Activation of TRPV1 and TRPM8 Channels in the Larynx and Associated Laryngopharyngeal Regions Facilitates the Swallowing Reflex

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
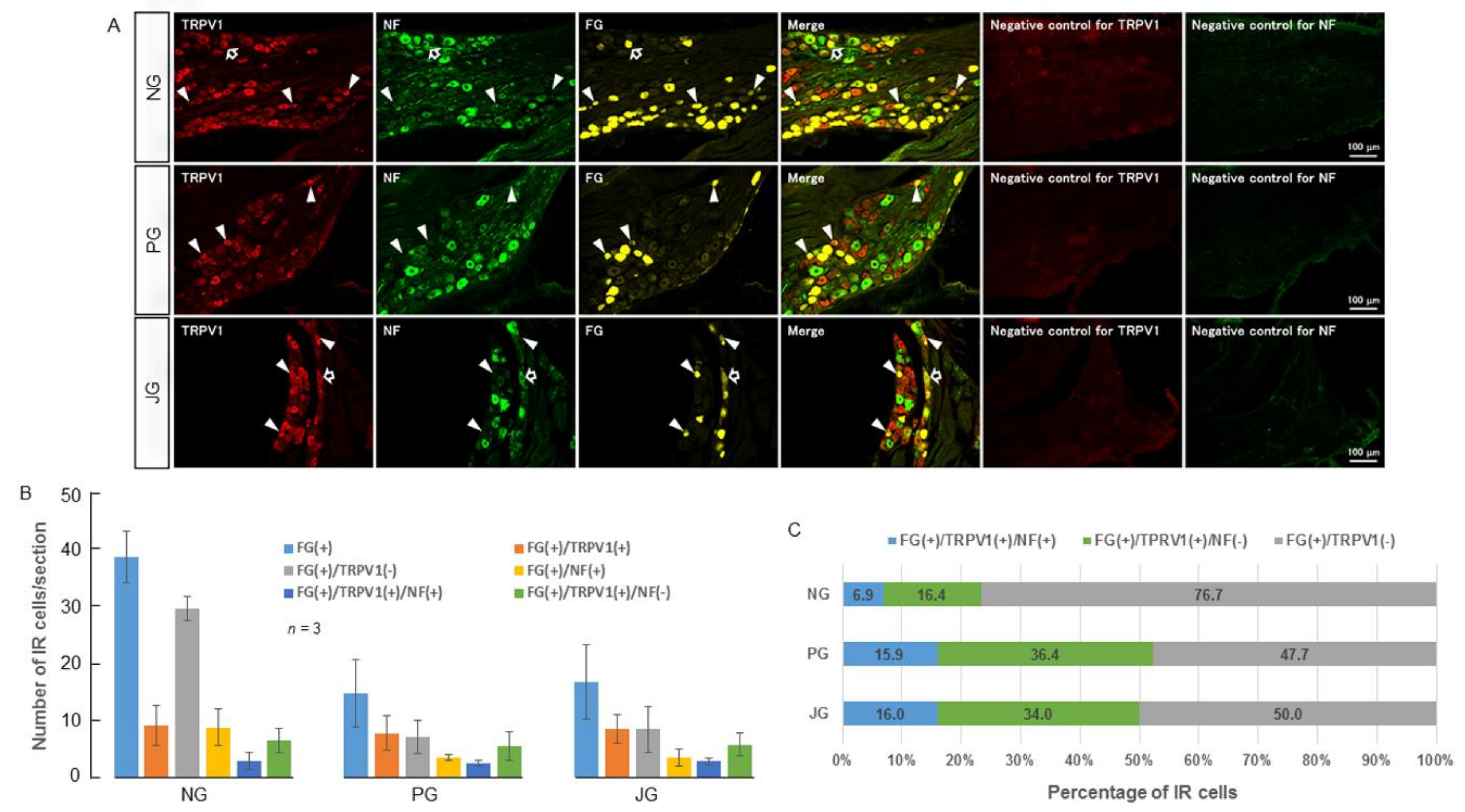
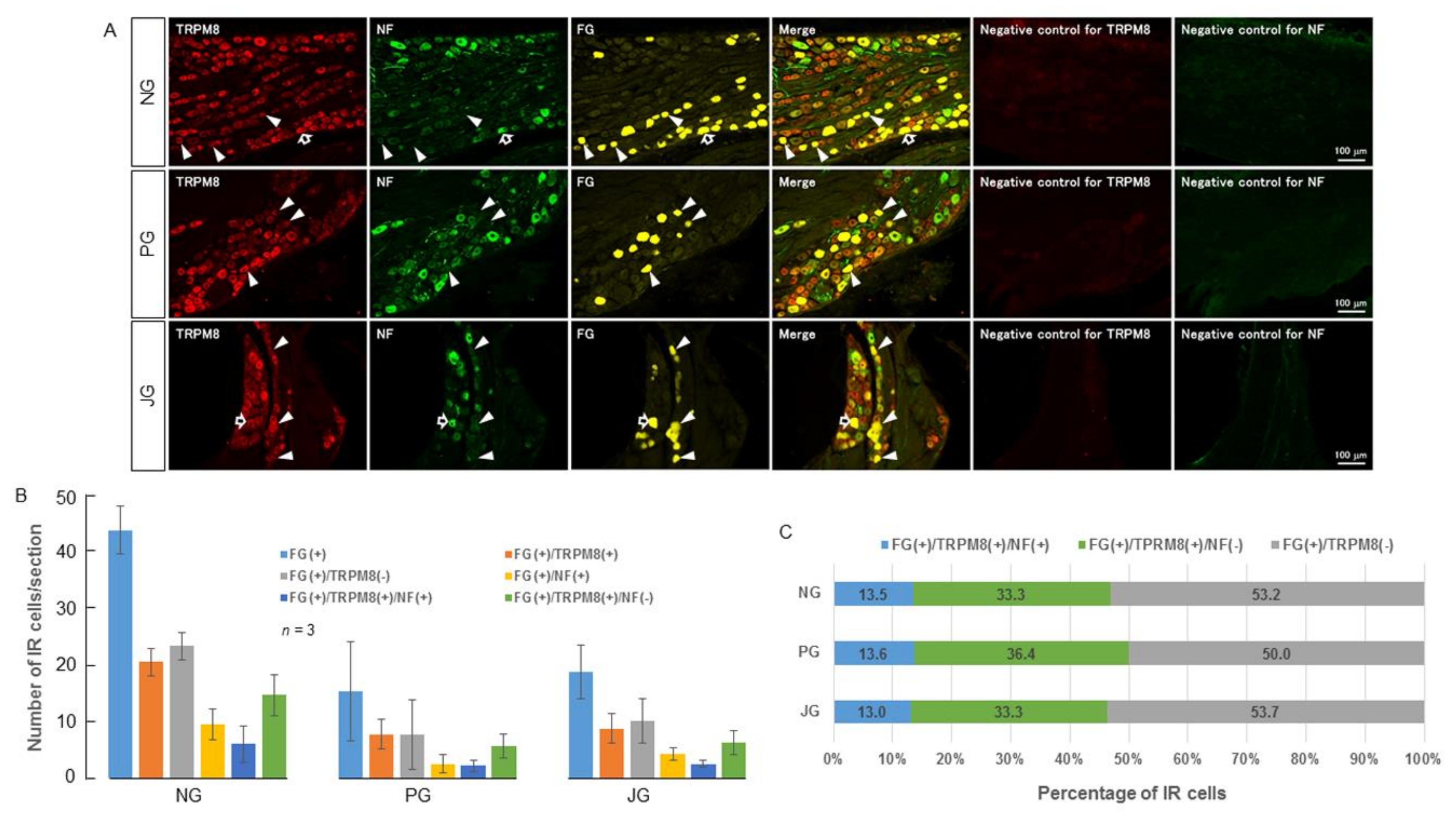
2.1. TRPV1 and TRPM8 Expression in the NPJc
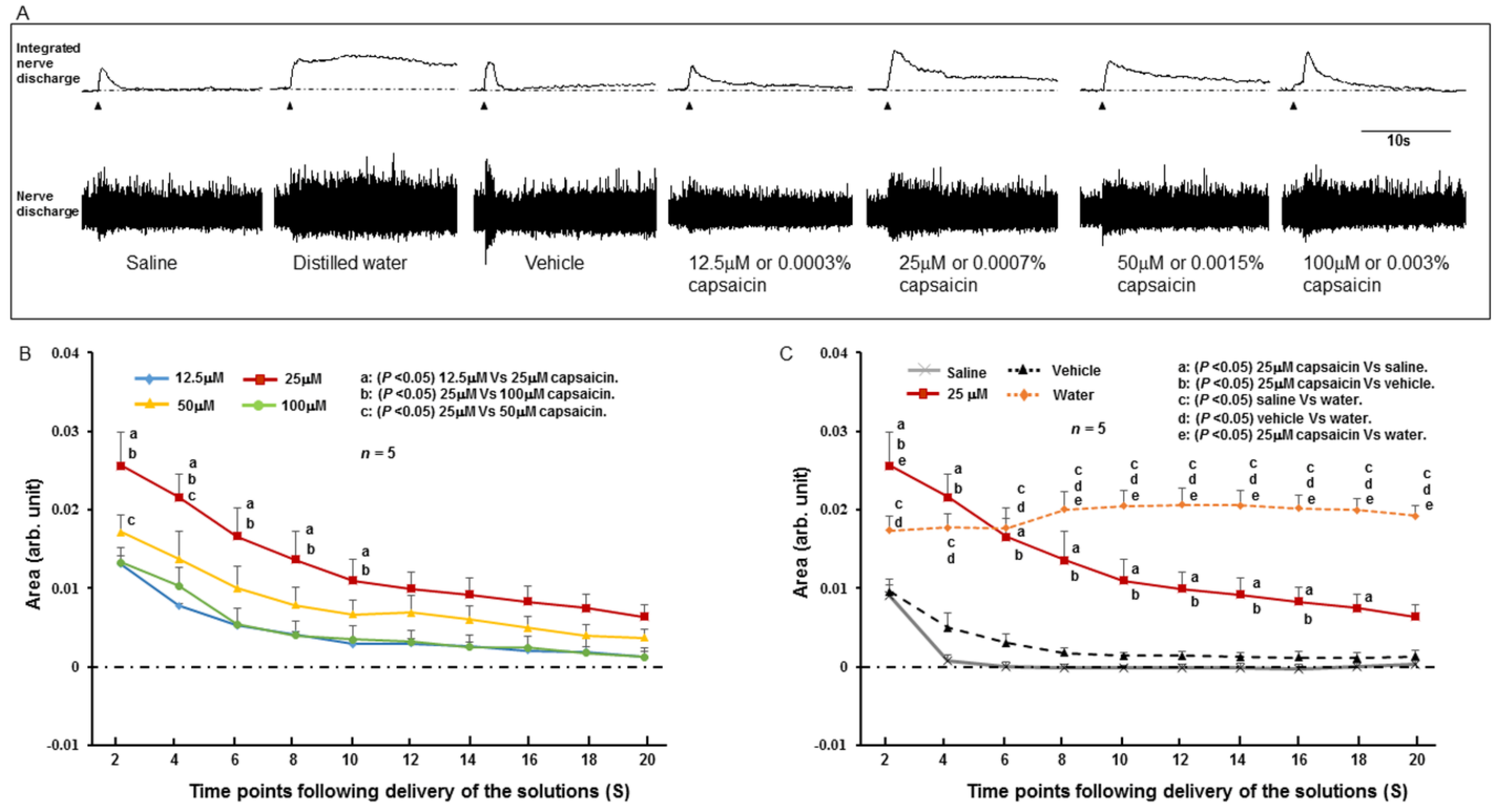
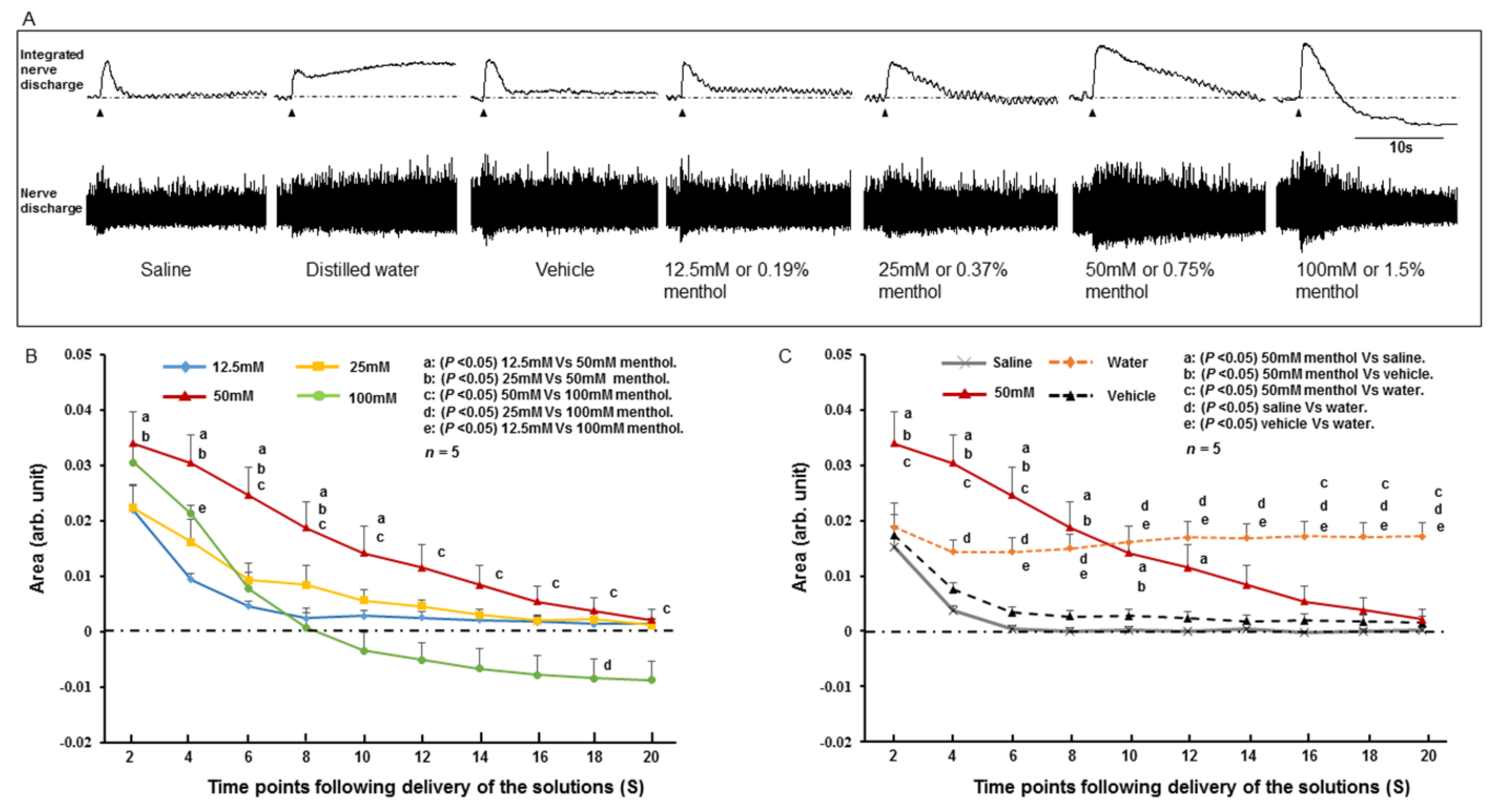
2.2. SLN Response to Stimulating Solutions
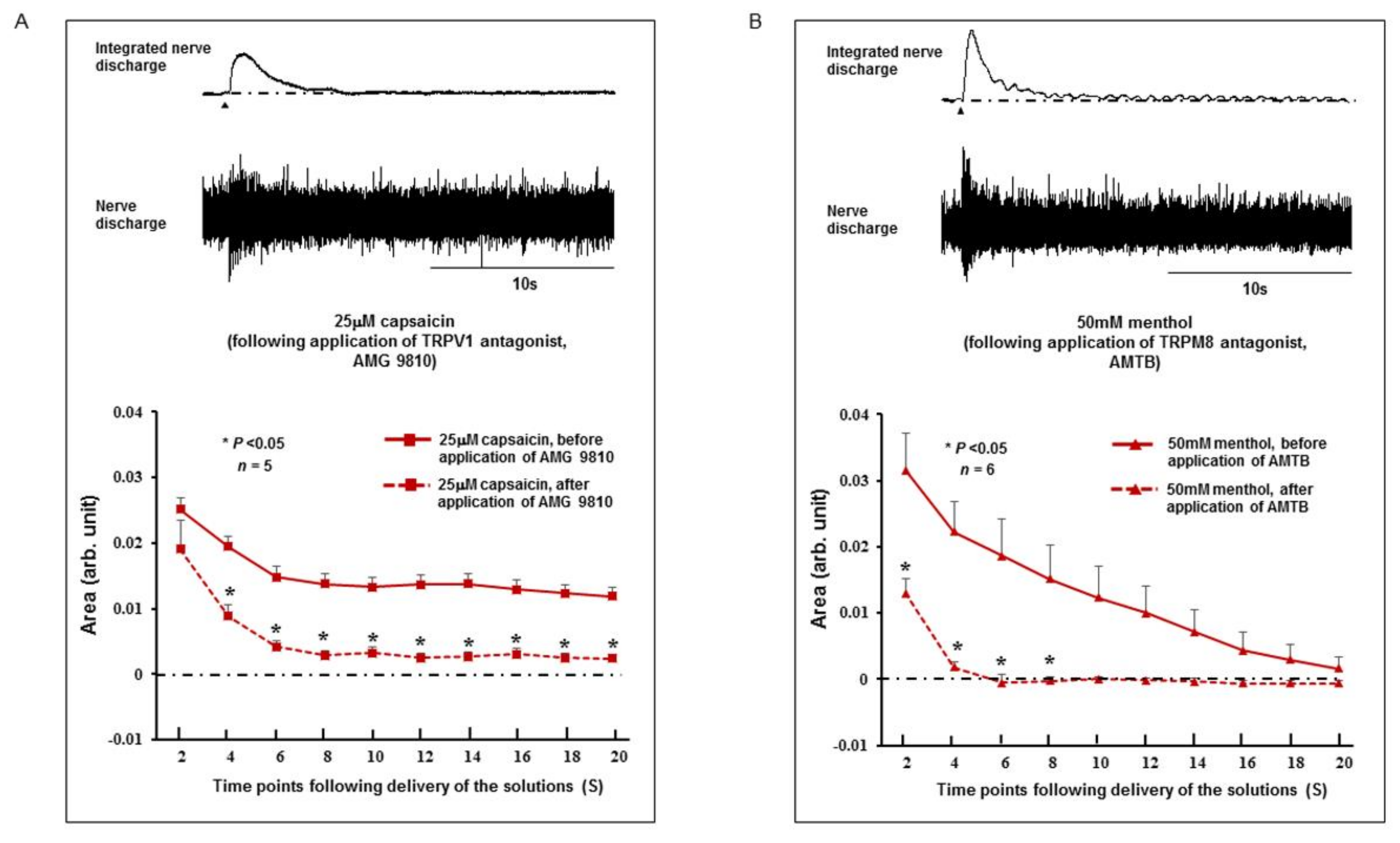
2.3. Effect of TRPV1 and TRPM8 Antagonists on the SLN Response
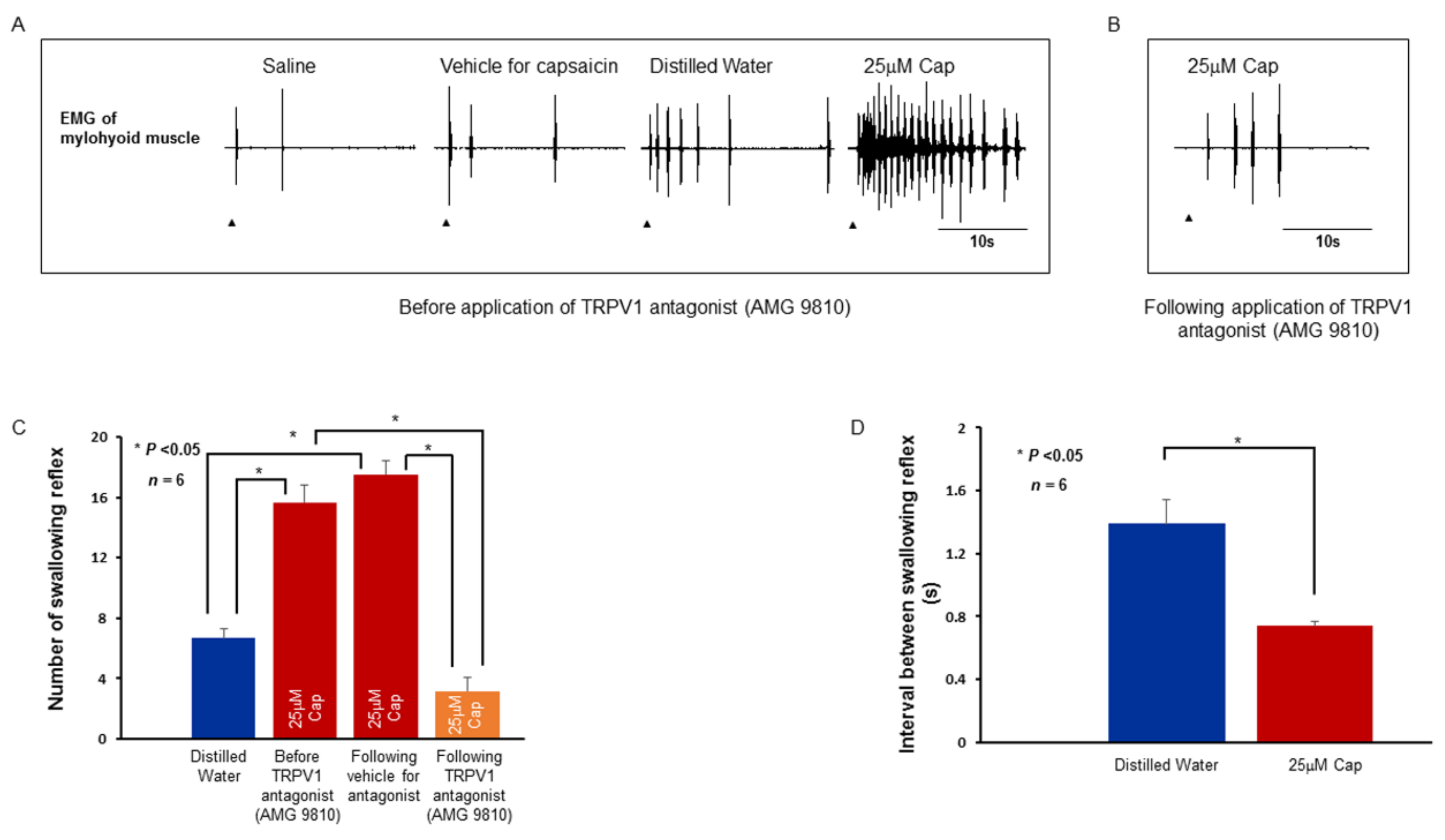
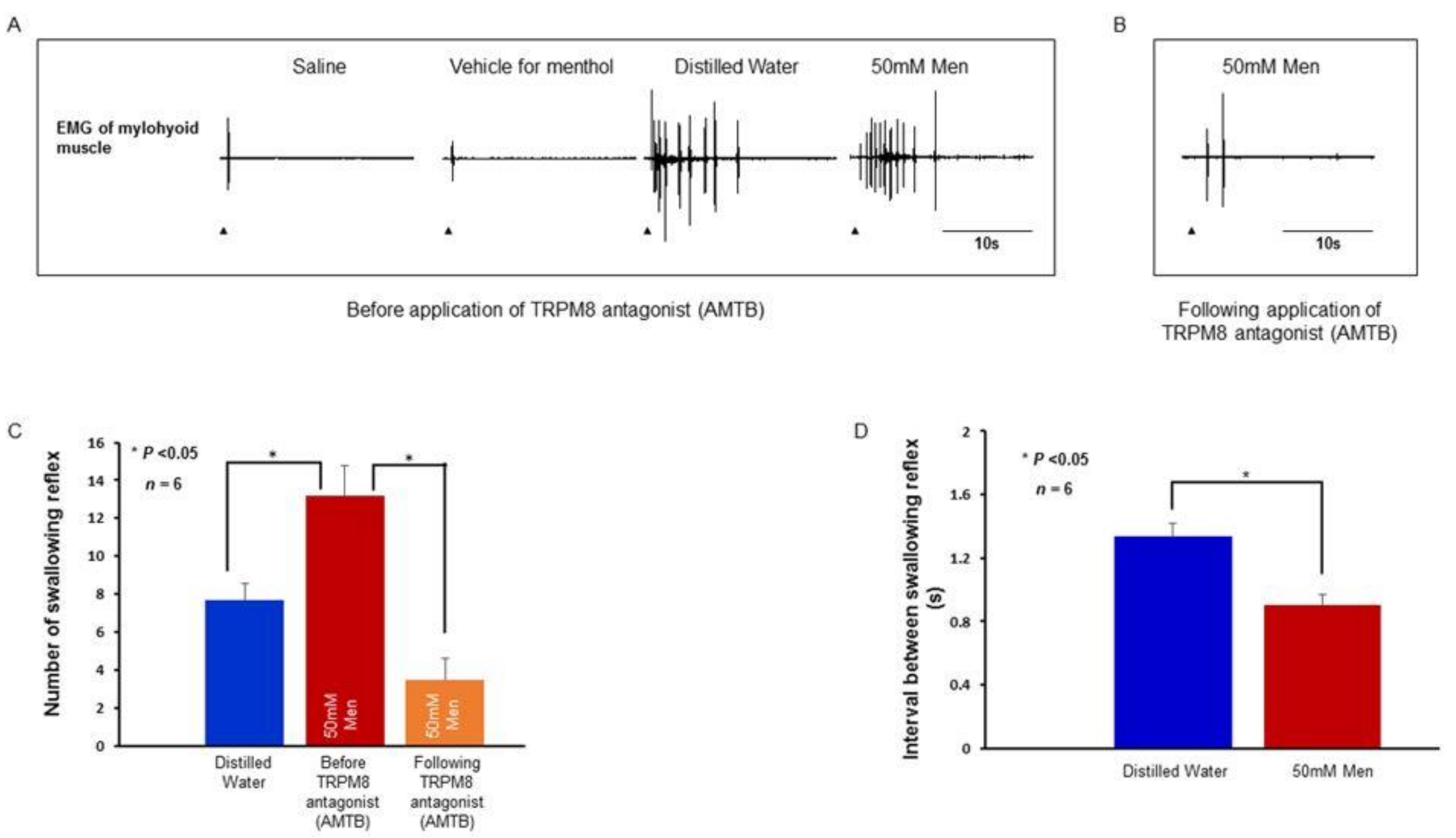
2.4. Swallowing Reflex Evoked by Stimulating Solutions and Effect of TRPV1 and TRPM8 Antagonists on the Reflex
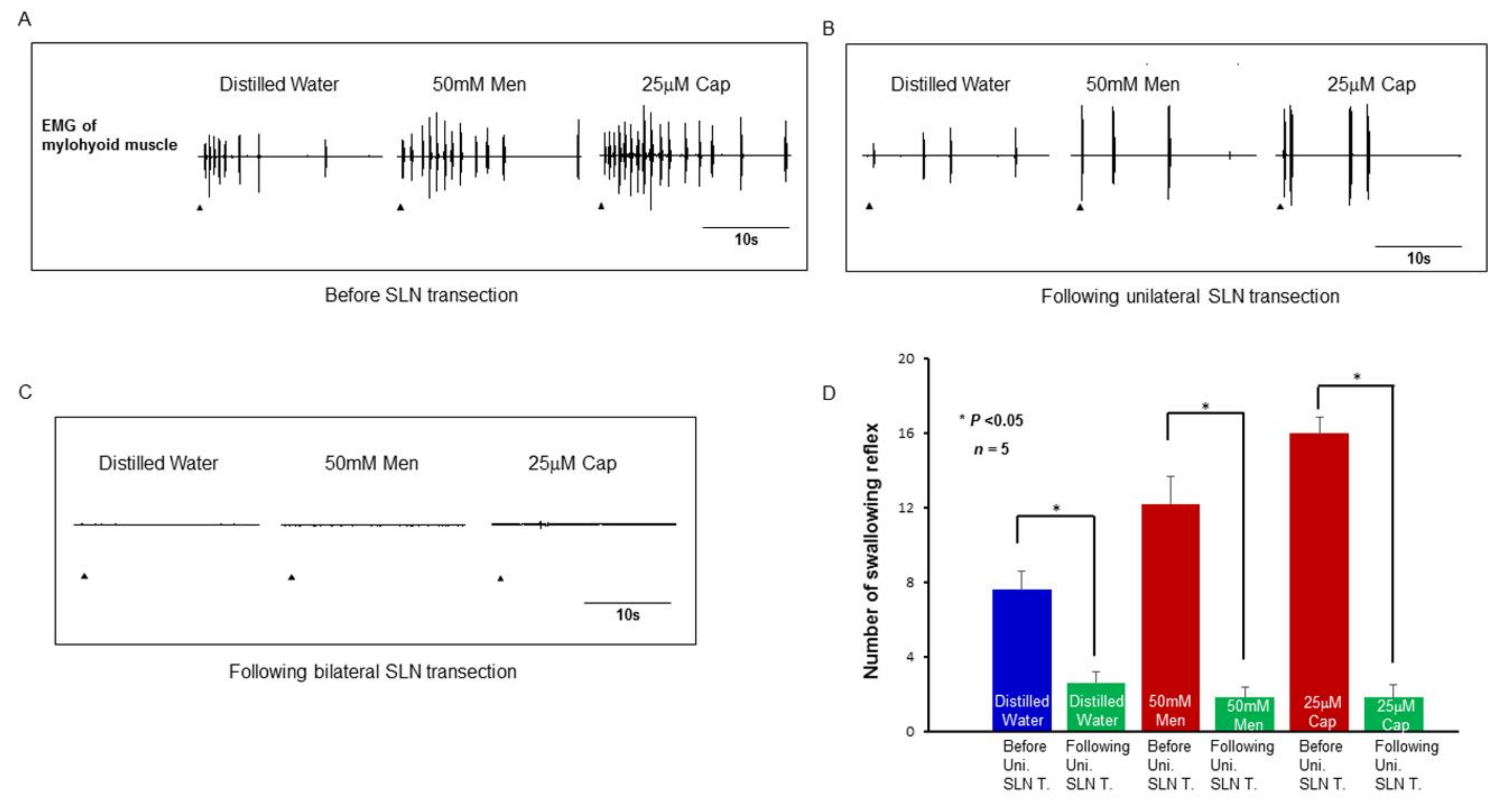
2.5. Effect of SLN Transection on the Evoked Swallowing Reflex
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Animals
4.3. Surgical Preparation
4.4. Transection of IX-ph, X-ph, and Recurrent Laryngeal Nerves
4.5. Exposing the SLN and Recording of SLN Activity
4.6. Recording of Swallowing Reflexes
4.7. Stimulating Solutions and Delivery of the Solutions
4.8. TRPM8 and TRPV1 Antagonists
4.9. Immunohistochemistry
4.10. Data Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Robbins, J. Upper aerodigestive tract neurofunctional mechanisms: Lifelong evolution and exercise. Head Neck 2011, 33 (Suppl. 1), S30–S36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludlow, C.L. Central Nervous System Control of Voice and Swallowing. J. Clin. Neurophysiol. 2015, 32, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takatsuji, H.; Zakir, H.M.; Mostafeezur, R.M.; Saito, I.; Yamada, Y.; Yamamura, K.; Kitagawa, J. Induction of the Swallowing Reflex by Electrical Stimulation of the Posterior Oropharyngeal Region in Awake Humans. Dysphagia 2012, 27, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, K.; Kitagawa, J.; Kurose, M.; Sugino, S.; Takatsuji, H.; Mostafeezur, R.M.; Zakir, H.M.; Yamada, Y. Neural mechanisms of swallowing and effects of taste and other stimuli on swallow initiation. Biol. Pharm. Bull. 2010, 33, 1786–1790. [Google Scholar] [CrossRef] [PubMed]
- Mostafeezur, R.M.; Zakir, H.M.; Takatsuji, H.; Yamada, Y.; Yamamura, K.; Kitagawa, J. Cannabinoids facilitate the swallowing reflex elicited by the superior laryngeal nerve stimulation in rats. PLoS ONE 2012, 7, e50703. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, J.; Shingai, T.; Takahashi, Y.; Yamada, Y. Pharyngeal branch of the glossopharyngeal nerve plays a major role in reflex swallowing from the pharynx. Am. J. Physiol. 2002, 282, R1342–R1347. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, W.J. Role of the pharyngeal plexus in initiation of swallowing. Am. J. Physiol. 1971, 221, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Storey, A.T. Laryngeal initiation of swallowing. Exp. Neurol. 1968, 20, 359–365. [Google Scholar] [CrossRef]
- Storey, A.T. A functional analysis of sensory units innervating epiglottis and larynx. Exp. Neurol. 1968, 20, 366–383. [Google Scholar] [CrossRef]
- Sanders, I.; Mu, L. Anatomy of the human internal superior laryngeal nerve. Anat. Rec. 1998, 252, 646–656. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Tanaka, Y.; Hirano, M.; Nakashima, T. Sensory innervation of the pharynx and larynx. Am. J. Med. 2000, 108 (Suppl 4a), 51S–61S. [Google Scholar] [CrossRef]
- Wank, M.; Neuhuber, W.L. Local differences in vagal afferent innervation of the rat esophagus are reflected by neurochemical differences at the level of the sensory ganglia and by different brainstem projections. J. Comp. Neurol. 2001, 435, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Kajii, Y.; Shingai, T.; Kitagawa, J.; Takahashi, Y.; Taguchi, Y.; Noda, T.; Yamada, Y. Sour taste stimulation facilitates reflex swallowing from the pharynx and larynx in the rat. Physiol. Behav. 2002, 77, 321–325. [Google Scholar] [CrossRef]
- Kitagawa, J.; Takahashi, Y.; Matsumoto, S.; Shingai, T. Response properties of the pharyngeal branch of the glossopharyngeal nerve for umami taste in mice and rats. Neurosci. Lett. 2007, 417, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Ohkuri, T.; Horio, N.; Stratford, J.M.; Finger, T.E.; Ninomiya, Y. Residual chemoresponsiveness to acids in the superior laryngeal nerve in “taste-blind” (P2X2/P2X3 double-KO) mice. Chem. Senses 2012, 37, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.M.; Stedman, H.M.; Mistretta, C.M. Superior laryngeal nerve response patterns to chemical stimulation of sheep epiglottis. Brain Res. 1983, 276, 81–93. [Google Scholar] [CrossRef]
- Smith, D.V.; Hanamori, T. Organization of gustatory sensitivities in hamster superior laryngeal nerve fibers. J. Neurophysiol. 1991, 65, 1098–1114. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E.; Runnels, L.W.; Strubing, C. The TRP ion channel family. Nat. Rev. 2001, 2, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed]
- Hondoh, A.; Ishida, Y.; Ugawa, S.; Ueda, T.; Shibata, Y.; Yamada, T.; Shikano, M.; Murakami, S.; Shimada, S. Distinct expression of cold receptors (TRPM8 and TRPA1) in the rat nodose-petrosal ganglion complex. Brain Res. 2010, 1319, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Fujita, M.; Kano, M.; Hosokawa, H.; Kondo, T.; Suzuki, T.; Kasahara, E.; Shoji, N.; Sasano, T.; Ichikawa, H. The distribution of transient receptor potential melastatin-8 in the rat soft palate, epiglottis, and pharynx. Cell. Mol. Neurobiol. 2013, 33, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, R.; Sato, T.; Yajima, T.; Kano, M.; Suzuki, T.; Ichikawa, H. The distribution of TRPV1 and TRPV2 in the rat pharynx. Cell. Mol. Neurobiol. 2013, 33, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, T.; Takumida, M.; Hirakawa, K.; Tatsukawa, T.; Ishibashi, T. Localization of transient receptor potential vanilloid (TRPV) in the human larynx. Acta Oto-Laryngol. 2009, 129, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Berdugo, D.; Rofes, L.; Farre, R.; Casamitjana, J.F.; Enrique, A.; Chamizo, J.; Padron, A.; Navarro, X.; Clave, P. Localization and expression of TRPV1 and TRPA1 in the human oropharynx and larynx. Neurogastroenterol. Motil. 2016, 28, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Jean, A. Brain stem control of swallowing: Neuronal network and cellular mechanisms. Physiol. Rev. 2001, 81, 929–969. [Google Scholar] [CrossRef] [PubMed]
- Dubner, R.; Sessle, B.J.; Storey, A.T. Pharynx and Larynx. In The Neural Basis of Oral and Facial Function; Dubner, R., Ed.; Springer: New York, NY, USA, 1978; pp. 175–207. ISBN 978-1-4757-1682-5. [Google Scholar]
- Mei, N.; Condamin, M.; Boyer, A. The composition of the vagus nerve of the cat. Cell Tissue Res. 1980, 209, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, S.B.; Undem, B.J. Vagal Afferent Innervation of the Airways in Health and Disease. Physiol. Rev. 2016, 96, 975–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hishida, N.; Tsubone, H.; Sugano, S. Fiber composition of the superior laryngeal nerve in rats and guinea pigs. J. Vet. Med. Sci. 1997, 59, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T. The swallowing reflex and its significance as an airway defensive reflex. Front. Physiol. 2013, 3, 489. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Palmer, J.B. Coordination of Mastication, Swallowing and Breathing. Jpn. Dent. Sci. Rev. 2009, 45, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R.; Costa, A.G.; Morais, H.C.; Cavalcante, T.F.; Lopes, M.V.; Araujo, T.L. Clinical factors predicting risk for aspiration and respiratory aspiration among patients with Stroke. Rev. Latino-Am. Enferm. 2015, 23, 216–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rofes, L.; Arreola, V.; Almirall, J.; Cabre, M.; Campins, L.; Garcia-Peris, P.; Speyer, R.; Clave, P. Diagnosis and management of oropharyngeal Dysphagia and its nutritional and respiratory complications in the elderly. Gastroenterol. Res. Pract. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, T.; Takahashi, H.; Ebihara, S.; Okazaki, T.; Sasaki, T.; Watando, A.; Nemoto, M.; Sasaki, H. Capsaicin troche for swallowing dysfunction in older people. J. Am. Geriatr. Soc. 2005, 53, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, T.; Sekizawa, K.; Nakazawa, H.; Sasaki, H. Capsaicin and swallowing reflex. Lancet 1993, 341, 432. [Google Scholar] [CrossRef]
- Nakato, R.; Manabe, N.; Shimizu, S.; Hanayama, K.; Shiotani, A.; Hata, J.; Haruma, K. Effects of Capsaicin on Older Patients with Oropharyngeal Dysphagia: A Double-Blind, Placebo-Controlled, Crossover Study. Digestion 2017, 95, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Rofes, L.; Arreola, V.; Martin, A.; Clave, P. Natural capsaicinoids improve swallow response in older patients with oropharyngeal dysphagia. Gut 2012, 62, 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Berdugo, D.; Rofes, L.; Arreola, V.; Martin, A.; Molina, L.; Clave, P. A comparative study on the therapeutic effect of TRPV1, TRPA1, and TRPM8 agonists on swallowing dysfunction associated with aging and neurological diseases. Neurogastroenterol. Motil. 2018, 30, e13185. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, T.; Ebihara, S.; Watando, A.; Okazaki, T.; Asada, M.; Ohrui, T.; Yamaya, M.; Arai, H. Effects of menthol on the triggering of the swallowing reflex in elderly patients with dysphagia. Br. J. Clin. Pharmacol. 2006, 62, 369–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Shingai, T.; Saito, I.; Yamamura, K.; Yamada, Y.; Kitagawa, J. Facilitation of the swallowing reflex with bilateral afferent input from the superior laryngeal nerve. Neurosci. Lett. 2014, 562, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, J.; Nakagawa, K.; Hasegawa, M.; Iwakami, T.; Shingai, T.; Yamada, Y.; Iwata, K. Facilitation of reflex swallowing from the pharynx and larynx. J. Oral Sci. 2009, 51, 167–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudioso, C.; Hao, J.; Martin-Eauclaire, M.F.; Gabriac, M.; Delmas, P. Menthol pain relief through cumulative inactivation of voltage-gated sodium channels. Pain 2012, 153, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, M.; Uchida, K.; Suzuki, Y.; Matsui, H.; Shimada, T.; Fujita, F.; Tominaga, M. Reciprocal effects of capsaicin and menthol on thermosensation through regulated activities of TRPV1 and TRPM8. J. Physiol. Sci. 2016, 66, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.H.; Sawyer, C.M.; Takechi, K.; Davoodi, A.; Ivanov, M.A.; Carstens, M.I.; Carstens, E. Topical hindpaw application of L-menthol decreases responsiveness to heat with biphasic effects on cold sensitivity of rat lumbar dorsal horn neurons. Neuroscience 2012, 219, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.H.; Sawyer, C.M.; Carstens, M.I.; Tsagareli, M.G.; Tsiklauri, N.; Carstens, E. Topical application of L-menthol induces heat analgesia, mechanical allodynia, and a biphasic effect on cold sensitivity in rats. Behav. Brain Res. 2010, 212, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Qin, F. Functional control of cold- and menthol-sensitive TRPM8 ion channels by phosphatidylinositol 4,5-bisphosphate. J. Neurosci. 2005, 25, 1674–1681. [Google Scholar] [CrossRef] [PubMed]
- Rohacs, T.; Lopes, C.M.; Michailidis, I.; Logothetis, D.E. PI(4,5)P2 regulates the activation and desensitization of TRPM8 channels through the TRP domain. Nat. Neurosci. 2005, 8, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Daniels, R.L.; Takashima, Y.; McKemy, D.D. Activity of the neuronal cold sensor TRPM8 is regulated by phospholipase C via the phospholipid phosphoinositol 4,5-bisphosphate. J. Biol. Chem. 2009, 284, 1570–1582. [Google Scholar] [CrossRef] [PubMed]
- Sarria, I.; Ling, J.; Zhu, M.X.; Gu, J.G. TRPM8 acute desensitization is mediated by calmodulin and requires PIP(2): Distinction from tachyphylaxis. J. Neurophysiol. 2011, 106, 3056–3066. [Google Scholar] [CrossRef] [PubMed]
- Vyklicky, L.; Novakova-Tousova, K.; Benedikt, J.; Samad, A.; Touska, F.; Vlachova, V. Calcium-dependent desensitization of vanilloid receptor TRPV1: A mechanism possibly involved in analgesia induced by topical application of capsaicin. Physiol. Res. 2008, 57 (Suppl. 3), S59–S68. [Google Scholar] [PubMed]
- Numazaki, M.; Tominaga, T.; Takeuchi, K.; Murayama, N.; Toyooka, H.; Tominaga, M. Structural determinant of TRPV1 desensitization interacts with calmodulin. Proc. Natl. Acad. Sci. USA 2003, 100, 8002–8006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohapatra, D.P.; Nau, C. Desensitization of capsaicin-activated currents in the vanilloid receptor TRPV1 is decreased by the cyclic AMP-dependent protein kinase pathway. J. Biol. Chem. 2003, 278, 50080–50090. [Google Scholar] [CrossRef] [PubMed]
- Shingai, T. Ionic mechanism of water receptors in the laryngeal mucosa of the rabbit. Jpn. J. Physiol. 1977, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Shingai, T. Water fibers in the superior laryngeal nerve of the rat. Jpn. J. Physiol. 1980, 30, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [PubMed]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Zakir, H.M.; Mostafeezur, R.M.; Suzuki, A.; Hitomi, S.; Suzuki, I.; Maeda, T.; Seo, K.; Yamada, Y.; Yamamura, K.; Lev, S.; et al. Expression of TRPV1 channels after nerve injury provides an essential delivery tool for neuropathic pain attenuation. PLoS ONE 2012, 7, e44023. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.Z.; Unno, S.; Ando, H.; Masuda, Y.; Kitagawa, J. Neuron-Glia Crosstalk and Neuropathic Pain: Involvement in the Modulation of Motor Activity in the Orofacial Region. Int. J. Mol. Sci. 2017, 18, 205. [Google Scholar] [CrossRef] [PubMed]
- Bakri, M.M.; Yahya, F.; Munawar, K.M.M.; Kitagawa, J.; Hossain, M.Z. Transient receptor potential vanilloid 4 (TRPV4) expression on the nerve fibers of human dental pulp is upregulated under inflammatory condition. Arch. Oral Biol. 2018, 89, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef]
- Sant’Ambrogio, G. Nervous receptors of the tracheobronchial tree. Annu. Rev. Physiol. 1987, 49, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, S.; Kohzuki, M.; Sumi, Y.; Ebihara, T. Sensory stimulation to improve swallowing reflex and prevent aspiration pneumonia in elderly dysphagic people. J. Pharmacol. Sci. 2011, 115, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Vulchanova, L.; Wang, J.; Li, X.; Elde, R. Immunocytochemical localization of the vanilloid receptor 1 (VR1): Relationship to neuropeptides, the P2X3 purinoceptor and IB4 binding sites. Eur. J. Neurosci. 1999, 11, 946–958. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.H.; McDougall, S.J.; Fawley, J.A.; Smith, S.M.; Andresen, M.C. Primary afferent activation of thermosensitive TRPV1 triggers asynchronous glutamate release at central neurons. Neuron 2010, 65, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.H.; McDougall, S.J.; Fawley, J.A.; Andresen, M.C. TRPV1 marks synaptic segregation of multiple convergent afferents at the rat medial solitary tract nucleus. PLoS ONE 2011, 6, e25015. [Google Scholar] [CrossRef] [PubMed]
- Shoudai, K.; Peters, J.H.; McDougall, S.J.; Fawley, J.A.; Andresen, M.C. Thermally active TRPV1 tonically drives central spontaneous glutamate release. J. Neurosci. 2010, 30, 14470–14475. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.W.; Bailey, T.W.; Jin, Y.H.; Andresen, M.C. Vanilloid receptors presynaptically modulate cranial visceral afferent synaptic transmission in nucleus tractus solitarius. J. Neurosci. 2002, 22, 8222–8229. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, M.E.; Andresen, M.C. Vanilloids selectively sensitize thermal glutamate release from TRPV1 expressing solitary tract afferents. Neuropharmacology 2016, 101, 401–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.H.; Bailey, T.W.; Li, B.Y.; Schild, J.H.; Andresen, M.C. Purinergic and vanilloid receptor activation releases glutamate from separate cranial afferent terminals in nucleus tractus solitarius. J. Neurosci. 2004, 24, 4709–4717. [Google Scholar] [CrossRef] [PubMed]
- Bieger, D. Central nervous system control mechanisms of swallowing: A neuropharmacological perspective. Dysphagia 1993, 8, 308–310. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Tamir, R.; Qu, Y.; Klionsky, L.; Zhang, T.J.; Immke, D.; Wang, J.; Zhu, D.; Vanderah, T.W.; Porreca, F.; et al. AMG 9810 [(E)-3-(4-t-butylphenyl)-N-(2,3-dihydrobenzo[b][1,4] dioxin-6-yl)acrylamide], a novel vanilloid receptor 1 (TRPV1) antagonist with antihyperalgesic properties. J. Pharmacol. Exp. Ther. 2005, 313, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, M.H.; Price, T.J.; Entrena, J.M.; Cervero, F. Spinal NKCC1 blockade inhibits TRPV1-dependent referred allodynia. Mol. Pain 2007, 3, 17. [Google Scholar] [CrossRef] [PubMed]
- Hummel, M.; Knappenberger, T.; Reilly, M.; Whiteside, G.T. Pharmacological evaluation of NSAID-induced gastropathy as a “Translatable” model of referred visceral hypersensitivity. World J. Gastroenterol. 2017, 23, 6065–6076. [Google Scholar] [CrossRef] [PubMed]
- Lashinger, E.S.; Steiginga, M.S.; Hieble, J.P.; Leon, L.A.; Gardner, S.D.; Nagilla, R.; Davenport, E.A.; Hoffman, B.E.; Laping, N.J.; Su, X. AMTB, a TRPM8 channel blocker: Evidence in rats for activity in overactive bladder and painful bladder syndrome. Am. J. Physiol. 2008, 295, F803–F810. [Google Scholar] [CrossRef] [PubMed]
- Salat, K.; Filipek, B. Antinociceptive activity of transient receptor potential channel TRPV1, TRPA1, and TRPM8 antagonists in neurogenic and neuropathic pain models in mice. J. Zhejiang Univ. Sci. 2015, 16, 167–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez de Vega, M.J.; Gomez-Monterrey, I.; Ferrer-Montiel, A.; Gonzalez-Muniz, R. Transient Receptor Potential Melastatin 8 Channel (TRPM8) Modulation: Cool Entryway for Treating Pain and Cancer. J. Med. Chem. 2016, 59, 10006–10029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinuela-Fernandez, I.; Sun, L.; Jerina, H.; Curtis, J.; Allchorne, A.; Gooding, H.; Rosie, R.; Holland, P.; Tas, B.; Mitchell, R.; et al. The TRPM8 channel forms a complex with the 5-HT(1B) receptor and phospholipase D that amplifies its reversal of pain hypersensitivity. Neuropharmacology 2014, 79, 136–151. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.Z.; Ando, H.; Unno, S.; Masuda, Y.; Kitagawa, J. Activation of TRPV1 and TRPM8 Channels in the Larynx and Associated Laryngopharyngeal Regions Facilitates the Swallowing Reflex. Int. J. Mol. Sci. 2018, 19, 4113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124113
Hossain MZ, Ando H, Unno S, Masuda Y, Kitagawa J. Activation of TRPV1 and TRPM8 Channels in the Larynx and Associated Laryngopharyngeal Regions Facilitates the Swallowing Reflex. International Journal of Molecular Sciences. 2018; 19(12):4113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124113
Chicago/Turabian StyleHossain, Mohammad Zakir, Hiroshi Ando, Shumpei Unno, Yuji Masuda, and Junichi Kitagawa. 2018. "Activation of TRPV1 and TRPM8 Channels in the Larynx and Associated Laryngopharyngeal Regions Facilitates the Swallowing Reflex" International Journal of Molecular Sciences 19, no. 12: 4113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124113