Expression of Hypoxia-Inducible Factor 1α (HIF-1α) and Genes of Related Pathways in Altered Gravity


Abstract
:1. Introduction
2. Results
2.1. Altered Gravity Regulates Hypoxia-Inducible Factor (HIF)-1α-Dependent Gene Expression in Human Jurkat T Cells and Myelomonocytic U937 Cells and HIF-1α Protein Content in MDA-MB468 cells
2.2. Analysis of Hypoxia-Inducible Factor 1α Gene Expression
2.3. Analysis of Hypoxia-Inducible Factor-Related Genes
2.4. Rapid Alterations of HIF-Related Genes after 20 s of Altered Gravity in Human Jurkat T cells
2.5. Strong Response of HIF-Related Genes in Human Jurkat T Cells After 75 s of Hypergravity but Not After 300 s of Microgravity
2.6. HIF-Related Genes Were Primarily Differentially Regulated in Hypergravity in Human Jurkat T Cells
2.7. Dynamic Reaction and Adaptation Processes of Initially Differentially Altered HIF-Related Genes in Altered Gravity in Human Jurkat T Cells
2.8. Rapid Alterations of HIF-Related Genes After 20 s of Altered Gravity in Human U937 Cells
2.9. Strong Response of HIF-Related Genes in Human U937 Cells after 75 s of Hypergravity and After 300 s of Microgravity
2.10. HIF-Related Probe Sets Were Primarily Differentially Regulated in Hypergravity in Human U937 T Cells
2.11. Dynamic Reaction and Adaptation Processes of Initially Differentially Altered HIF-Related Genes in Altered Gravity in Human U937 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Parabolic Flight Experiment Platform
4.3. Preparation and Execution of the Parabolic Flight Experiments
4.4. RNA Isolation after the Parabolic Flight
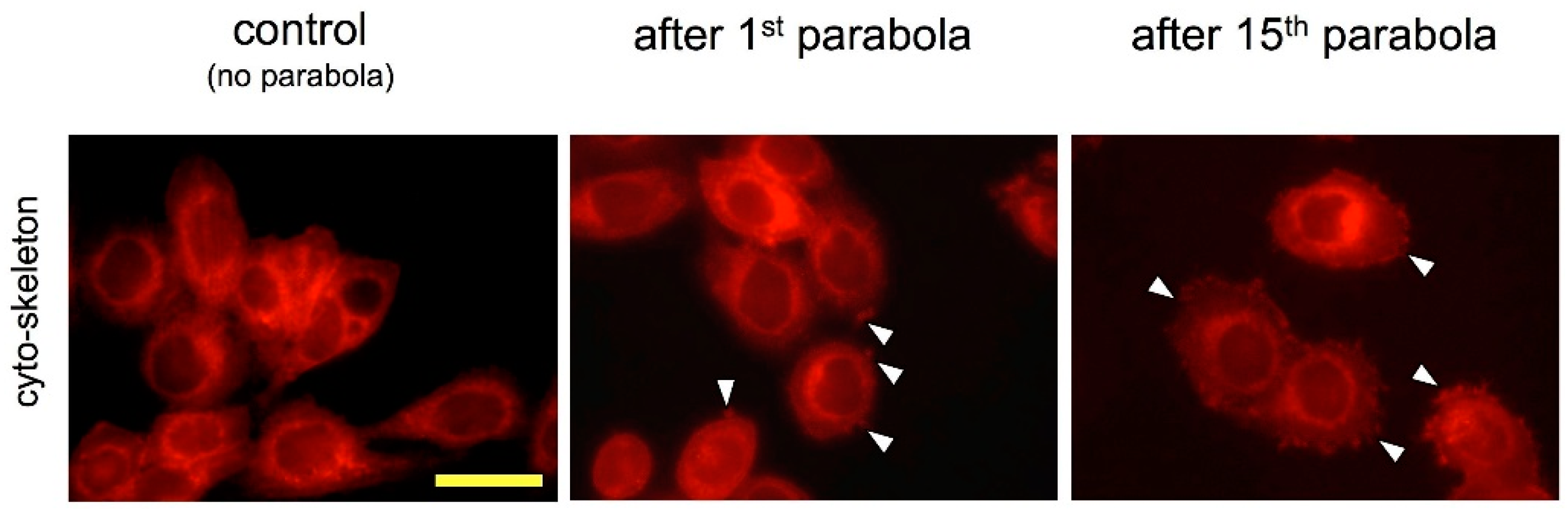
4.5. Western Blotting and Cytoskeleton Visualisation of MDA-MB-468 Cells
4.6. Technologische Experimente unter Schwerelosigkeit (TEXUS)-49 Suborbital Ballistic Rocket Experiment
4.7. TEXUS-51 Suborbital Ballistic Rocket Experiment
4.8. Procedures for the TEXUS Mission
4.9. Experimental Preparation and Integration for TEXUS Experiments
4.10. RNA Isolation After TEXUS Landing
4.11. RNA Sample Processing and Microarray Data Analysis for U937 Cells
4.12. RNA Sample Processing and Microarray Data Analysis for Jurkat T Cells
4.13. Selection of Array Probe sets for Analysis of HIF-1 Signaling Pathway Related Genes
4.14. Intra-Platform and Inter-Platform Comparisons
4.15. Data Availability
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DLR | German Aerospace Center |
| ESRANGE | European Space and Sounding Rocket Range |
| FC | fold change |
| HIF | hypoxia inducible factor |
| RIN | RNA Integrity Number |
| TEXUS | Technologische Experimente unter Schwerelosigkeit |
References
- Clément, G. Fundamentals of Space Medicine. Space Technology Library, 2nd ed.; Springer: Berlin, Germany, 2011; pp. 273–279. [Google Scholar]
- Choukèr, A.U.O. The Immune System in Space: Are we Prepared? Springer International Publishing: New York, NY, USA, 2016. [Google Scholar]
- Frippiat, J.P.; Crucian, B.E.; de Quervain, D.J.; Grimm, D.; Montano, N.; Praun, S.; Roozendaal, B.; Schelling, G.; Thiel, M.; Ullrich, O.; et al. Towards human exploration of space: The THESEUS review series on immunology research priorities. NPJ Microgr. 2016, 2, 16040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NASA. Office of Inspector General Report no. IG-16-003 “NASA’s Efforts to Manage Health and Human Performance Risks for Space Exploration”; NASA: Washington, DC, USA, 2015.
- The National Academies of Science. Engineering and Medicine. Recapturing a Future for Space Exploration: Life and Physical Sciences Research for a New Era; National Academic Press: Washington, DC, USA, 2001; Volume 6, pp. 99–203. [Google Scholar]
- McNamee, E.N.; Korns Johnson, D.; Homann, D.; Clambey, E.T. Hypoxia and hypoxia-inducible factors as regulators of T cell development, differentiation, and function. Immunol. Res. 2013, 55, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K.; Mauch, P.; Vergilio, J.A.; Sackstein, R.; Down, J.D. Distribution of hematopoietic stem cells in the bone marrow according to regional hypoxia. Proc. Natl. Acad. Sci. USA 2007, 104, 5431–5436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldwell, C.C.; Kojima, H.; Lukashev, D.; Armstrong, J.; Farber, M.; Apasov, S.G.; Sitkovsky, M.V. Differential effects of physiologically relevant hypoxic conditions on T lymphocyte development and effector functions. J. Immunol. 2001, 167, 6140–6149. [Google Scholar] [CrossRef] [PubMed]
- Colgan, S.P.; Taylor, C.T. Hypoxia: An alarm signal during intestinal inflammation. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Thiel, M.; Caldwell, C.C.; Kreth, S.; Kuboki, S.; Chen, P.; Smith, P.; Ohta, A.; Lentsch, A.B.; Lukashev, D.; Sitkovsky, M.V. Targeted deletion of HIF-1alpha gene in T cells prevents their inhibition in hypoxic inflamed tissues and improves septic mice survival. PLoS ONE 2007, 2, e853. [Google Scholar] [CrossRef] [PubMed]
- Karhausen, J.; Furuta, G.T.; Tomaszewski, J.E.; Johnson, R.S.; Colgan, S.P.; Haase, V.H. Epithelial hypoxia-inducible factor-1 is protective in murine experimental colitis. J. Clin. Investig. 2004, 114, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Higashiyama, M.; Hokari, R.; Hozumi, H.; Kurihara, C.; Ueda, T.; Watanabe, C.; Tomita, K.; Nakamura, M.; Komoto, S.; Okada, Y.; et al. HIF-1 in T cells ameliorated dextran sodium sulfate-induced murine colitis. J. Leukoc. Biol. 2012, 91, 901–909. [Google Scholar] [CrossRef]
- Gaber, T.; Haupl, T.; Sandig, G.; Tykwinska, K.; Fangradt, M.; Tschirschmann, M.; Hahne, M.; Dziurla, R.; Erekul, K.; Lautenbach, M.; et al. Adaptation of human CD4+ T cells to pathophysiological hypoxia: A transcriptome analysis. J. Rheumatol. 2009, 36, 2655–2669. [Google Scholar] [CrossRef]
- Jewell, U.R.; Kvietikova, I.; Scheid, A.; Bauer, C.; Wenger, R.H.; Gassmann, M. Induction of HIF-1alpha in response to hypoxia is instantaneous. FASEB J. 2001, 15, 1312–1314. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-inducible factors: Mediators of cancer progression and targets for cancer therapy. Trends Pharmacol. Sci. 2012, 33, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.H.; Barbi, J.; Pan, F. Hypoxia-inducible factors in T lymphocyte differentiation and function. A Review in the Theme: Cellular Responses to Hypoxia. Am. J. Physiol. Cell Physiol. 2015, 309, C580–C589. [Google Scholar] [CrossRef] [PubMed]
- Blouin, C.C.; Page, E.L.; Soucy, G.M.; Richard, D.E. Hypoxic gene activation by lipopolysaccharide in macrophages: Implication of hypoxia-inducible factor 1alpha. Blood 2004, 103, 1124–1130. [Google Scholar] [CrossRef]
- Hong, W.X.; Hu, M.S.; Esquivel, M.; Liang, G.Y.; Rennert, R.C.; McArdle, A.; Paik, K.J.; Duscher, D.; Gurtner, G.C.; Lorenz, H.P.; et al. The Role of Hypoxia-Inducible Factor in Wound Healing. Adv. Wound Care 2014, 3, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imtiyaz, H.Z.; Simon, M.C. Hypoxia-inducible factors as essential regulators of inflammation. Curr. Top. Microbiol. Immunol. 2010, 345, 105–120. [Google Scholar]
- Antoniou, X.; Gassmann, M.; Ogunshola, O.O. Cdk5 interacts with Hif-1alpha in neurons: A new hypoxic signalling mechanism? Brain Res. 2011, 1381, 1–10. [Google Scholar] [CrossRef]
- Janmaleki, M.; Pachenari, M.; Seyedpour, S.M.; Shahghadami, R.; Sanati-Nezhad, A. Impact of Simulated Microgravity on Cytoskeleton and Viscoelastic Properties of Endothelial Cell. Sci. Rep. 2016, 6, 32418. [Google Scholar] [CrossRef] [Green Version]
- Stohr, N.; Lederer, M.; Reinke, C.; Meyer, S.; Hatzfeld, M.; Singer, R.H.; Huttelmaier, S. ZBP1 regulates mRNA stability during cellular stress. J. Cell Biol. 2006, 175, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Prasanth, K.V.; Prasanth, S.G.; Xuan, Z.; Hearn, S.; Freier, S.M.; Bennett, C.F.; Zhang, M.Q.; Spector, D.L. Regulating gene expression through RNA nuclear retention. Cell 2005, 123, 249–263. [Google Scholar] [CrossRef]
- Moeller, B.J.; Cao, Y.; Li, C.Y.; Dewhirst, M.W. Radiation activates HIF-1 to regulate vascular radiosensitivity in tumors: Role of reoxygenation, free radicals, and stress granules. Cancer Cell 2004, 5, 429–441. [Google Scholar] [CrossRef]
- Lukashev, D.; Klebanov, B.; Kojima, H.; Grinberg, A.; Ohta, A.; Berenfeld, L.; Wenger, R.H.; Ohta, A.; Sitkovsky, M. Cutting edge: Hypoxia-inducible factor 1alpha and its activation-inducible short isoform I.1 negatively regulate functions of CD4+ and CD8+ T lymphocytes. J. Immunol. 2006, 177, 4962–4965. [Google Scholar] [CrossRef] [PubMed]
- Dang, E.V.; Barbi, J.; Yang, H.Y.; Jinasena, D.; Yu, H.; Zheng, Y.; Bordman, Z.; Fu, J.; Kim, Y.; Yen, H.R.; et al. Control of T(H)17/T(reg) balance by hypoxia-inducible factor 1. Cell 2011, 146, 772–784. [Google Scholar] [CrossRef] [PubMed]
- Waickman, A.T.; Powell, J.D. mTOR, metabolism, and the regulation of T-cell differentiation and function. Immunol. Rev. 2012, 249, 43–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalek, R.D.; Gerriets, V.A.; Jacobs, S.R.; Macintyre, A.N.; MacIver, N.J.; Mason, E.F.; Sullivan, S.A.; Nichols, A.G.; Rathmell, J.C. Cutting edge: Distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets. J. Immunol. 2011, 186, 3299–3303. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Regulation of metabolism by hypoxia-inducible factor 1. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.Z.; Wang, R.; Huang, G.; Vogel, P.; Neale, G.; Green, D.R.; Chi, H. HIF1alpha-dependent glycolytic pathway orchestrates a metabolic checkpoint for the differentiation of TH17 and Treg cells. J. Exp. Med. 2011, 208, 1367–1376. [Google Scholar] [CrossRef]
- Kryczek, I.; Zhao, E.; Liu, Y.; Wang, Y.; Vatan, L.; Szeliga, W.; Moyer, J.; Klimczak, A.; Lange, A.; Zou, W. Human TH17 cells are long-lived effector memory cells. Sci. Transl. Med. 2011, 3, 104ra100. [Google Scholar] [CrossRef]
- Riboldi, E.; Porta, C.; Morlacchi, S.; Viola, A.; Mantovani, A.; Sica, A. Hypoxia-mediated regulation of macrophage functions in pathophysiology. Int. Immunol. 2013, 25, 67–75. [Google Scholar] [CrossRef]
- Jurgensen, J.S.; Rosenberger, C.; Wiesener, M.S.; Warnecke, C.; Horstrup, J.H.; Grafe, M.; Philipp, S.; Griethe, W.; Maxwell, P.H.; Frei, U.; et al. Persistent induction of HIF-1alpha and -2alpha in cardiomyocytes and stromal cells of ischemic myocardium. FASEB J. 2004, 18, 1415–1417. [Google Scholar] [CrossRef]
- Azimi, I.; Petersen, R.M.; Thompson, E.W.; Roberts-Thomson, S.J.; Monteith, G.R. Hypoxia-induced reactive oxygen species mediate N-cadherin and SERPINE1 expression, EGFR signalling and motility in MDA-MB-468 breast cancer cells. Sci. Rep. 2017, 7, 15140. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, M.; Grenacher, B.; Rohde, B.; Vogel, J. Quantifying Western blots: Pitfalls of densitometry. Electrophoresis 2009, 30, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Hauschild, S.; Huge, A.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Dynamic gene expression response to altered gravity in human T cells. Sci. Rep. 2017, 7, 5204. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Tauber, S.; Christoffel, S.; Huge, A.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Rapid coupling between gravitational forces and the transcriptome in human myelomonocytic U937 cells. Sci. Rep. 2018, 8, 13267. [Google Scholar] [CrossRef] [PubMed]
- Tauber, S.; Christoffel, S.; Thiel, C.S.; Ullrich, O. Transcriptional Homeostasis of Oxidative Stress-Related Pathways in Altered Gravity. Int. J. Mol. Sci. 2018, 19, 2814. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Huge, A.; Hauschild, S.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Stability of gene expression in human T cells in different gravity environments is clustered in chromosomal region 11p15.4. NPJ Microgr. 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.P.; Niessing, D. Assembly of mRNA-protein complexes for directional mRNA transport in eukaryotes—An overview. Curr. Protein Pept. Sci. 2012, 13, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.; Heym, R.G.; Mayer, A.; Kramer, K.; Schmid, M.; Cramer, P.; Urlaub, H.; Jansen, R.P.; Niessing, D. A cytoplasmic complex mediates specific mRNA recognition and localization in yeast. PLoS Biol. 2011, 9, e1000611. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.E.; Bullock, S.L. Subcellular mRNA localization in animal cells and why it matters. Science 2009, 326, 1212–1216. [Google Scholar] [CrossRef]
- Rothman, J.E. Mechanisms of intracellular protein transport. Nature 1994, 372, 55–63. [Google Scholar] [CrossRef]
- Hoffman, L.; Jensen, C.C.; Yoshigi, M.; Beckerle, M. Mechanical signals activate p38 MAPK pathway-dependent reinforcement of actin via mechanosensitive HspB1. Mol. Biol. Cell 2017, 28, 2661–2675. [Google Scholar] [CrossRef] [Green Version]
- Uhler, C.; Shivashankar, G.V. Regulation of genome organization and gene expression by nuclear mechanotransduction. Nat. Rev. Mol. Cell Biol. 2017, 18, 717–727. [Google Scholar] [CrossRef]
- Abercromby, A.F.J.; Conkin, J.; Gernhardt, M.L. Modeling Oxygen Prebreathe Protocols for Exploration Extravehicular Activities Using Variable Pressure Suits. In Proceedings of the 88th Annual Scientific Meeting of the Aerospace Medical Association, Denver, CO, USA, 30 April–4 May 2017. [Google Scholar]
- Patrick, N.; Locke, J.; Trevino, L.; Trevino, R. Extravehicular Activity Operations and Advancements. The Space Shuttle and its Operations. Available online: https://www.nasa.gov/centers/johnson/pdf/584725main_Wings-ch3d-pgs110-129.pdf (accessed on 18 January 2019).
- Gotoh, T.M.; Fujiki, N.; Tanaka, K.; Matsuda, T.; Gao, S.; Morita, H. Acute hemodynamic responses in the head during microgravity induced by free drop in anesthetized rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R1063–R1068. [Google Scholar] [CrossRef] [PubMed]
- Imray, C.; Wright, A.; Subudhi, A.; Roach, R. Acute mountain sickness: Pathophysiology, prevention, and treatment. Prog. Cardiovasc. Dis. 2010, 52, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.F.; Hargens, A.R. Spaceflight-Induced Intracranial Hypertension and Visual Impairment: Pathophysiology and Countermeasures. Physiol. Rev. 2018, 98, 59–87. [Google Scholar] [CrossRef] [PubMed]
- Simonson, T.S. Altitude Adaptation: A Glimpse Through Various Lenses. High Alt. Med. Biol. 2015, 16, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shweiki, D.; Itin, A.; Soffer, D.; Keshet, E. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature 1992, 359, 843–845. [Google Scholar] [CrossRef]
- Zachary, I. VEGF signalling: Integration and multi-tasking in endothelial cell biology. Biochem. Soc. Trans.s 2003, 31, 1171–1177. [Google Scholar] [CrossRef]
- Connolly, D.T.; Heuvelman, D.M.; Nelson, R.; Olander, J.V.; Eppley, B.L.; Delfino, J.J.; Siegel, N.R.; Leimgruber, R.M.; Feder, J. Tumor vascular permeability factor stimulates endothelial cell growth and angiogenesis. J. Clin. Investig. 1989, 84, 1470–1478. [Google Scholar] [CrossRef]
- Frede, S.; Stockmann, C.; Freitag, P.; Fandrey, J. Bacterial lipopolysaccharide induces HIF-1 activation in human monocytes via p44/42 MAPK and NF-κB. Biochem. J. 2006, 396, 517–527. [Google Scholar] [CrossRef]
- Stockmann, C.; Schadendorf, D.; Klose, R.; Helfrich, I. The impact of the immune system on tumor: Angiogenesis and vascular remodelling. Front. Oncol. 2014, 4, 69. [Google Scholar] [CrossRef] [PubMed]
- Meehan, R.T. Immune suppression at high altitude. Ann. Emerg. Med. 1987, 16, 974–979. [Google Scholar] [CrossRef]
- Klokker, M.; Kharazmi, A.; Galbo, H.; Bygbjerg, I.; Pedersen, B.K. Influence of in vivo hypobaric hypoxia on function of lymphocytes, neutrocytes, natural killer cells, and cytokines. J. Appl. Physiol. 1993, 74, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Sams, C. Immune system dysregulation during spaceflight: Clinical risk for exploration-class missions. J. Leukoc. Biol. 2009, 86, 1017–1018. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.B.; Liu, X.C.; Qin, X.Y.; Chen, J.; Ren, P.G.; Deng, W.F.; Zhang, J. Effect of High-Fat Diet on Immature Female Mice and Messenger and Noncoding RNA Expression Profiling in Ovary and White Adipose Tissue. Reprod. Sci. 2018, 1933719118765966. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Jung, H.J.; Choi, H.J.; Cho, J.I.; Park, H.J.; Kwon, T.H. miR-34c-5p and CaMKII are involved in aldosterone-induced fibrosis in kidney collecting duct cells. American journal of physiology. Ren. Physiol. 2018, 314, F329–F342. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhao, Y.; Wu, G.; Lin, B.; Li, Z.; Liu, X. Differential miRNAomics of the synovial membrane in knee osteoarthritis induced by bilateral anterior cruciate ligament transection in rats. Mol. Med. Rep. 2018, 18, 4051–4057. [Google Scholar] [CrossRef]
- Datar, J.; Regassa, A.; Kim, W.K.; Taylor, C.G.; Zahradka, P.; Suh, M. Lipid Metabolism is Closely Associated with Normal Testicular Growth Based on Global Transcriptome Profiles in Normal and Underdeveloped Testis of Obese Zucker (fa/fa) Rats. Lipids 2017, 52, 951–960. [Google Scholar] [CrossRef]
- Hsiao, A.; Worrall, D.S.; Olefsky, J.M.; Subramaniam, S. Variance-modeled posterior inference of microarray data: Detecting gene-expression changes in 3T3-L1 adipocytes. Bioinformatics 2004, 20, 3108–3127. [Google Scholar] [CrossRef]
- Miller, R.A.; Galecki, A.; Shmookler-Reis, R.J. Interpretation, design, and analysis of gene array expression experiments. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2001, 56, B52–B57. [Google Scholar] [CrossRef]
- Budhraja, V.; Spitznagel, E.; Schaiff, W.T.; Sadovsky, Y. Incorporation of gene-specific variability improves expression analysis using high-density DNA microarrays. BMC Biol. 2003, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Allison, D.B.; Cui, X.; Page, G.P.; Sabripour, M. Microarray data analysis: From disarray to consolidation and consensus. Nature reviews. Genetics 2006, 7, 55–65. [Google Scholar] [PubMed]
- Callow, M.J.; Dudoit, S.; Gong, E.L.; Speed, T.P.; Rubin, E.M. Microarray expression profiling identifies genes with altered expression in HDL-deficient mice. Genome Res. 2000, 10, 2022–2029. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.S.; Korotchkina, L.G. Regulation of mammalian pyruvate dehydrogenase complex by phosphorylation: Complexity of multiple phosphorylation sites and kinases. Exp. Mol. Med. 2001, 33, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Tchernyshyov, I.; Semenza, G.L.; Dang, C.V. HIF-1-mediated expression of pyruvate dehydrogenase kinase: A metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006, 3, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papandreou, I.; Cairns, R.A.; Fontana, L.; Lim, A.L.; Denko, N.C. HIF-1 mediates adaptation to hypoxia by actively downregulating mitochondrial oxygen consumption. Cell Metab. 2006, 3, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Koukourakis, M.I.; Giatromanolaki, A.; Sivridis, E.; Gatter, K.C.; Harris, A.L. Pyruvate dehydrogenase and pyruvate dehydrogenase kinase expression in non small cell lung cancer and tumor-associated stroma. Neoplasia 2005, 7, 1–6. [Google Scholar] [CrossRef]
- Fu, Y.; Maianu, L.; Melbert, B.R.; Garvey, W.T. Facilitative glucose transporter gene expression in human lymphocytes, monocytes, and macrophages: A role for GLUT isoforms 1, 3, and 5 in the immune response and foam cell formation. Blood Cells Mol. Dis. 2004, 1, 182–190. [Google Scholar] [CrossRef]
- Kubala, M.H.; Punj, V.; Placencio-Hickok, V.R.; Fang, H.; Fernandez, G.E.; Sposto, R.; DeClerck, Y.A. Plasminogen Activator Inhibitor-1 Promotes the Recruitment and Polarization of Macrophages in Cancer. Cell Rep. 2018, 25, 2177–2191. [Google Scholar] [CrossRef]
- Corcoran, S.E.; O’Neill, L.A. HIF1α and metabolic reprogramming in inflammation. J. Clin. Investig. 2016, 126, 3699–3707. [Google Scholar] [CrossRef] [Green Version]
- Selak, M.A.; Armour, S.M.; MacKenzie, E.D.; Boulahbel, H.; Watson, D.G.; Mansfield, K.D.; Pan, Y.; Simon, M.C.; Thompson, C.B.; Gottlieb, E. Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-alpha prolyl hydroxylase. Cancer Cell 2005, 7, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Kuschel, A.; Simon, P.; Tug, S. Functional regulation of HIF-1α under normoxia—Is there more than post-translational regulation? J. Cell Physiol. 2012, 227, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.T.; Cummins, E.P. The role of NF-κB in hypoxia-induced gene expression. Ann. N. Y. Acad. Sci. 2009, 1177, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Rius, J.; Guma, M.; Schachtrup, C.; Akassoglou, K.; Zinkernagel, A.S.; Nizet, V.; Johnson, R.S.; Haddad, G.G.; Karin, M. NF-κB links innate immunity to the hypoxic response through transcriptional regulation of HIF-1alpha. Nature 2008, 453, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, K.; Thiel, C.; Timm, J.; Schmidt, P.M.; Huber, K.; Tauber, S.; Hemmersbach, R.; Seibt, D.; Kroll, H.; Grote, K.-H.; et al. Microgravity-induced alterations in signal transduction in cells of the immune system. Acta Astronaut. 2010, 67, 1116–1125. [Google Scholar] [CrossRef]
- Chang, T.T.; Walther, I.; Li, C.F.; Boonyaratanakornkit, J.; Galleri, G.; Meloni, M.A.; Pippia, P.; Cogoli, A.; Hughes-Fulford, M. The Rel/NF-κB pathway and transcription of immediate early genes in T cell activation are inhibited by microgravity. J. Leukoc. Biol. 2012, 92, 1133–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewell, U.R.; Gassmann, M. Mammalian gene expression in hypoxic conditions. Zoology 2001, 104, 192–197. [Google Scholar] [CrossRef]
- Wang, T.; Chen, H.; Lv, K.; Ji, G.; Liang, F.; Zhang, Y.; Wang, Y.; Liu, X.; Cao, H.; Kan, G.; Xiong, J.; Li, Y.; Qu, L. Activation of HIF-1α and its downstream targets in rat hippocampus after long-term simulated microgravity exposure. Biochem. Biophys. Res. Commun. 2017, 485, 591–597. [Google Scholar] [CrossRef]
- Zhang, Y.; Moreno-Villanueva, M.; Krieger, S.; Ramesh, G.T.; Neelam, S.; Wu, H. Transcriptomics, NF-κB Pathway, and Their Potential Spaceflight-Related Health Consequences. Int. J. Mol. Sci. 2017, 18, 1166. [Google Scholar] [CrossRef]
- Thiel, C.S.; Paulsen, K.; Bradacs, G.; Lust, K.; Tauber, S.; Dumrese, C.; Hilliger, A.; Schoppmann, K.; Biskup, J.; Golz, N.; et al. Rapid alterations of cell cycle control proteins in human T lymphocytes in microgravity. Cell Commun. Signal. CCS 2012, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, K.; Tauber, S.; Dumrese, C.; Bradacs, G.; Simmet, D.M.; Golz, N.; Hauschild, S.; Raig, C.; Engeli, S.; Gutewort, A.; et al. Regulation of ICAM-1 in cells of the monocyte/macrophage system in microgravity. BioMed Res. Int. 2015, 2015, 538786. [Google Scholar] [CrossRef] [PubMed]
- Welinder, C.; Ekblad, L. Coomassie staining as loading control in Western blot analysis. J. Proteome Res. 2011, 10, 1416–1419. [Google Scholar] [CrossRef] [PubMed]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment Group Description | Gravity Condition | 19th DLR PFC 23rd DLR PFC | TEXUS-49 TEXUS-51 |
|---|---|---|---|
| Cell culture control | 1 g | n/a | CC (TEXUS-51) |
| Hardware 1 g ground control | 1 g | H/W 1 g GC | H/W 1 g GC |
| 1 g in-flight | 1 g | 1 g IF | 1 g IF (TEXUS-51) |
| Baseline (BL)/Hypergravity [directly before µg phase] | hyp-g | BL-PFC hyp-g [1.8 g, 20 s] | BL-TX hyp-g [max. 13.5 g, 75 s] |
| Microgravity (µg) | µg | µg [20 s] | µg [300 s] |
| Primary Comparisons | ||||
|---|---|---|---|---|
| Gene Expression | 1 g IF vs. H/W 1 g GC | BL-PFC hyp-g vs. 1 g IF | µg vs. BL-PFC hyp-g | BL-PFC hyp-g vs. H/W 1 g GC |
| up-regulated | 85 | 19 | 3 | 51 |
| down-regulated | 29 | 19 | 11 | 19 |
| total | 114 | 38 | 14 | 70 |
| Gene Expression | Hypergravity-Sensitive Probe Sets | Microgravity-Sensitive Probe Sets | ||||
|---|---|---|---|---|---|---|
| Significantly differentially expressed in the primary comparison BL-PFC hyp-g vs. H/W 1 g GC | Significantly differentially expressed in the primary comparison µg vs. BL-PFC hyp-g NOT significantly expressed in the same direction in the primary comparisons µg vs. BL-PFC hyp-g and BL-PFC hyp-g vs. H/W 1 g GC | |||||
| Number | average FC of BL-PFC hyp-g vs. 1 g IF | min/max FC of BL-PFC hyp-g vs. H/W 1 g GC | Number | average FC µg vs. BL-PFC hyp-g | min/max µg vs. BL-PFC hyp-g | |
| up-regulated | 51 | 1.15 | 1.39 | 3 | 1.08 | 1.11 |
| down-regulated | 19 | −1.17 | −1.62 | 7 | −1.04 | −1.07 |
| total | 70 | - | - | 10 | - | - |
| Primary Comparisons | ||||||
|---|---|---|---|---|---|---|
| Gene Expression | 1 g IF vs. H/W 1 g GC | BL-TX hyp-g vs. 1 g IF | µg vs. BL-TX hyp-g | µg vs. 1 g IF | BL-TX hyp-g vs. H/W 1 g GC | H/W 1 g GC vs. CC |
| up-regulated | 29 | 37 | 5 | 33 | 41 | 125 |
| down-regulated | 5 | 114 | 4 | 109 | 82 | 52 |
| total | 34 | 151 | 9 | 142 | 123 | 177 |
| Gene Expression | Hypergravity-Sensitive Probe Sets | Microgravity-Sensitive Probe Sets | ||||
|---|---|---|---|---|---|---|
| Significantly differentially expressed in the primary comparison BL-TX hyp-g vs. H/W 1 g GC | Significantly differentially expressed in the primary comparison µg vs. BL-TX hyp-g NOT significantly expressed in the same direction in the primary comparisons µg vs. BL-TX hyp-g and BL-TX hyp-g vs. H/W 1 g GC | |||||
| Number | average fold change (FC) of BL-TX hyp-g vs. H/W 1 g GC | min/max FC of BL-TX hyp-g vs. H/W 1 g GC | Number | average FC µg vs. BL-TX hyp-g | min/max µg vs. BL-TX hyp-g | |
| up-regulated | 41 | 1.26 | 1.76 | 5 | 1.05 | 1.08 |
| down-regulated | 82 | −1.14 | −1.35 | 4 | −1.10 | −1.15 |
| total | 123 | - | - | 9 | - | - |
| Gene Name | Gene ID | Probe Set ID | Fold Change 23rd DLR PFC BL-PFC hyp-g vs. H/W 1 g GC | Fold Change TEXUS-51 BL-TX hyp-g vs. H/W 1 g GC |
|---|---|---|---|---|
| SLC2A1 | 6513 | TC01002578.hg.1 | −1.11 | −1.03 |
| PIK3R3; RP11-322N21.2 | 8503 | TC01002616.hg.1 | +1.05 | +1.3 |
| ARNT | 405 | TC01003212.hg.1 | +1.15 | +1.1 |
| PDK1 | 5163 | TC02001031.hg.1 | +1.07 | +1.17 |
| VHL | 7428 | TC03000055.hg.1 | +1.07 | +1.17 |
| PIK3CA | 5290 | TC03000951.hg.1 | +1.18 | +1.63 |
| CAMK2D | 817 | TC04001486.hg.1 | +1.11 | +1.06 |
| MAPK14 | 1432 | TC06000523.hg.1 | +1.08 | +1.14 |
| IFNGR1 | 3459 | TC06002152.hg.1 | 11.13 | +1.36 |
| NOS3; ATG9B | 4846 | TC07001009.hg.1 | −1.03 | −1.12 |
| RPS6 | 6194 | TC09000938.hg.1 | +1.13 | +1.32 |
| VIM | 7431 | TC10000126.hg.1 | +1.19 | +1.14 |
| CDKN1B | 1027 | TC12000178.hg.1 | +1.38 | +1.06 |
| FLT1 | 2321 | TC13000517.hg.1 | +1.12 | +1.32 |
| HIF1A | 3091 | TC14002197.hg.1 | +1.25 | +1.66 |
| MAP2K1 | 5604 | TC15000613.hg.1 | +1.3 | +1.07 |
| IGF1R | 3480 | TC15000949.hg.1 | +1.18 | +1.08 |
| PRKCB | 5579 | TC16000260.hg.1 | +1.06 | +1.12 |
| STAT3 | 6774 | TC17001531.hg.1 | +1.33 | +1.26 |
| AKT2 | 208 | TC19001532.hg.1 | −1.08 | −1.09 |
| PFKL | 5211 | TC21000222.hg.1 | −1.13 | −1.07 |
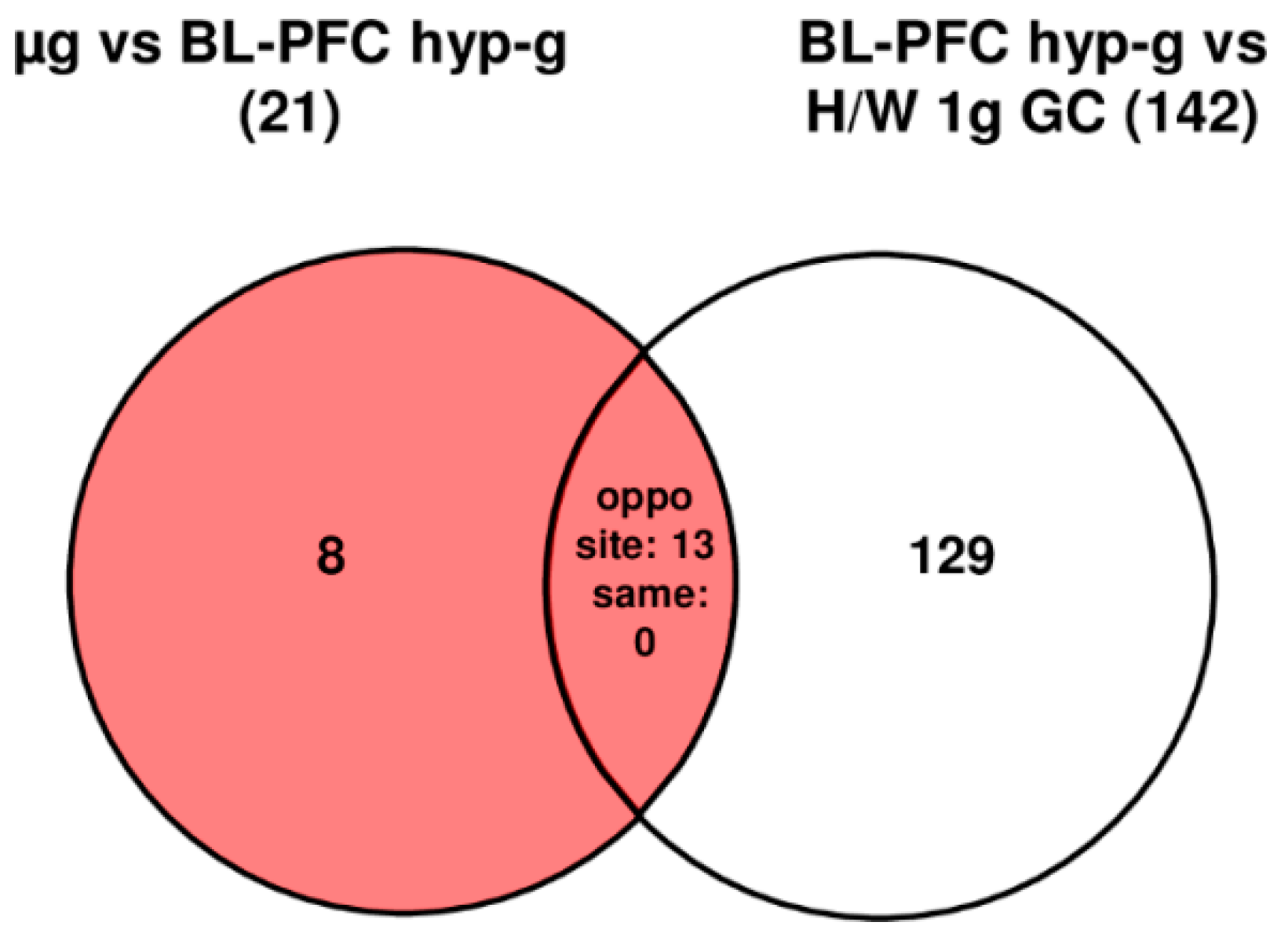
| Primary Comparisons | ||||
|---|---|---|---|---|
| Gene Expression | 1 g IF vs. H/W 1 g GC | BL-PFC hyp-g vs. 1 g IF | µg vs. BL-PFC hyp-g | BL-PFC hyp-g vs. H/W 1 g GC |
| up-regulated | 15 | 61 | 6 | 75 |
| down-regulated | 30 | 43 | 15 | 67 |
| total | 45 | 104 | 21 | 142 |
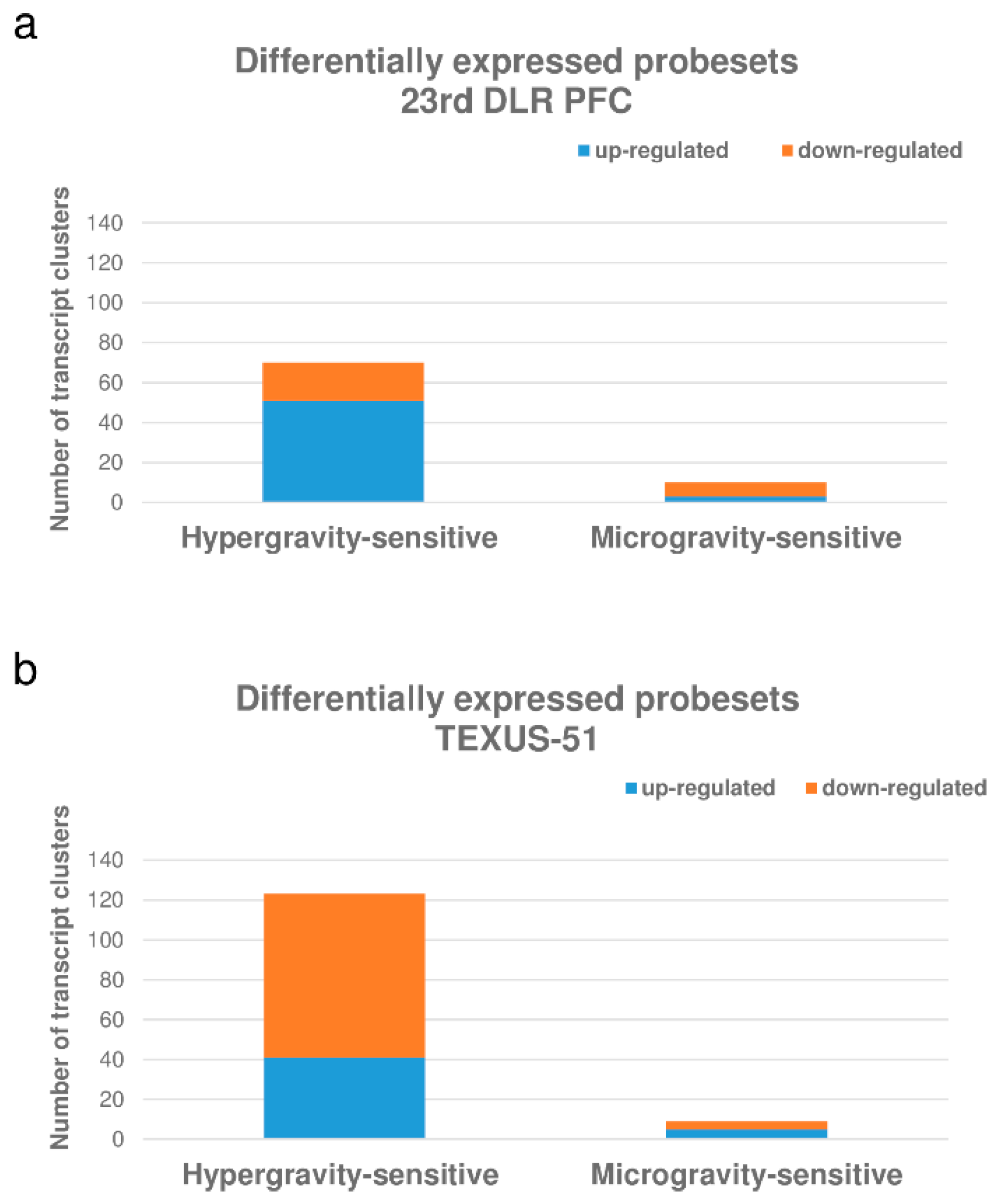
| Gene Expression | Hypergravity-Sensitive Probe Sets | Microgravity-Sensitive Probe Sets | ||||
|---|---|---|---|---|---|---|
| Significantly differentially expressed in the primary comparison BL-PFC hyp-g vs. H/W 1 g GC | Significantly differentially expressed in the primary comparison µg vs. BL-PFC hyp-g NOT significantly expressed in the same direction in the primary comparisons µg vs. BL-PFC hyp-g and BL-PFC hyp-g vs. H/W 1 g GC | |||||
| Number | average FC of BL-PFC hyp-g vs. H/W 1 g GC | min/max FC of BL- PFC hyp-g vs. H/W 1 g GC | Number | average FC µg vs. BL- PFC hyp-g | min/max µg vs. BL- PFC hyp-g | |
| up-regulated | 75 | 2.00 | 3.60 | 6 | 1.33 | 1.82 |
| down-regulated | 67 | −2.00 | −6.46 | 15 | −1.46 | −2.09 |
| total | 142 | - | - | 21 | - | - |
| Primary Comparisons | |||
|---|---|---|---|
| Gene Expression | µg vs. BL-TX hyp-g | BL-TX hyp-g vs. H/W 1 g GC | µg vs. H/W 1 g GC |
| up-regulated | 5 | 90 | 6 |
| down-regulated | 33 | 15 | 9 |
| total | 38 | 105 | 15 |
| Gene Expression | Hypergravity-Sensitive Probe Sets | Microgravity-Sensitive Probe Sets | ||||
|---|---|---|---|---|---|---|
| . | Significantly differentially expressed in the primary comparison BL-TX hyp-g vs. H/W 1 g GC | Significantly differentially expressed in the primary comparison µg vs. BL-TX hyp-g NOT significantly expressed in the same direction in the primary comparisons µg vs. BL-TX hyp-g and BL-TX hyp-g vs. H/W 1 g GC | ||||
| Number | average FC of BL-TX hyp-g vs. H/W 1 g GC | min/max FC of BL-TX hyp-g vs. H/W 1 g GC | Number | average FC µg vs. BL-TX hyp-g | min/max µg vs. BL-TX hyp-g | |
| up-regulated | 90 | 1.92 | 4.69 | 5 | 1.79 | 2.48 |
| down-regulated | 15 | −1.74 | −2.08 | 33 | −1.69 | −2.35 |
| total | 105 | - | - | 38 | - | - |
| Gene Name | Gene ID | Probe Set ID | Fold Change 19th DLR PFC BL-PFC hyp-g vs. H/W 1 g GC | Fold Change TEXUS-49 BL-TX hyp-g vs. H/W 1 g GC |
|---|---|---|---|---|
| SERPINE1 | 5054 | NM_000602 | +2.16 | + 1.50 |
| PDK1 | 5163 | BC039158 | +1.30 | +1.48 |
| PDK1 | 5163 | NM_002610 | +1.23 | +1.62 |
| SLC2A3 | 6515 | NM_006931 | + 1.81 | +1.50 |
| Gene Name | Gene ID | Probe Set ID | Fold Change 19th DLR PFC µg vs. BL-PFC hyp-g | Fold Change TEXUS-49 µg vs. BL-TX hyp-g |
|---|---|---|---|---|
| IL1B | 3553 | NM_000576 | −1.65 | −1.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogel, J.; Thiel, C.S.; Tauber, S.; Stockmann, C.; Gassmann, M.; Ullrich, O. Expression of Hypoxia-Inducible Factor 1α (HIF-1α) and Genes of Related Pathways in Altered Gravity. Int. J. Mol. Sci. 2019, 20, 436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020436
Vogel J, Thiel CS, Tauber S, Stockmann C, Gassmann M, Ullrich O. Expression of Hypoxia-Inducible Factor 1α (HIF-1α) and Genes of Related Pathways in Altered Gravity. International Journal of Molecular Sciences. 2019; 20(2):436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020436
Chicago/Turabian StyleVogel, Johannes, Cora Sandra Thiel, Svantje Tauber, Christian Stockmann, Max Gassmann, and Oliver Ullrich. 2019. "Expression of Hypoxia-Inducible Factor 1α (HIF-1α) and Genes of Related Pathways in Altered Gravity" International Journal of Molecular Sciences 20, no. 2: 436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020436