The Importance of Conserved Serine for C-Terminally Encoded Peptides Function Exertion in Apple

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
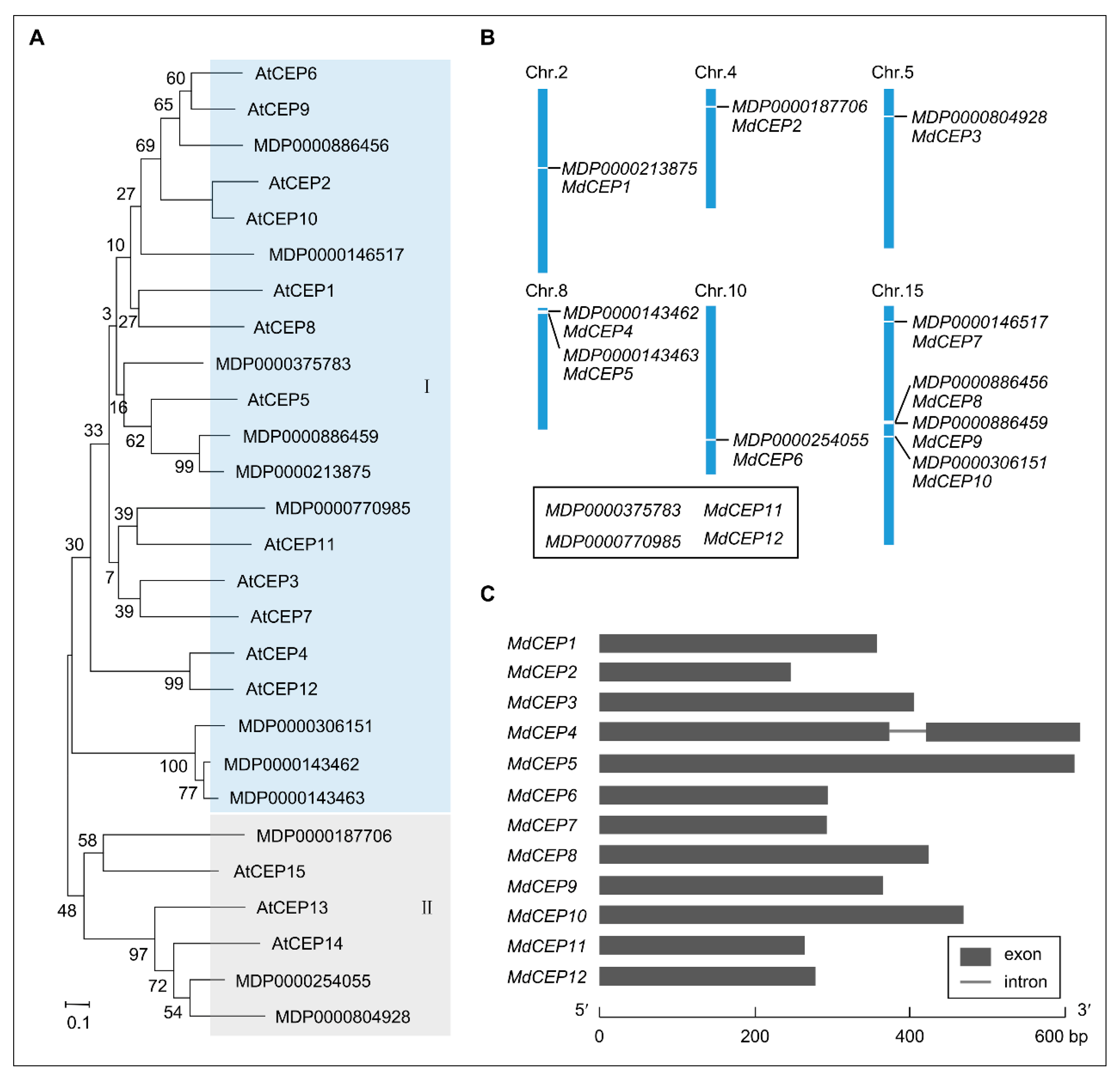
2.1. Identification and Phylogenetic Analysis of MdCEPs in Apple
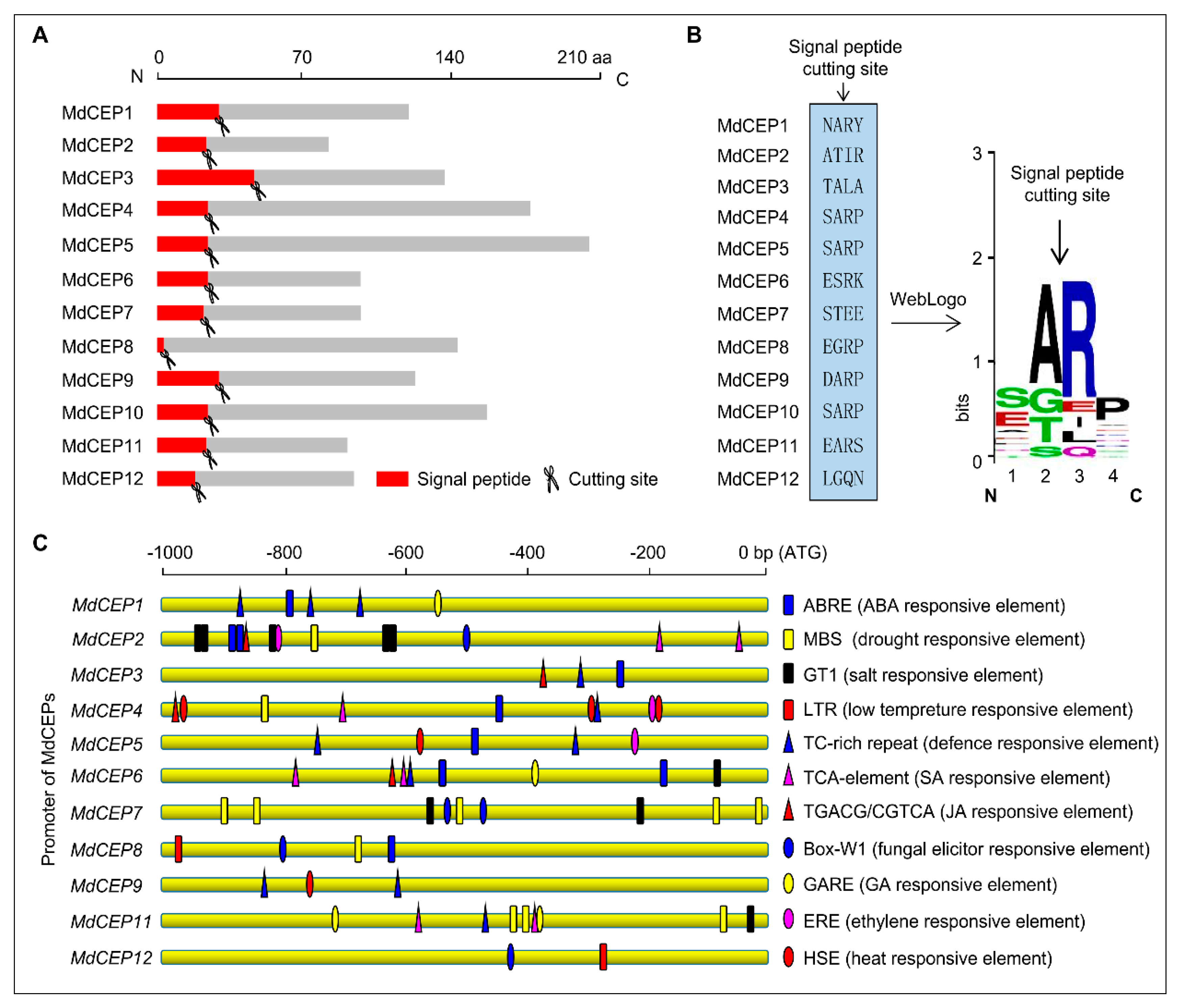
2.2. Signal Peptide Cleavage Sites Prediction and Promoter Analysis of MdCEP Members
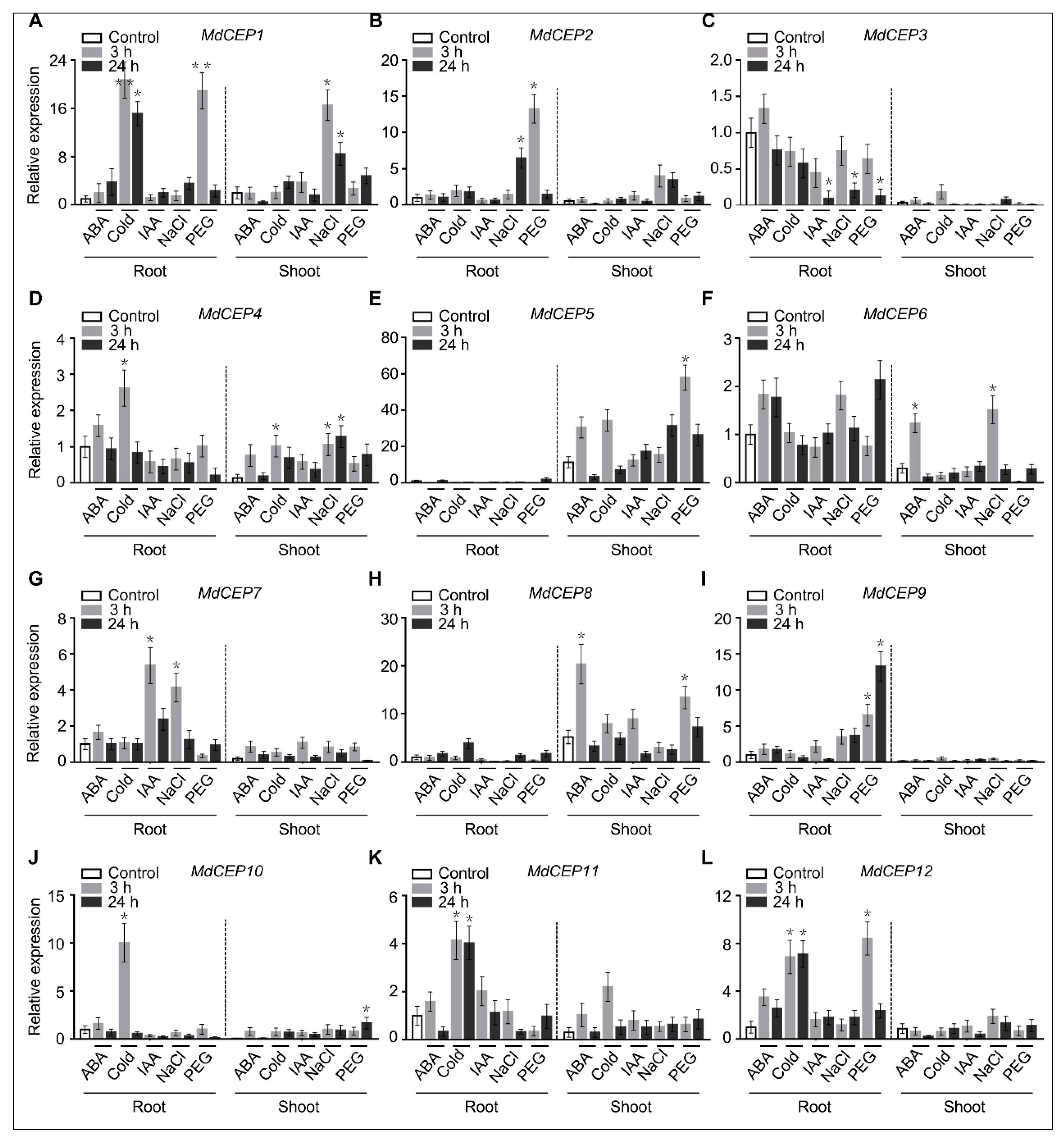
2.3. MdCEPs Display Distinct Spatial Expression Patterns in Apple
2.4. MdCEPs Involved in Multiple Biotic and Abiotic Stresses Response
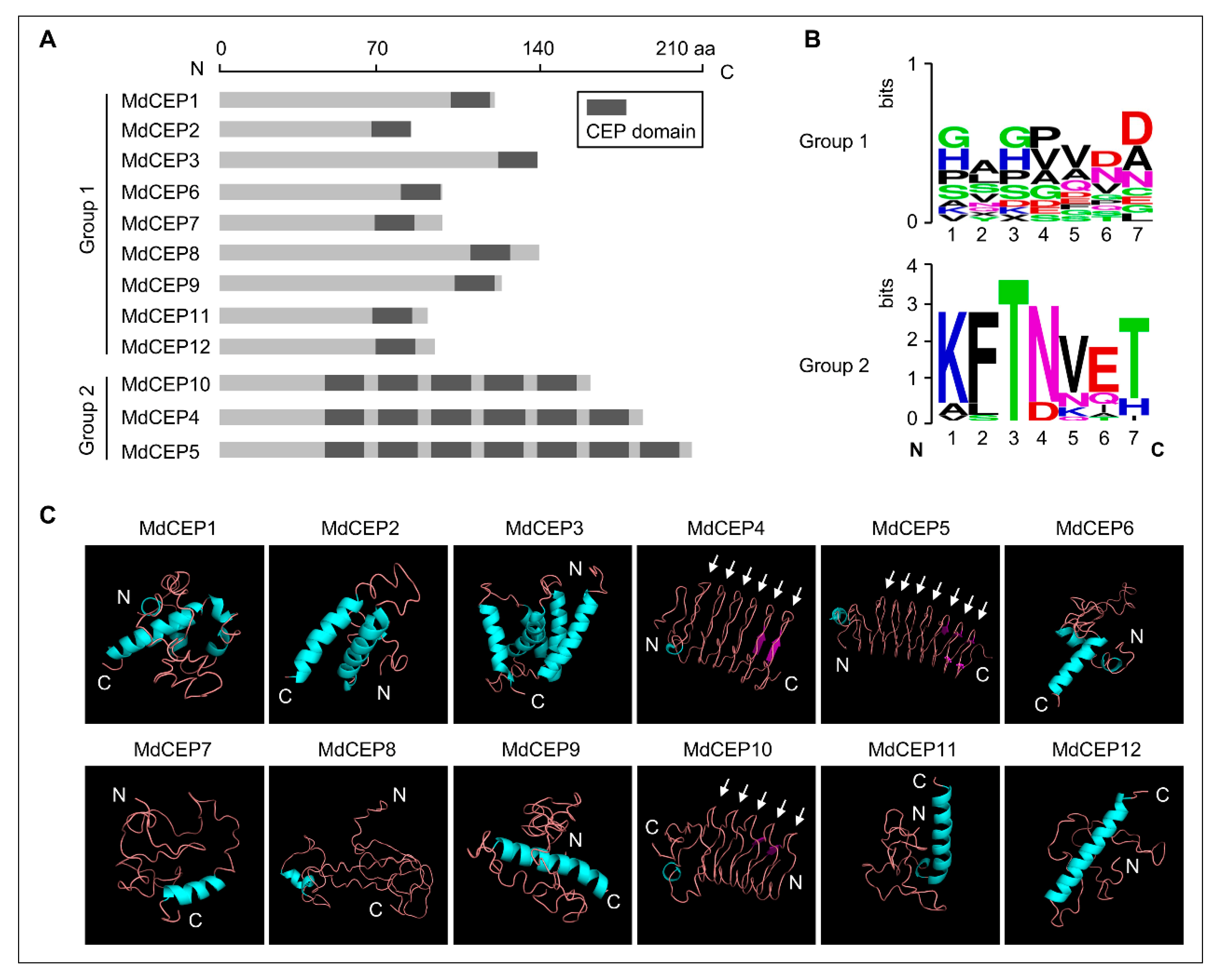
2.5. Distinct Post-Translational Modifications of MdCEPs in Apple
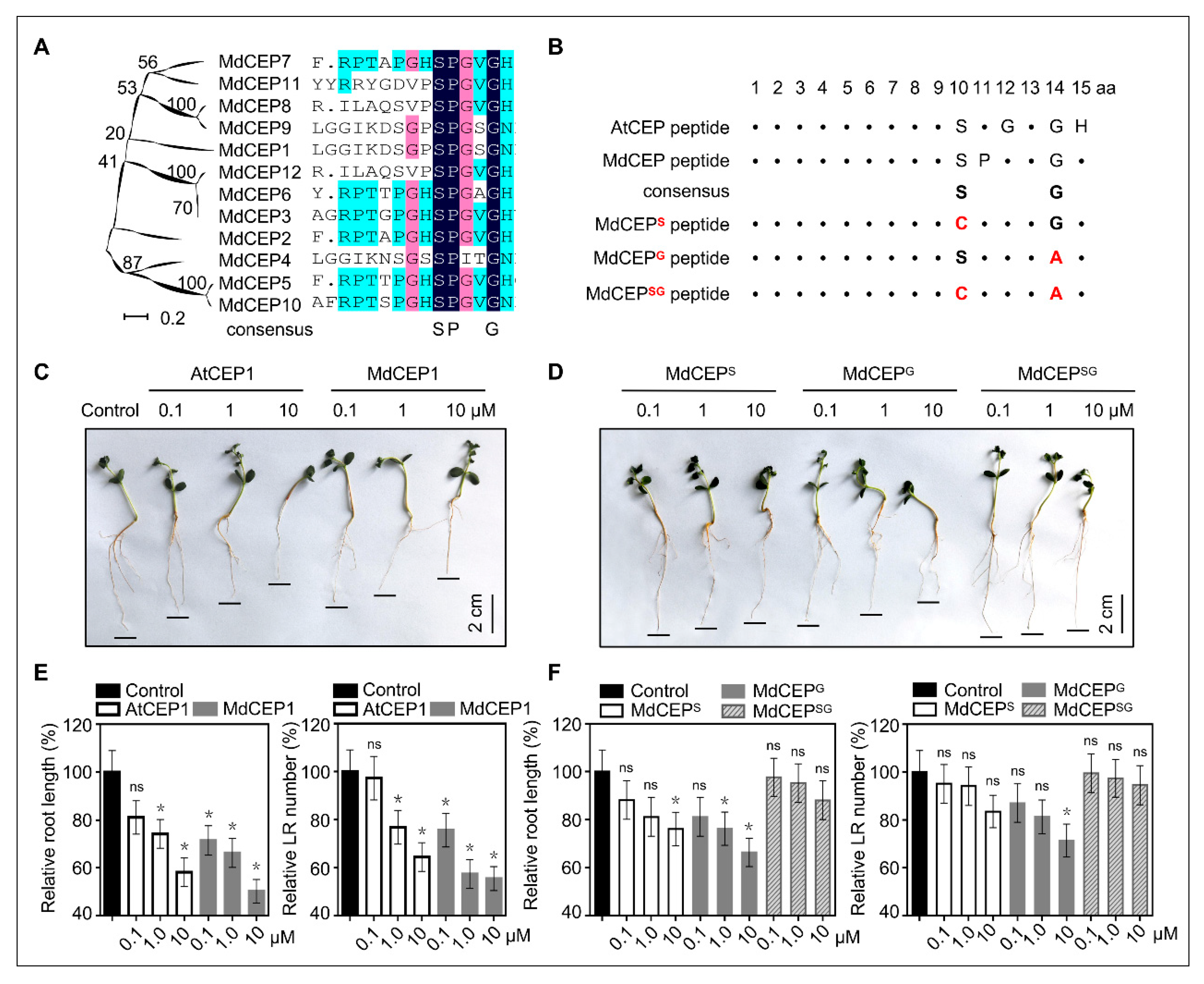
2.6. The Conserved Serine and Glycine Are Critical for Function Exertion of MdCEPs
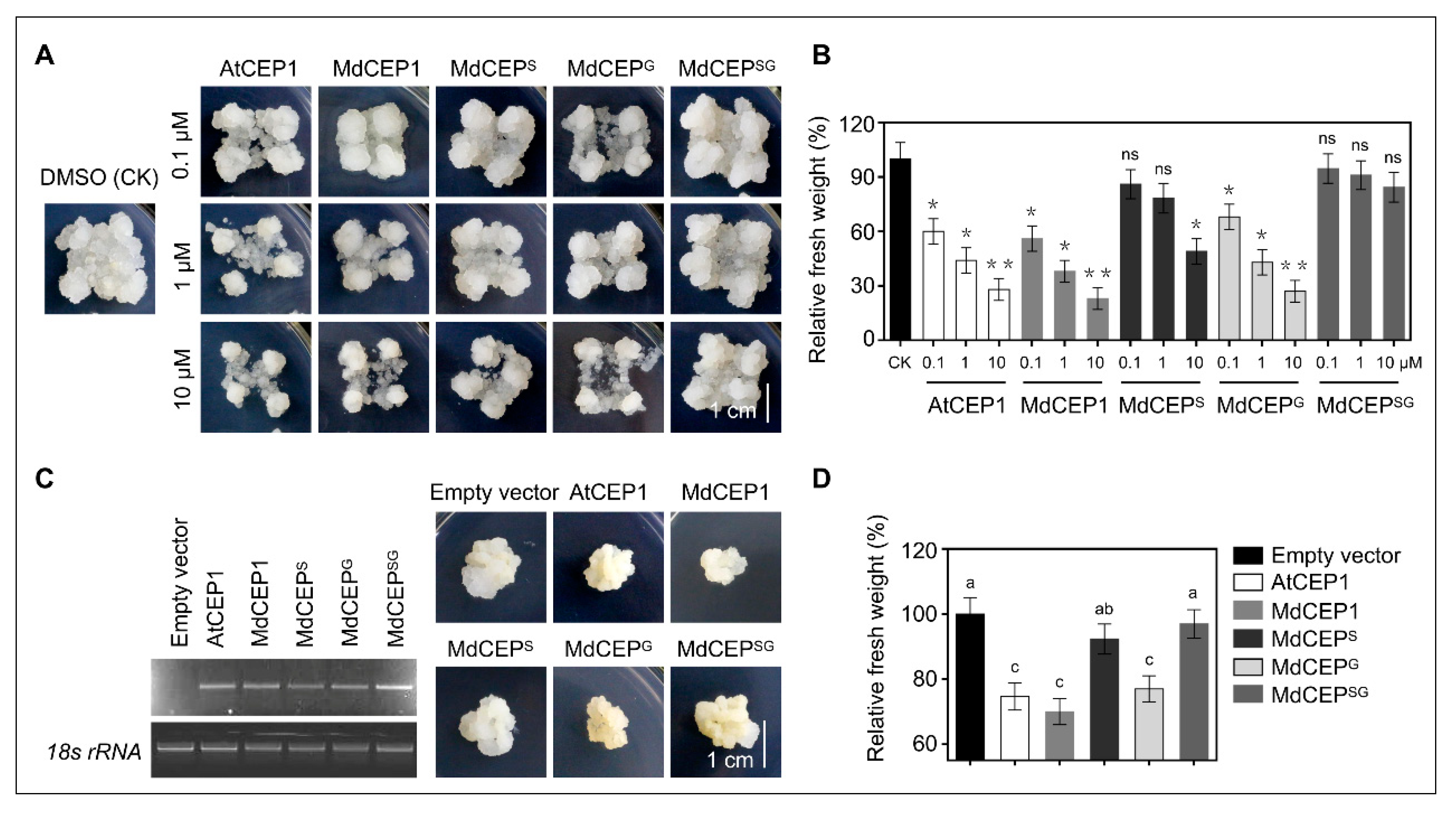
2.7. MdCEP Overexpression Leads to a Retarded Growth in Apple Callus
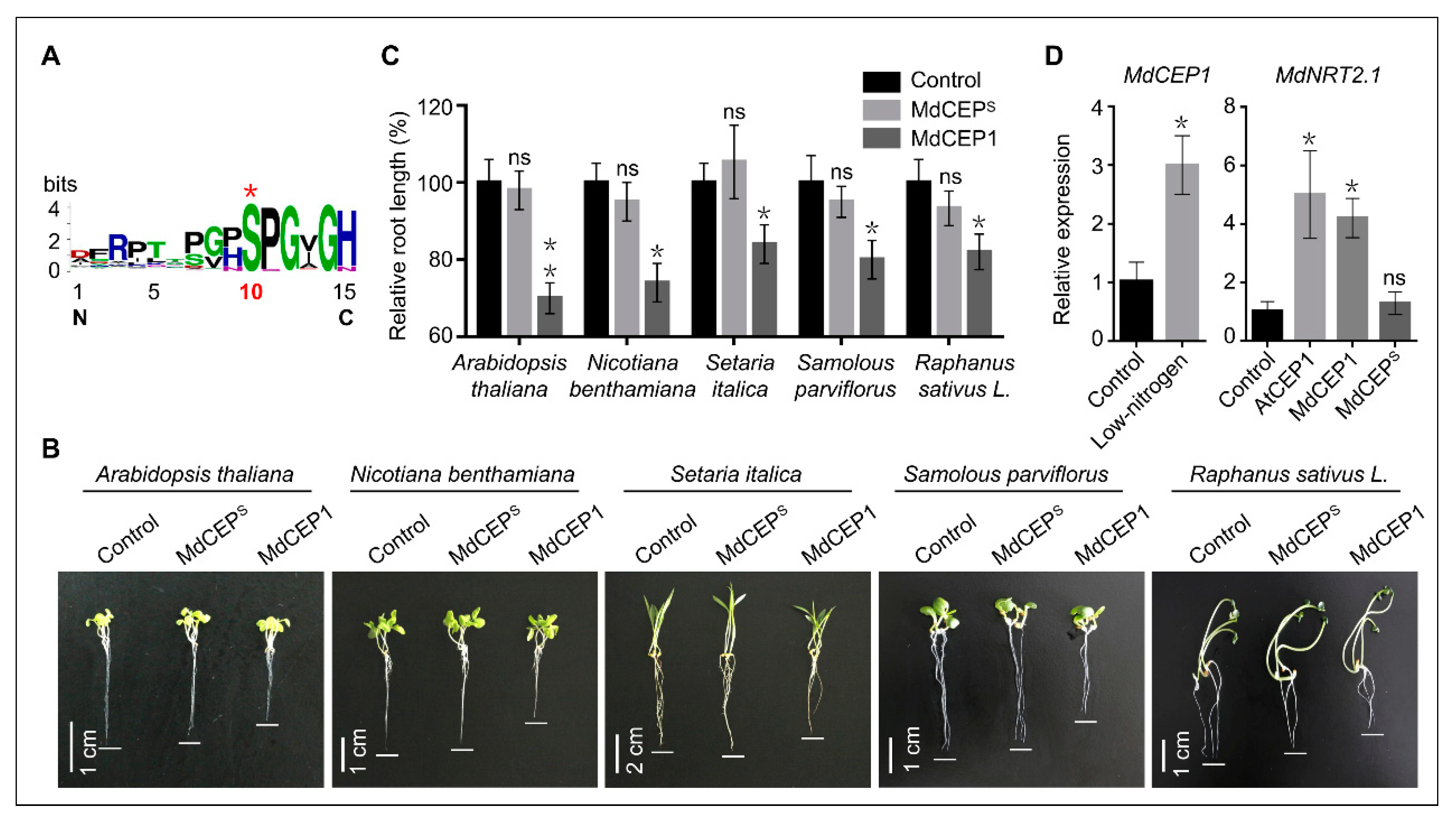
2.8. Serine is Evolutionarily Conserved in Higher Plants and MdCEPS Loses the Role in Inhibiting Root Growth
3. Discussion
4. Materials and Methods
4.1. Identification of MdCEPs in Apple
4.2. The Chromosomal Location and Gene Structures of MdCEPs
4.3. Sequence Alignment and Phylogenetic Analysis
4.4. Plant Growth and Treatment
4.5. RNA Extraction and qRT-PCR Analysis
4.6. Gene Cloning
4.7. In vitro and in vivo Growth Inhibition Assays
4.8. Data Availability
5. Statistical Tests
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CEP | C-terminally encoded peptide |
| PTMs | post-translationally modified peptides |
| CRPs | cysteine-rich peptides; |
| DMSO | dimethyl sulfoxide |
| qRT-PCR | quantitative real-time polymerase chain reaction |
References
- Gallagher, K.L.; Benfey, P.N. Not just another hole in the wall: Understanding intercellular protein trafficking. Genes. Dev. 2005, 19, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y. Regulation of short-distance transport of RNA and protein. Curr. Opin. Plant Biol. 2005, 8, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Stahl, Y.; Simon, R. Gated communities: Apoplastic and symplastic signals converge at plasmodesmata to control cell fates. J. Exp. Bot. 2013, 64, 5237–5241. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Smith, S.; De Smet, I. Small signaling peptides in Arabidopsis development: How cells communicate over a short distance. Plant Cell 2012, 24, 3198–3217. [Google Scholar] [CrossRef] [PubMed]
- Van Norman, J.M.; Breakfield, N.W.; Benfey, P.N. Intercellular communication during plant development. Plant Cell 2011, 23, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Czyzewicz, N.; Yue, K.; Beeckman, T.; De Smet, I. Message in a bottle: Small signalling peptide outputs during growth and development. J. Exp. Bot. 2013, 64, 5281–5296. [Google Scholar] [CrossRef] [PubMed]
- Butenko, M.A.; Vie, A.K.; Brembu, T.; Aalen, R.B.; Bones, A.M. Plant peptides in signalling: Looking for new partners. Trends Plant Sci. 2009, 14, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y. Small post-translationally modified Peptide signals in Arabidopsis. Arabidopsis Book 2011, 9, e0150. [Google Scholar] [CrossRef]
- Yamada, M.; Sawa, S. The roles of peptide hormones during plant root development. Curr. Opin. Plant Biol. 2013, 16, 56–61. [Google Scholar] [CrossRef]
- Ostrowski, M.; Kowalczyk, S. Plant signaling peptides. Cysteine-rich peptides. Postepy. Biochem. 2015, 61, 79–92. [Google Scholar]
- Marshall, E.; Costa, L.M.; Gutierrez-Marcos, J. Cysteine-rich peptides (CRPs) mediate diverse aspects of cell-cell communication in plant reproduction and development. J. Exp. Bot. 2011, 62, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Hemu, X.; Serra, A.; Darwis, D.A.; Cornvik, T.; Sze, S.K.; Tam, J.P. Peptidomic Identification of Cysteine-Rich Peptides from Plants. Methods Mol. Biol. 2018, 1719, 379–393. [Google Scholar] [PubMed]
- Tanco, S.; Gevaert, K.; Van Damme, P. C-terminomics: Targeted analysis of natural and posttranslationally modified protein and peptide C-termini. Proteomics 2015, 15, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, K.; Ogawa, M.; Matsubayashi, Y. Identification of a biologically active, small, secreted peptide in Arabidopsis by in silico gene screening, followed by LC-MS-based structure analysis. Plant J. 2008, 55, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Roberts, I.; Smith, S.; De Rybel, B.; Van Den Broeke, J.; Smet, W.; De Cokere, S.; Mispelaere, M.; De Smet, I.; Beeckman, T. The CEP family in land plants: Evolutionary analyses, expression studies, and role in Arabidopsis shoot development. J. Exp. Bot. 2013, 64, 5371–5381. [Google Scholar] [CrossRef] [PubMed]
- Tabata, R.; Sumida, K.; Yoshii, T.; Ohyama, K.; Shinohara, H.; Matsubayashi, Y. Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 2014, 346, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Huault, E.; Laffont, C.; Wen, J.; Mysore, K.S.; Ratet, P.; Duc, G.; Frugier, F. Local and systemic regulation of plant root system architecture and symbiotic nodulation by a receptor-like kinase. PLoS Genet. 2014, 10, e1004891. [Google Scholar] [CrossRef]
- Ohkubo, Y.; Tanaka, M.; Tabata, R.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Shoot-to-root mobile polypeptides involved in systemic regulation of nitrogen acquisition. Nat. Plants 2017, 3, 17029. [Google Scholar] [CrossRef]
- Roberts, I.; Smith, S.; Stes, E.; De Rybel, B.; Staes, A.; van de Cotte, B.; Njo, M.F.; Dedeyne, L.; Demol, H.; Lavenus, J.; et al. CEP5 and XIP1/CEPR1 regulate lateral root initiation in Arabidopsis. J. Exp. Bot. 2016, 67, 4889–4899. [Google Scholar] [CrossRef] [Green Version]
- Delay, C.; Imin, N.; Djordjevic, M.A. CEP genes regulate root and shoot development in response to environmental cues and are specific to seed plants. J. Exp. Bot. 2013, 64, 5383–5394. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Xu, Y.; Huang, J.; Jiang, Z.; Shu, H.; Wang, H.; Zhang, S. Global Identification, Classification, and Expression Analysis of MAPKKK genes: Functional Characterization of MdRaf5 Reveals Evolution and Drought-Responsive Profile in Apple. Sci. Rep. 2017, 7, 13511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus x domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Tabata, R.; Sawa, S. Maturation processes and structures of small secreted peptides in plants. Front Plant Sci. 2014, 5, 311. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, H.A.; Imin, N.; Djordjevic, M.A. Diversification of the C-TERMINALLY ENCODED PEPTIDE (CEP) gene family in angiosperms, and evolution of plant-family specific CEP genes. BMC Genom. 2014, 15, 870. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, H.; Matsubayashi, Y. Chemical synthesis of Arabidopsis CLV3 glycopeptide reveals the impact of hydroxyproline arabinosylation on peptide conformation and activity. Plant Cell Physiol. 2013, 54, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 1, 2–3. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, Z.; Zhang, D.; Huang, J.; Wu, C.; Yang, G.; Yan, K.; Zhang, S.; Zheng, C. CYSTM, a Novel Non-Secreted Cysteine-Rich Peptide Family, Involved in Environmental Stresses in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 423–438. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, X.; Song, Y.; Zhu, L.; Yu, Z.; Gan, L.; Zhou, S.; Liu, H.; Wen, F.; Zhu, C. NtLTP4, a lipid transfer protein that enhances salt and drought stresses tolerance in Nicotiana tabacum. Sci. Rep. 2018, 8, 8873. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Li, H.H.; Song, L.Q.; Su, L.; Liu, X.; You, C.X.; Wang, X.F.; Hao, Y.J. The molecular cloning and functional characterization of MdMYC2, a bHLH transcription factor in apple. Plant Physiol. Biochem. 2016, 108, 24–31. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Xu, Y.; Liu, L.; Guo, Y.; Yuan, X.; Man, X.; Liu, C.; Yang, G.; Huang, J.; Yan, K.; et al. The Importance of Conserved Serine for C-Terminally Encoded Peptides Function Exertion in Apple. Int. J. Mol. Sci. 2019, 20, 775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030775
Yu Z, Xu Y, Liu L, Guo Y, Yuan X, Man X, Liu C, Yang G, Huang J, Yan K, et al. The Importance of Conserved Serine for C-Terminally Encoded Peptides Function Exertion in Apple. International Journal of Molecular Sciences. 2019; 20(3):775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030775
Chicago/Turabian StyleYu, Zipeng, Yang Xu, Lin Liu, Yarong Guo, Xisen Yuan, Xinyu Man, Chang Liu, Guodong Yang, Jinguang Huang, Kang Yan, and et al. 2019. "The Importance of Conserved Serine for C-Terminally Encoded Peptides Function Exertion in Apple" International Journal of Molecular Sciences 20, no. 3: 775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030775