaHAK16-6AL | TRIAE_CS42_6AL_TGACv1_471561_AA1510890.1 | 806 | 11 | 9 | 10 | PM | 526114436 | 526120099 |
| TaHAK16-6BL | TRIAE_CS42_6BL_TGACv1_502060_AA1622780.1 | 802 | 11 | 9 | 10 | PM | 574459052 | 574465508 |
| TaHAK16-6DL | TRIAE_CS42_6DL_TGACv1_527466_AA1704460.1 | 805 | 11 | 9 | 10 | PM | 384217110 | 384223492 |
| TaHAK17a-5AL | TRIAE_CS42_5AL_TGACv1_374155_AA1191940.2 | 719 | 12 | 7 | 8 | PM | 448614349 | 448619914 |
| TaHAK17a-5BL | TRIAE_CS42_5BL_TGACv1_405525_AA1329450.1 | 719 | 12 | 7 | 8 | PM | 408955185 | 408959389 |
| TaHAK17a-5DL | TRIAE_CS42_5DL_TGACv1_433406_AA1412260.1 | 719 | 12 | 7 | 8 | PM | 348479742 | 348483109 |
| TaHAK17b-5DL | TRIAE_CS42_5DL_TGACv1_434380_AA1434940.1 | 712 | 11 | 7 | 8 | PM | 348346780 | 348352011 |
| TaHAK18-5BL | TRIAE_CS42_5BL_TGACv1_407556_AA1357920.1 | 785 | 14 | 8 | 9 | PM | 546832103 | 546826324 |
| TaHAK18-5DL | TRIAE_CS42_5DL_TGACv1_433145_AA1403590.1 | 785 | 14 | 8 | 9 | PM | 447886169 | 447880484 |
| TaHAK19a-6AS | TRIAE_CS42_6AS_TGACv1_488672_AA1576040.1 | 735 | 11 | 5 | 6 | PM | 33582679 | 33586043 |
| TaHAK19a-6BS | TRIAE_CS42_6BS_TGACv1_514904_AA1665660.1 | 737 | 11 | 5 | 6 | PM | 61222797 | 61226427 |
| TaHAK19a-6D | TRIAE_CS42_U_TGACv1_644308_AA2138720.1 | 734 | 11 | 7 | 8 | PM | 28943744 | 28939705 |
| TaHAK19b-6BS | TRIAE_CS42_6BS_TGACv1_513620_AA1645910.1 | 736 | 11 | 5 | 6 | PM | 60865245 | 60860866 |
| TaHAK19b-6D | TRIAE_CS42_U_TGACv1_644372_AA2139050.1 | 738 | 11 | 6 | 7 | PM | 28998084 | 29001654 |
| TaHAK19c-3B | TRIAE_CS42_3B_TGACv1_221313_AA0736890.1 | 745 | 10 | 6 | 7 | PM | 765005247 | 765001714 |
| TaHAK19c-3DL | TRIAE_CS42_3DL_TGACv1_251389_AA0881060.1 | 744 | 10 | 6 | 7 | PM | 574916450 | 574919986 |
| TaHAK22-2AL | TRIAE_CS42_2AL_TGACv1_096842_AA0321670.1 | 875 | 12 | 2 | 3 | PM | 392724188 | 392729472 |
| TaHAK22-2BL | TRIAE_CS42_2BL_TGACv1_133206_AA0441840.1 | 873 | 12 | 2 | 3 | PM | 394364701 | 394359570 |
| TaHAK22-2DL | TRIAE_CS42_2DL_TGACv1_158627_AA0523560.1 | 878 | 12 | 2 | 3 | PM | 325365084 | 325359910 |
| TaHAK23-5AL | TRIAE_CS42_5AL_TGACv1_377379_AA1246300.1 | 814 | 10 | 9 | 10 | PM | 403355442 | 403350814 |
| TaHAK23-5BL | TRIAE_CS42_5BL_TGACv1_406332_AA1344100.3 | 916 | 12 | 8 | 9 | PM | 355690579 | 355685671 |
| TaHAK23-5DL | TRIAE_CS42_5DL_TGACv1_435669_AA1453280.1 | 898 | 12 | 8 | 9 | PM | 309789789 | 309784581 |
| TaHAK24-7AS | TRIAE_CS42_7AS_TGACv1_569122_AA1807860.1 | 772 | 11 | 8 | 9 | PM | 172470095 | 172464552 |
| TaHAK24-7BS | TRIAE_CS42_7BS_TGACv1_593324_AA1950450.2 | 771 | 13 | 8 | 9 | PM | 135583478 | 135578011 |
| TaHAK24-7DS | TRIAE_CS42_7DS_TGACv1_621622_AA2021390.1 | 771 | 11 | 8 | 9 | PM | 169916315 | 169910689 |
| TaHAK25-6AL | TRIAE_CS42_6AL_TGACv1_471048_AA1501690.1 | 769 | 12 | 8 | 9 | PM | 523950482 | 523944232 |
| TaHAK25-6BL | TRIAE_CS42_6BL_TGACv1_501501_AA1618220.2 | 768 | 13 | 8 | 9 | PM | 570980883 | 570976570 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, X.; Liu, X.; Mao, W.; Zhang, X.; Chen, S.; Zhan, K.; Bi, H.; Xu, H. Genome-Wide Identification and Analysis of HAK/KUP/KT Potassium Transporters Gene Family in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2018, 19, 3969. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123969
Cheng X, Liu X, Mao W, Zhang X, Chen S, Zhan K, Bi H, Xu H. Genome-Wide Identification and Analysis of HAK/KUP/KT Potassium Transporters Gene Family in Wheat (Triticum aestivum L.). International Journal of Molecular Sciences. 2018; 19(12):3969. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123969
Chicago/Turabian StyleCheng, Xiyong, Xiaodan Liu, Weiwei Mao, Xurui Zhang, Shulin Chen, Kehui Zhan, Huihui Bi, and Haixia Xu. 2018. "Genome-Wide Identification and Analysis of HAK/KUP/KT Potassium Transporters Gene Family in Wheat (Triticum aestivum L.)" International Journal of Molecular Sciences 19, no. 12: 3969. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123969