Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology


1
Department of Anatomy, Division of Basic Medical Science, Tokai University School of Medicine, Kanagawa 259-1193, Japan
2
Department of Anatomy, Tokyo Medical University, Tokyo 160-8402, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(4), 957; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040957
Submission received: 30 January 2019
/
Revised: 20 February 2019
/
Accepted: 20 February 2019
/
Published: 22 February 2019
(This article belongs to the Special Issue Reproductive Immunology: Cellular and Molecular Biology)
Abstract
:Substantial improvements in cancer treatment have resulted in longer survival and increased quality of life in cancer survivors with minimized long-term toxicity. However, infertility and gonadal dysfunction continue to be recognized as adverse effects of anticancer therapy. In particular, alkylating agents and irradiation induce testicular damage that results in prolonged azoospermia. Although damage to and recovery of spermatogenesis after cancer treatment have been extensively studied, there is little information regarding the role of differences in testicular immunology in cancer treatment-induced male infertility. In this review, we briefly summarize available rodent and human data on immunological differences in chemotherapy or radiotherapy.
1. Introduction and Immunological Tolerance of the Testis
The testis is an immune-privileged site at which immunogenic germ cells are protected from the detrimental effects of immune responses. The most commonly recognized mechanism for the immunological privilege is the blood–testis barrier (BTB), physically formed by the borders of adjacent Sertoli cells, limiting the access of germ cell antigens to interstitial immune cells and the passage of antibodies from the interstitium to the tubular lumen. Immunological privilege outside the BTB involves secretion of immunosuppressive factors mainly by macrophages, Sertoli cells, peritubular cells, and Leydig cells [1,2,3,4,5] (Table 1).
The testicular capacity to inhibit local immune responses has been confirmed in many studies; however, the mechanisms through which immune-privilege contributes to protection of spermatogenesis, a highly specialized process, have not been clearly defined. Elevated levels of intratesticular testosterone and/or progesterone may cause inhibition of local immune responses [6]. However, the testes contain high levels of steroidal molecules that are immunosuppressive by nature [7]. The expression of functional FasL by Sertoli cells [8] and/or by germ cells [9] as an active mechanism induces cell death via apoptosis, such as activated T cells in inflammation [10,11]. Furthermore, various anti-inflammatory cytokines, such as interleukin-10, are abundantly produced by testicular cells, particularly testicular macrophages [12], and members of the transforming growth factor-β family are highly expressed by Leydig and Sertoli cells [13]. The immune environment in the testes must be tightly controlled to maintain immune homeostasis for normal spermatogenesis. Disruption of immune homeostasis may result in autoimmune or infectious aspermatogenesis, thereby impairing testicular function. The mechanisms underlying the autoimmune inflammatory response and the mechanisms of orchitis have been broadly investigated and comprehensively reviewed in rats and mice [14,15,16].
The growing population of young cancer survivors and the trend toward postponing pregnancy until later in life have shifted researchers’ focus toward understanding treatment-induced sequelae, particularly the effects of cancer and/or treatment on fertility [17]. Although prepubertal testes do not undergo spermatogenesis and do not produce mature spermatozoa, the testes are sensitive to cytotoxic drugs and irradiation at this age. Testicular damage is drug specific and dose related, and the recovery of spermatogenesis varies following cytotoxic insults; additionally, the extent and speed of recovery are related to the agent used and the dose received [18,19,20,21]. Radiotherapy-induced testicular damage is similarly dose dependent, with speed of onset, chance of reversal, and time to recovery of spermatogenesis all related to the testicular dose of irradiation [17,22].
In this review, we aimed to outline immunological differences in cancer treatment-induced male infertility. Knowledge of this immunopathologic microenvironment will be useful to understand infertility and gonadal dysfunction as adverse effects of anticancer therapy.
2. Chemotherapy
Many drugs, particularly alkylating agents, have been shown to be gonadotoxic. Indeed, some chemotherapies used in the treatment of lymphoma or in preparation for bone transplantation have been shown to cause azoospermia, with feedback-raised follicle-stimulating hormone (FSH) levels in over 90% of men following cyclical chemotherapy [22,23].
Busulfan is a chemotherapeutic agent that is used to treat various malignant diseases, such as chronic myeloid leukemia and polycythemia vera [24,25]. Additionally, busulfan is also commonly used prior to hematopoietic stem cell transplantation [24]. Unlike other chemicals that destroy differentiated spermatogonia, busulfan is a potent agent that preferentially kills spermatogonial stem cells [26] and does not have any effect on DNA synthesis. Consequently, busulfan treatment is the most common method used to prepare recipients of spermatogonial stem cells transplantation [27,28] and to study spermatogonial stem cells kinetics and fertility recovery [29,30,31]. Busulfan can eliminate almost all endogenous germ cells in the recipient, creating an empty space in the spermatogonial stem cells niche; therefore, this drug has been used successfully to prepare recipients in mice [32,33], rats [34,35], pigs [36,37], dogs [38], hamsters [39], monkeys [40], and roosters [41].
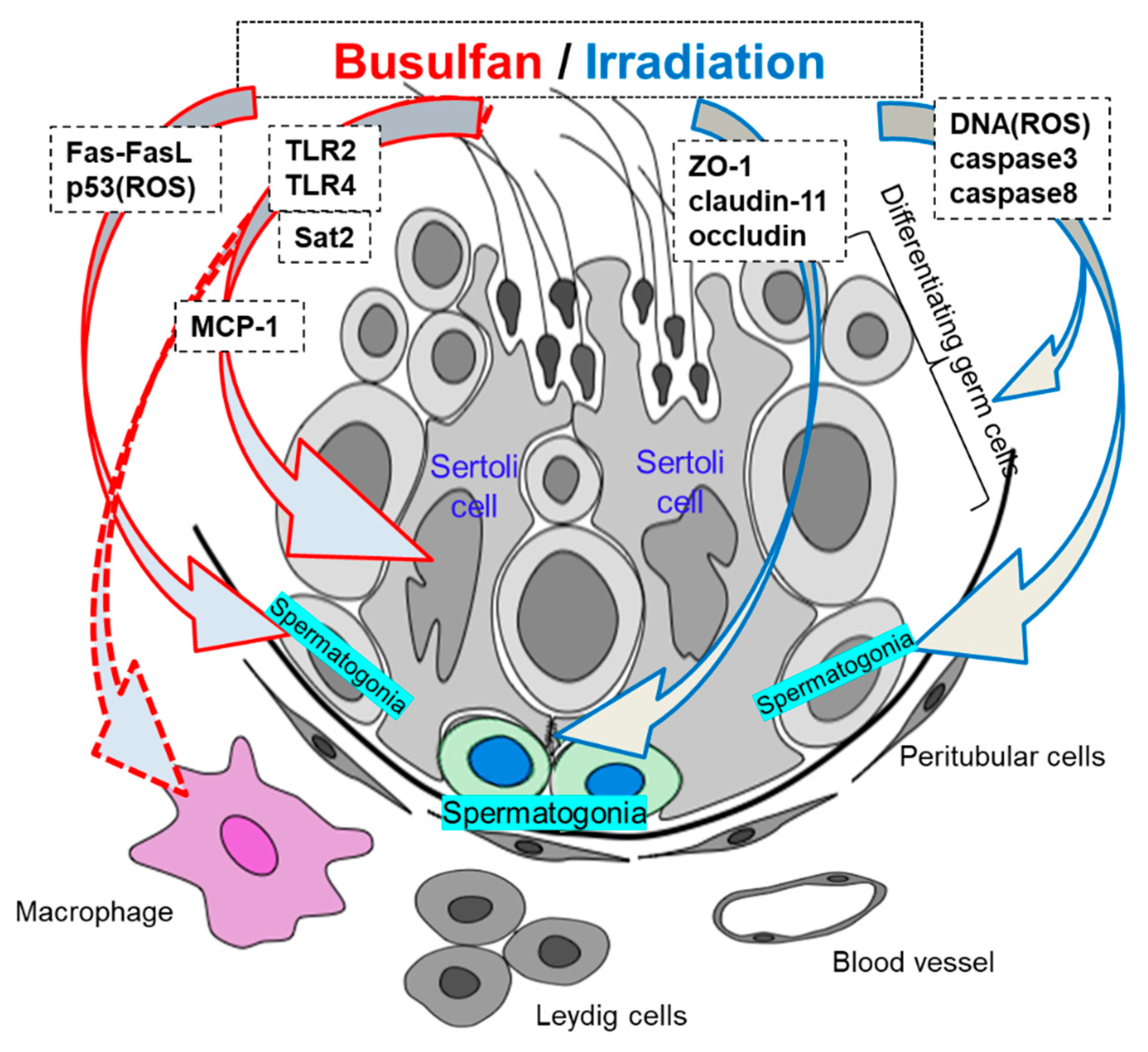
Busulfan treatment has been shown to disrupt spermatogenesis by damaging germ cells and Sertoli cells [42,43]. Although busulfan has been extensively studied regarding to induction of testicular damage through various cellular and molecular mechanisms, the effects of treatment on testicular immunology have not been thoroughly evaluated (Table 1). Choi et al. showed that busulfan may induce germ cell apoptosis through loss of c-kit signaling in a Fas/FasL- and p53-independent manner in 8–12-week-old mice [44]. Moreover, using the same adult mice, Li et al. demonstrated that p53 is a key protein with roles in busulfan-induced apoptosis through reactive oxygen species (ROS)-dependent activation of the extracellular signal-regulated kinase/p38 pathway, and decreased concentrations of deacetylated p53 result in spermatogonial cell resistance to apoptosis [45]. Xian et al. showed that spermidine/spermine N1-acetyltransferase 2 (Sat2) is present in adult mice testicular Sertoli cells and that its expression is significantly increased by busulfan treatment. Furthermore, upregulation of Sat2 by busulfan alters the growth and function of Sertoli cells and causes male infertility [46]. Zhang et al. demonstrated that busulfan-induced spermatogenic cell damage upregulates tumor necrosis factor (TNF) α and macrophage chemotactic protein (MCP) 1 expression in Sertoli cells and facilitates macrophage infiltration into the testes of 8–10 weeks age mice [47]. Additionally, damaged germ cells in busulfan-treated mice release endogenous Toll-like receptor (TLR) ligands to activate TLR2 and TLR4 in Sertoli cells, thus initiating endogenous inflammation in the testes [47,48]. Activation of TLRs induces inflammatory gene expression, which may facilitate injury repair and lead to further pathological conditions, such as autoimmune diseases [49,50,51].
Taken together, these studies suggested that the key factors mediating testicular immunology in busulfan-induced aspermatogenesis are Sertoli cells and macrophages, being similar with that in autoimmune orchitis excluding inflammatory reactions (Table 1, Figure 1).
Sinisi et al. investigated the occurrence of antisperm antibody (ASA) in 264 prepubertal male boys (ages 1.2–13 years) treated with chemotherapy and confirmed that 26 of these patients were ASA-positive [52]. Moreover, of the 26 ASA-positive boys, 24 had genital tract abnormalities, such as cryptorchidism, testicular torsion, and hypospadias, and two had leukemia with testicular infiltration [52]. Therefore, these findings suggested that chemotherapy did not modify antibody positivity. Only one experiment showed that remaining spermatogonia after 40 mg busulfan-treatment reacted strongly to IgG antibodies and that serum IgG levels increased in a manner corresponding with the increase in testicular IgG levels in adult (8–12-week-old) ICR mice [53]. Researchers demonstrated that serum IgG increased from 4 weeks after busulfan treatment, peaked at 7 weeks, and dropped rapidly to control concentrations after 8 weeks. In contrast, the testicular levels of IgG showed a gradual increase that accelerated after 3 weeks and peaked at 6 weeks, before dropping to control serum levels at 8 weeks. In our previous busulfan study in 4-week-old C57BL/6 mice, we examined serum anti-germ cell antibodies in 40 mg busulfan-treated mice and demonstrated that no anti-germ cell antibody production could be detected at 60 days after busulfan treatment (according to immunohistochemistry using serum samples and anti-mouse IgG) [48]. Moreover, we demonstrated that busulfan treatment progressively decreased the weight of the testes and the epididymal sperm count from day 60 to 120 and that 40 mg busulfan-induced aspermatogenesis was irreversible for at least 360 days [48,54,55]. We surmised that the primary cause of this aspermatogenesis involved factors other than ASA. We also showed that normalized macrophage migration and reduced expression of TLR2 and TLR4 after busulfan treatment could completely rescue the injured seminiferous epithelium and alleviate aspermatogenesis [48]. These differences may be related to the different experimental periods because it is well known that the infertility after busulfan is in a time- and dose-dependent manner, and is also possibly related with different species or ages of mice.
3. Radiotherapy
Irradiation has been shown to decrease spermatogenesis, alter the production of various hormones, and induce infertility. In both rodents and humans, the extent of testicular injury is directly related to the dose of irradiation delivered [56,57], and the germinal epithelium is very sensitive to radiation-induced damage [58], with changes in spermatogonia following doses as low as 0.1 Gy and permanent infertility after fractionated doses of 2 Gy and above [59].
Damage may be caused during direct irradiation of the testes or from scattered radiation during treatment directed at adjacent tissues. No recovery of spermatognesis was observed in 10 patients, mean age 32.9 years (range 24–40), receiving doses of 1.4–2.6 Gy after 17–43 months follow-up; however, recovery of fertility was observed in two patients with testicular radiation doses of 1.2 Gy and recovery of spermatogenesis was observed in all eight patients who received radiation doses of 0.28–0.9 Gy for testicular seminoma [60]. In postpubertal men, testicular doses of less than 0.2 Gy have no significant effect on FSH levels or sperm counts, whereas doses between 0.2 and 0.7 Gy cause transient dose-dependent increases in FSH and a reduction in sperm concentrations [61]. Notably, total body irradiation (TBI), used for bone marrow transplantation, is associated with appreciable gonadal toxicity, and previous studies have shown that 99.5% of men (mean age 31 years ranging from 11- to 62-years-old) who received 12.0 Gy TBI showed permanent infertility [23]. Moreover, TBI doses as low as 5–6 Gy could cause decreased spermatogenesis in the seminiferous tubules in prepubertal mice [62,63].
The deleterious effects of irradiation in biological systems are mainly mediated through the generation of ROS and cause lipid peroxidation in the cellular membrane, thereby inducing DNA damage in immature germ cells [64,65]. DNA damage caused by irradiation in premeiotic germ cells is detectable in primary spermatocytes and is still present in mature spermatozoa [64,66]. Furthermore, apoptosis of germ cells has been reported as a mechanism responsible for infertility in irradiated testes [67,68]. Some studies have indicated that irradiation-mediated oxidative stress induces apoptosis primarily in adult mouse and rat germ cells [69,70,71,72]. Additionally, irradiation-induced germ cell apoptosis depends on activation of caspase-3 in three-month-old rats [72], concomitant with increased expression of caspase-8 and decreased expression of caspase-9 in adult rats and prepubertal mice, respectively [72,73] (Table 1, Figure 1). However, irradiation-induced apoptosis does not occur in all types of testicular somatic cells. Some studies have shown that Sertoli cells and Leydig cells are resistant to irradiation-induced apoptosis [67,71,74,75,76], whereas some studies have described minimal changes in Sertoli cells [77,78].
Sertoli cells, through formation of the BTB, protect postmeiotic germ cells from exogenous toxicants introduced by testicular blood and lymph. Recently, some studies have demonstrated that Sertoli cell junctional proteins are the primary cellular targets of reproductive toxicants, such as cadmium chloride, dichlorodiphenyltrichloroethane, cisplatin, and bisphenol A [79,80,81]. In contrast, some reports have shown irradiation-induced BTB disruption with a decrease in zonula occludens-1 (ZO-1), occludin, and/or claudin-11 [82,83,84]. The integral membrane proteins claudin-11 [85,86] and occludin [87,88,89] and the adaptor protein ZO-1 [90,91], which all function in tight junctions, are critical components of the BTB. In both prepubertal and adult claudin11-knockout mice, the lumen of the seminiferous tubules are narrowed [92,93], round spermatids are the most mature germ cells, and increased germ cell apoptosis is observed [93]. In occludin-knockout mice, the seminiferous tubules are atrophied and have a Sertoli-cell-only phenotype at 40–60 weeks of age [94].
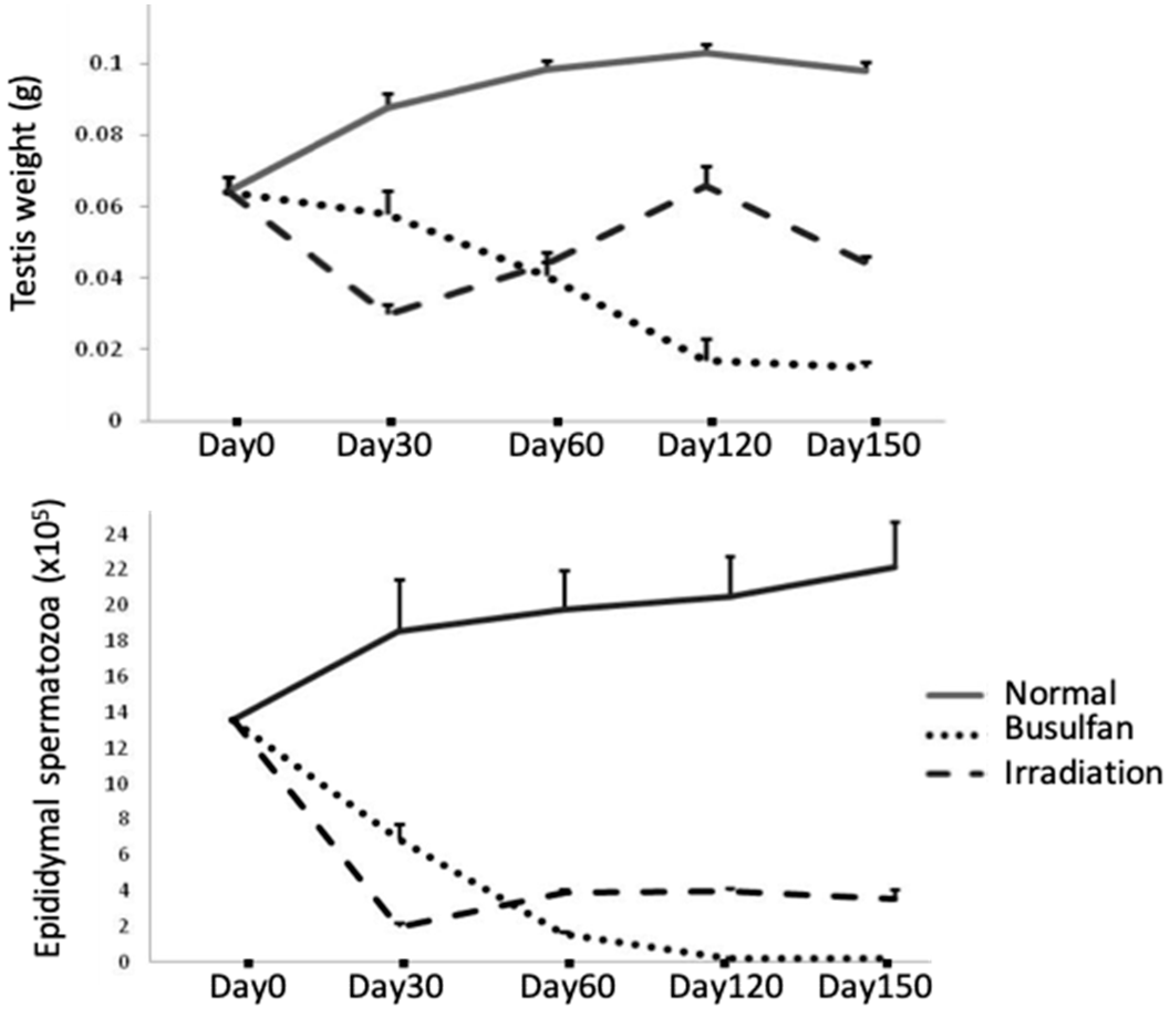
Breakdown of the BTB with its subsequent leukocytic infiltration of tubules can be found in human biopsy specimens from cases of idiopathic infertility and in models of testicular inflammation [95,96,97] (Table 1). An increase in BTB permeability is known to enhance ASA production, resulting in infertility in males [85,86,98]. Recently, we reported that single-dose TBI induced ASA that preferentially reacted with mature spermatids and spermatozoa in prepubertal mice [73]. Our results showed that 6 Gy of TBI induced a disruption of spermatogenesis with a decrease in inter-Sertoli tight junction mRNA levels and the production of ASA (Table 1). The contributions of claudin-11, occludin, and ZO-1 to BTB integrity were further determined in TBI-induced aspermatogenesis, and recovery of spermatogenesis was found to depend on the recovery of the above disorganized tight junctions. Furthermore, differences in busulfan- and irradiation-induced aspermatogenesis and the treatment duration in response to busulfan- and irradiation-induced aspermatogenesis were evaluated. Busulfan treatment in the same prepubertal mice was found to progressively decrease the weight of the testes and the epididymal sperm count from day 30 to day 120 (Figure 2); whereas an administered an oriental medicine completely rescued these effects on day 120 after busulfan treatment (testes weight: 0.100 ± 0.006 g; epididymal spermaotzoa: (21.680 ± 1.700) × 105 cells) [48]. In contrast, irradiation treatment induced significant decreases in the weights of the testes and epididymal sperm cell counts on day 30; marginal recovery was observed from day 60 to day 120, but further decreases in all parameters were noted on day 150 (Figure 2). Notably, supplementation with the above oriental medicine significantly restored the epididymal spermatozoa count and fertility on day 150 but not day 120 [73]. This belated recovery of spermatogenesis in the irradiated group compared with that in the busulfan-treated group suggested that the presence of ASAs may be the other immunological cause of aspermatogenesis. Although no leukocytic infiltration was detected in the irradiated testes, the breakdown of BTB and the immune responses against the testicular autoantigen are similar to that in the orchitis testes (Table 1).
4. Conclusion
From the above limited reported data, the impaired reproductive functions induced by cancer treatment including chemotherapy and radiotherapy are related with the different immune-pathophysiological conditions. Especially, with the breakdown of BTB by irradiation, germ cell autoantigens inside BTB might leak out repeatedly, leading to a continuous supply of autoantigens for immune stimulation with resultant ASA production and prolongation of the testicular inflammation. It is well known that the mechanisms of immune privilege in male reproductive organs are still far from being completely understood. The mechanisms underlying the autoimmune inflammatory response and the mechanisms of orchitis have been broadly investigated and comprehensively reviewed in rodent and human. Although damage to and recovery of spermatogenesis after cancer treatment have been extensively studied, there is little information regarding the role of differences in testicular immunology in cancer treatment-induced male infertility. Presently, the information on therapy for cancer treatment-induced male infertility is also limited. Because treatment with cytotoxic chemotherapy and radiotherapy indicated the increased FSH levels and reduced sperm counts, the effects of suppression of testosterone and gonadotrophin analogs on stimulation of spermatogonial differentiation, resulting in spermatogenic progression after cancer treatment, have been well demonstrated [99,100]. However, hormone suppression has multiple side effects and recovery occurs gradually, the application of hormone suppression treatments to enhance endogenous spermatogenic recovery has so far been successful in clinical trials [99]. Furthermore, chemotherapy and radiotherapy are often used in combination associated with greater testicular dysfunction and germinal epithelial damage. To lead to an availability of therapy on male infertility after cancer treatment, elucidation of the immunological mechanisms underlying aspermatogenesis by cancer treatment could be helpful.
Author Contributions
Writing—original draft preparation, N.Q.; writing—review and editing, M.I. and K.S. All authors critically reviewed and revised the manuscript and approved the final version.
Funding
This research was supported by a JSPS KAKENHI Grant (grant no. 15K08937) from the Ministry of Education Science Sports and Culture in Japan and 2018 Tokai University School of Medicine Research Aid.
Acknowledgments
We would like to thank Miyuki Kuramasu, Kenta Nagahori, Shogo Hayashi, Yoshie Hirayanagi, and Yoki Ogawa for their excellent secretarial and technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ASA | antisperm antibody |
| BTB | blood–testis barrier |
| FSH | follicle-stimulating hormone |
| MCP1 | macrophage chemotactic protein 1 |
| ROS | reactive oxygen species |
| Sat2 | spermidine/spermine N1-acetyltransferase 2 |
| TBI | total body irradiation |
| TLR | Toll-like receptor |
| TNF | tumor necrosis factor |
| ZO-1 | zonula occludens-1 |
References
- Head, J.R.; Billingham, R.E. Immune privilege in the testis. II. Evaluation of potential local factors. Transplantation 1982, 40, 269–295. [Google Scholar] [CrossRef]
- Pollanen, P.; Maddocks, S. Macrophages, lymphocytes and MHC II antigen in the ram and rat testis. J. Reprod. Fertil. 1988, 82, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, C.R.; Law, L.; Magnuson, J.A.; Griswold, M.D.; Magnuson, N.S. Suppression of lymphocyte proliferation by cultured Sertoli cells. J. Reprod. Immunol. 1988, 14, 27–40. [Google Scholar] [CrossRef]
- Itoh, M.; Terayama, H.; Naito, M.; Ogawa, Y.; Tainosho, S. Tissue microcircumstances for leukocytic infiltration into the testis and epididymis in mice. J. Reprod. Immunol. 2005, 67, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, S.; Meinhardt, A. The macrophages in testis function. J. Reprod. Immunol. 2017, 119, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Grossman, C. Interactions between the gonadal steroid and the immune system. Science 1985, 227, 157. [Google Scholar] [CrossRef]
- Russell, L.; Brinster, R. Ultrastructural observations of spermatogenesis following transplantation or rat testis cells into mouse seminiferous tubules. J. Androl. 1996, 17, 615–627. [Google Scholar]
- Bellgrau, D.; Gold, D.; Selawry, H.; Moore, J.; Franzusoff, A.; Duke, R.C. A role for CD95 ligand in preventing graft rejection. Nature 1995, 377, 630–632. [Google Scholar] [CrossRef]
- D’Alessio, A.; Riccioli, A.; Lauretti, P.; Padula, F.; Muciaccia, B.; De Cesaris, P. Testicular FasL is expressed by sperm cell. Proc. Natl. Acad. Sci. USA 2001, 98, 3316–3321. [Google Scholar] [CrossRef]
- O’Connell, J.; O’Sullivan, G.; Collins, K.; Shanahan, F. The Fas counter attack: Fas mediated T cell killing by colon cancer cells expressing Fas-ligand. J. Exp. Med. 1996, 184, 1075–1082. [Google Scholar] [CrossRef]
- Shimamoto, K.; Yamamoto, Y.; Sofikitis, N.; Miyagawa, I. Influence of anterior chamber eye-cells interferon-r and caspase inhibitor on the cellular immune reactions following transplantation of spermatogonia. J. Urol. 1999, 161, 345. [Google Scholar] [CrossRef]
- Watanabe, M.; Kashiwakura, Y.; Kusumi, N.; Tamayose, K.; Nasu, Y.; Nagai, A. Adeno-associated virus-mediated human IL-10 gene transfer suppresses the development of experimental autoimmune orchites. Gene Ther. 2005, 12, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Hedger, M.P.; Meinhardt, A. Cytokines and the immune-testicular axis. J. Reprod. Immunol. 2003, 58, 1–26. [Google Scholar] [CrossRef]
- Jacobo, P.; Guazzone, V.A.; Theas, M.S.; Lustig, L. Testicular autoimmunity. Autoimmun. Rev. 2011, 10, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Naito, M.; Terayama, H.; Hirai, S.; Qu, N.; Lustig, L.; Itoh, M. Experimental autoimmune orchitis as a model of immunological male infertility. Med. Mol. Morphol. 2012, 45, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Fijak, M.; Pilatz, A.; Hedger, M.P.; Nicolas, N.; Bhushan, S.; Michel, V.; Tung, K.S.K.; Schuppe, H.C.; Meinhardt, A. Infectious, inflammatory and ‘autoimmune’ male factor infertility: How do rodent models inform clinical practice? Hum. Reprod. Update 2018, 24, 416–441. [Google Scholar] [CrossRef] [PubMed]
- Meistirch, M.L. Effects of chemotherapy and radiotherapy on spermatogenesis in human. Fertil. Steril. 2013, 100, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, M.F.; Meistrich, M.L.; Fuller, L.M.; Cundiff, J.H.; Hagemeister, F.B.; Velasquez, W.S. Recovery of spermatogenesis after treatment for Hodgkin’s disease: Limiting dose of MOPP chemotherapy. J. Clin. Oncol. 1984, 2, 571–577. [Google Scholar] [CrossRef]
- Watson, A.R.; Rance, C.P.; Bain, J. Long term effects of cyclophosphamide on testicular function. Br. Med. J. Clin. Res. Ed. 1985, 291, 1457–1460. [Google Scholar] [CrossRef]
- Meistrich, M.L.; Chawla, S.P.; da Cunha, M.F.; Johnson, S.L.; Plager, C.; Papadopoulos, N.E. Recovery of sperm production after chemotherapy for osteosarcoma. Cancer 1989, 63, 2115–2123. [Google Scholar] [CrossRef] [Green Version]
- Pryzant, R.M.; Meistrich, M.L.; Wilson, G.; Brown, B.; McLaughlin, P. Long-term reduction in sperm count after chemotherapy with and without radiation therapy for non-Hodgkin’s lymphomas. J. Clin. Oncol. 1993, 11, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Rivkees, S.A.; Crawford, J.D. The relationship of gonaldal activity and chemotherapy-induced gonadal damage. JAMA 1988, 259, 2123–2125. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.E.; Hawley, J.; Levy, W.; Gooley, T.; Buckner, C.D.; Deeg, H.J.; Doney, K.; Storb, R.; Sullivan, K.; Witherspoon, R.; et al. Pregnancies following high-dose cyclophosphamide with or without high-dose busulfan or total-body irradiation and bone marrow transplantation. Blood 1996, 87, 3045–3052. [Google Scholar] [PubMed]
- Buggia, I.; Locatelli, F.; Regazzi, M.B.; Zecca, M. Busulfan. Ann. Pharmacother. 1994, 28, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Galaup, A.; Paci, A. Pharmacology of dimethanesulfonate alkylating agents: Busulfan and treosulfan. Expert Opin. Drug Metab. Toxicol. 2013, 9, 333–347. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, D.G.; Kramer, M.F. The effect of three alkylating agents on the seminiferous epithelium of rodents. I. Depletory effect. Virchows. Arch. B. Cell. Pathol. 1970, 4, 267–275. [Google Scholar] [PubMed]
- Shinohara, T.; Avarbock, M.R.; Brinster, R.L. Beta 1- and alpha 6-integrin are surface markers on mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 1999, 96, 5504–5509. [Google Scholar] [CrossRef]
- Ogawa, T. Spermatogonial transplantation: The principle and possible application. J. Mol. Med. 2001, 79, 368–374. [Google Scholar] [CrossRef]
- Perez-Crespo, M.; Pericuesta, E.; Perez-Cerezales, S.; Arenas, M.I.; Lobo, M.O.; Diaz-Gil, J.J. Effect of liver growth factor on both testicular regeneration and recovery of spermatogenesis in busulfan-treated mice. Reprod. Biol. Endocrinol. 2011, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Zohni, K.; Zhang, X.; Tan, S.L.; Chan, P.; Nagano, M.C. The efficiency of male fertility restoration is dependent on the recovery kinetics of spermatogonial stem cells after cytotoxic treatment with busulfan in mice. Hum. Reprod. 2012, 27, 44–53. [Google Scholar] [CrossRef]
- Gutierrez, K.; Glanzner, W.G.; Chemeris, R.O.; Rigo, M.L.; Comim, F.V.; Bordignon, V.; Gonçalves, P.B. Gonadotoxic effects of busulfan in two strains of mice. Reprod. Toxicol. 2016, 59, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Arechaga, J.M.; Avarbock, M.R.; Brinster, R.L. Transplantation of testis germinal cells into mouse seminiferous tubules. Int. J. Dev. Biol. 1997, 41, 111–122. [Google Scholar] [PubMed]
- Ogawa, T.; Ohmura, M.; Yumura, Y.; Sawada, H.; Kubota, Y. Expansion of murine spermatogonial stem cells through serial transplantation. Biol. Reprod. 2003, 68, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.X.; Short, R.V. Male germ cell transplantation in rats: Apparent synchronization of spermatogenesis between host and donor seminiferous epithelia. Int. J. Androl. 1995, 18, 326–330. [Google Scholar] [CrossRef]
- Zhang, Z.; Renfree, M.B.; Short, R.V. Successful intra- and interspecific male germ cell transplantation in the rat. Biol. Reprod. 2003, 68, 961–967. [Google Scholar] [CrossRef]
- Honaramooz, A.; Behboodi, E.; Hausler, C.L.; Blash, S.; Ayres, S.; Azuma, C.; Echelard, Y.; Dobrinski, I. Depletion of endogenous germ cells in male pigs and goats in preparation for germ cell transplantation. J. Androl. 2005, 26, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Bao, J.; Kong, Q.; Bai, Y.; Luo, F.; Zhou, S.; Wu, Y.; Huang, J. Effective production of recipient male pigs for spermatogonial stem cell transplantation by intratesticular injection with busulfan. Theriogenology 2017, 89, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Hur, T.Y.; Lee, S.H.; Ock, S.A.; Song, H.; Park, H.J.; Lee, R.; Sung, S.H.; Jhun, H.; Lee, W.Y. Dose-dependent effects of busulfan on dog testes in preparation for spermatogonial stem cell transplantation. Lab. Anim. Res. 2017, 33, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahdati, A.; Fathi, A.; Hajihoseini, M.; Aliborzi, G.; Hosseini, E. The regenerative effect of bone marrow-derived stem cells in spermatogenesis of infertile hamster. World J. Plast. Surg. 2017, 6, 18–25. [Google Scholar] [PubMed]
- Hermann, B.P.; Sukhwani, M.; Lin, C.C.; Sheng, Y.; Tomko, J.; Rodriguez, M.; Shuttleworth, J.J.; McFarland, D.; Hobbs, R.M.; Pandolfi, P.P.; et al. Characterization, cryopreservation, and ablation of spermatogonial stem cells in adult rhesus macaques. Stem Cells 2007, 25, 2330–2338. [Google Scholar] [CrossRef]
- Ghadimi, F.; Shakeri, M.; Zhandi, M.; Zaghari, M.; Piryaei, A.; Moslehifar, P.; Rajabinejad, A. Different approaches to establish infertile rooster. Anim. Reprod. Sci. 2017, 186, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Bhartiya, D.; Sriraman, K.; Mallick, A. Underlying mechanisms that restore spermatogenesis on transplanting healthy niche cells in busulphan treated mouse testis. Stem Cell Rev. Rep. 2016, 12, 682–697. [Google Scholar] [CrossRef] [PubMed]
- Bhartiya, D.; Anand, S. Effects of oncotherapy on testicular stem cells and niche. Mol. Hum. Reprod. 2017, 23, 654–655. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Ok, D.W.; Kwon, D.N.; Chung, J.I.; Kim, H.C.; Yeo, S.M.; Kim, T.; Seo, H.G.; Kim, J.H. Murine male germ cell apoptosis induced by busulfan treatment correlates with loss of c-kit-expression in a Fas/FasL- and p53-independent manner. FEBS Lett. 2004, 575, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; He, X.; Zhuang, M.; Niu, B.; Wu, C.; Mu, H.; Tang, F.; Cui, Y.; Liu, W.; Zhao, B.; et al. Melatonin ameliorates busulfan-induced spermatogonial stem cell oxidative apoptosis in mouse testes. Antioxid. Redox Signal. 2018, 28, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Xian, Y.; Wu, M.; Liu, Y.; Hao, J.; Wu, Y.; Liao, X.; Li, G. Increased Sat2 expression is associated with busulfan-induced testicular Sertoli cell injury. Toxicol. In Vitro 2017, 43, 47–57. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, T.; Deng, T.; Xiong, W.; Lui, P.; Li, N.; Chen, Y.; Han, D. Damaged spermatogenic cells induce inflammatory gene expression in mouse Sertoli cells through the activation of Toll-like receptors 2 and 4. Mol. Cell. Endocrinol. 2013, 365, 162–173. [Google Scholar] [CrossRef]
- Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Nagahori, K.; Hayashi, S.; Ogawa, Y.; Terayama, H.; Suyama, K.; Naito, M.; Sakabe, K.; et al. Gosha-Jinki-Gan recovers spermatogenesis in mice with busulfan-induced aspermatogenesis. Int. J. Mol. Sci. 2018, 19, 2606. [Google Scholar] [CrossRef]
- Piccinini, A.M.; Midwood, K.S. DAMPening inflammation by modulating TLR signalling. Mediat. Inflamm. 2010, 2010, 672395. [Google Scholar] [CrossRef]
- Yu, L.; Wang, L.; Chen, S. Endogenous toll-like receptor ligands and their biological significance. J. Cell. Mol. Med. 2010, 14, 2592–2603. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, S.; Tchatalbachev, S.; Lu, Y.; Frohlich, S.; Fijak, M.; Vijayan, V.; Chakraborty, T.; Meinhardt, A. Differential activation of inflammatory pathways in testicular macrophages provides a rationale for their subdued inflammatory capacity. J. Immunol. 2015, 194, 5455–5464. [Google Scholar] [CrossRef] [PubMed]
- Sinisi, A.A.; D’Apuzzo, A.; Pasquali, D.; Venditto, T.; Esposito, D.; Pisano, G.; De Bellis, A.; Ventre, I.; Papparelia, A.; Perrone, L.; et al. Antisperm antibodies in prepubertal boys treated with chemotherapy for malignant or non-malignant diseases and in boys with genital tract abnormalities. Int. J. Androl. 1997, 20, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Song, H.; Kwon, D.N.; Cho, S.K.; Kang, S.J.; Yoe, S.M.; Kim, H.C.; Lee, H.T.; Park, C.; Kim, J.H. Significant IgG-immunoreactivity of the spermatogonia of the germ cell-depleted testis after busulfan treatment. Anim. Reprod. Sci. 2006, 91, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Qu, N.; Naito, M.; Li, J.; Terayama, H.; Hirai, S.; Itoh, M. Xenogeneic and endogenous spermatogenesis following transplantation of rat germ cells into testes of immunocompetent mice. Reprod. Fertil. Dev. 2012, 24, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Hirayanagi, Y.; Qu, N.; Hirai, S.; Naito, M.; Terayama, H.; Hayashi, S.; Hatayama, N.; Kuramasu, M.; Ogawa, Y.; Itoh, M. Busulfan pretreatment for transplantation of rat spermatogonia differentially affects immune and reproductive systems in male recipient mice. Anat. Sci. Int. 2015, 90, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Kuban, D.A.; Meistrich, M.L.; Cheung, R.; Hussey, D.H. Chapter 27—The Testicle. In Radiation Oncology, 9th ed.; Cox, J.D., Ang, K.K., Eds.; Mosby Elsevier: Philadelphia, PA, USA, 2010; pp. 653–675. [Google Scholar]
- Abuelhija, M.; Weng, C.C.; Shetty, G.; Meistrich, M.L. Rat models of post-irradiation recovery of spermatogenesis: Interstrain differences. Andrology 2013, 1, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Osterberg, E.C.; Ramasamy, R.; Masson, P.; Brannigan, R.E. Current practices in fertility preservation male cancer patients. Urol. Ann. 2014, 6, 13–17. [Google Scholar] [CrossRef]
- Howell, S.J.; Shalet, S.M. Spermaotgenesis after cancer treatment: Damage and recovery. J. Natl. Cancer Inst. Monogr. 2005, 34, 12–17. [Google Scholar] [CrossRef]
- Centola, G.M.; Keller, J.W.; Henzler, M.; Rubin, P. Effect of low-dose testicular irradiation on sperm count and fertility in patients with testicular seminoma. J. Androl. 1994, 15, 608–613. [Google Scholar]
- Kinsella, T.J.; Trivette, G.; Rowland, J.; Sorace, R.; Miller, R.; Fraass, B. Long-term follow-up of testicular function following radiation therapy for early-stage Hodgkin’s disease. J. Clin. Oncol. 1989, 7, 718–724. [Google Scholar] [CrossRef]
- Vergouwen, R.P.; Huiskamp, R.; Bas, R.J.; Roepers-Gajadien, H.L.; de Jong, F.H.; van Eerdenburg, F.J.; Davids, J.A.; de Rooij, D.G. Radiosensitivity of testicular cells in the prepubertal mouse. Radiat. Res. 1994, 139, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Adhikari, J.S.; Rizvi, M.A.; Chaudhury, N.K. Radioprotective potential of melatonin against Co γ-ray-induced testicular injury in male C57BL/6 mice. J. Biomed. Sci. 2015, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Cordelli, E.; Fresegna, A.M.; Leter, G.; Eleuteri, P.; Spano, M.; Villani, P. Evaluation of DNA damage in different stages of mouse spermatogenesis after testicular X irradiation. Radiat. Res. 2003, 160, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Ergur, B.U.; Kiray, M.; Pekcetin, C.; Bagriyanik, H.A.; Erbil, G. Protective effect of erythropoietin pretreatment in testicular ischmiareperfusion injury in rats. J. Pediatr. Surg. 2008, 43, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Xu, B.; Yang, M.; Li, Z.; Zhang, Y.; Jiang, D. Protection by hydrogen against gamma ray-induced testicular damage in rats. Basic Clin. Pharmacol. Toxicol. 2013, 112, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Pino-Lataillade, G.; Velez de la Calle, J.F.; Viguier-Martinez, M.C. Influence of germ cells upon Sertoli cells during continuous low-dose rate gamma-irradiation of adult rats. Mol. Cell. Endocrinol. 1988, 58, 51–63. [Google Scholar] [CrossRef]
- Hasegawa, M.; Wilson, G.; Russell, L.D. Radiation-induced cell death in the mouse testis: Relationship to apoptosis. Radiat. Res. 1997, 147, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Park, J.S.; Kim, Y.J. Differential expression of Prx I and II in mouse testis and their up-regulation by radiation. Biochem. Biophys. Res. Commun. 2002, 296, 337–342. [Google Scholar] [CrossRef]
- Ma, Y.; Nie, H.; Sheng, C.; Chen, H.; Wang, B.; Liu, T.; Shao, J.; He, X.; Zhang, T.; Zheng, C.; et al. Roles of oxidative stress in synchrotron radiation X-ray-induced testicular damage of rodents. Int. J. Physiol. Pathophysiol. Pharmacol. 2012, 4, 108–114. [Google Scholar] [PubMed]
- Aktoz, T.; Caloglu, M.; Uurut-Caloglu, V.; Yalcin, O.; Aydogdu, N.; Vurlu, D.; Arda, E.; Inci, O. Histopathological and biochemical comparisons of the protective effects of amifostine and L-carnitine against radiation-induced acute testicular toxicity in rats. Andrologia 2017, 49, e12754. [Google Scholar] [CrossRef]
- Silva, A.M.; Correia, S.; Casalta-Lopes, J.E.; Mamede, A.C.; Cavaco, J.E.; Botelho, M.F.; Socorro, S.; Maia, C.J. The protective effect of regucalcin against radiation-induced damage in testicular cells. Life Sci. 2016, 164, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nagahori, K.; Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Hayashi, S.; Ogawa, Y.; Hatayama, N.; Terayama, H.; Suyama, K.; et al. The traditional Japanese medicine Goshajinkigan makes a recovery from irradiation-induced aspermatogenesis in mice. 2019; manuscript under review. [Google Scholar]
- Vergouwen, R.P.; Huiskamp, R.; Bas, R.J. Radiosensitivity of testicular cells in the fetal mouse. Radiat. Res. 1995, 141, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Guitton, N.; Brouazin-Jousseaume, V.; Dupaix, A.; Jegou, B.; Chenal, C. Radiation effect on rat Sertoli cell function in vitro and in vivo. Int. J. Radiat. Biol. 1999, 75, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, A.V.; Almeida, F.R.; Weng, C.C.; Shetty, G.; Meistrich, M.L.; Chiarini-Garcia, H. Spermatogonial behavior in rats during radiation-induced arrest and recovery after hormone suppression. Reproduction 2013, 146, 363–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kangasniemi, M.; Huhtaniemi, I.; Meistrich, M.L. Failure of spermatogenesis to recover despite the presence of a spermatogonia in the irradiated LBNF1 rat. Biol. Reprod. 1996, 54, 1200–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuttlesworth, G.A.; de Rooij, D.G.; Huhtaniemi, I.; Reissmann, T.; Russell, L.D.; Shetty, G.; Meistrich, M.L. Enhancement of a spermatogonial proliferation and differentiation in irradiated rats by gonadotropin-releasing hormone antagonist administration. Endocrinology 2000, 141, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Monsees, T.K.; Franz, M.; Gebhardt, S.; Winterstein, U.; Schill, W.B.; Hayatpour, J. Sertoli cells as a target for reproductive hazards. Andrologia 2000, 32, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Defamie, N.; Mograbi, B.; Roger, C.; Cronier, L.; Malassine, A.; Brucker-Davis, F.; Fenichel, P.; Segretain, D.; Pointis, G. Disruption of gap junctional intercellular communication by lindane is associated with aberrant localization of connexin43 and zonula occludens-1 in 42GPA9 Sertoli cells. Carcinogenesis 2001, 22, 1537–1542. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, C.; Tilloy-Ellul, A.; Chevalier, S.; Charuel, C.; Pointis, G. Sertoli cell junctional proteins as early targets for different classes of reproductive toxicants. Reprod. Toxicol. 2004, 18, 413–421. [Google Scholar] [CrossRef]
- Wang, X.W.; Ding, G.R.; Shi, C.H.; Zeng, L.H.; Liu, J.Y.; Li, J.; Zhao, T.; Chen, Y.B.; Guo, G.Z. Mechanisms involved in the blood-testis barrier increased permeability induced by EMP. Toxicology 2010, 276, 58–63. [Google Scholar] [CrossRef]
- Son, Y.; Heo, K.; Bae, M.J.; Lee, C.G.; Cho, W.S.; Kim, S.D.; Yang, K.; Shin, I.S.; Lee, M.Y.; Kim, J.S. Injury to the blood-testis barrier after low-dose-rate chronic radiation exposure in mice. Radiat. Prot. Dosim. 2015, 167, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, T.; Shao, J.; Sheng, C.; Hong, Y.; Ying, W.; Xia, W. Antioxidant protects blood-testis barrier against synchrotron radiation X-ray-induced disruption. Spermatogenesis 2015, 5, e1009313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinhardt, A.; Hedger, M.P. Immunological, paracrine and endocrine aspects of testicular immune privilege. Mol. Cell. Endocrinol. 2011, 335, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Proc. Natl. Acad. Sci. USA 1999, 96, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, K.; Sasaki, H.; Fujimoto, K.; Furuse, M.; Tsukita, S. Claudin-11/OSP-based tight junction of myelin sheaths in brain and Sertoli cells in testis. J. Cell Biol. 1999, 145, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Hirase, T.; Itoh, M.; Nagafuchi, A.; Yonemura, S.; Tsukita, S.; Tsukita, S. Occludin: A novel integral membrane protein localizing at tight junction. J. Cell Biol. 1993, 123, 1777–1788. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, B.R.; Siliciano, J.D.; Mooseker, M.S.; Goodenough, D.A. Identification of ZO-1: A high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J. Cell Biol. 1986, 103, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The tight junction protein ZO-1 establishes a link between the transmembrane protein occluding and the actin cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar] [CrossRef]
- Gow, A.; Southwood, C.M.; Li, J.S.; Pariali, M.; Riordan, G.P.; Brodie, S.E.; Danias, J.; Bronstein, J.M.; Kachar, B.; Lazzarini, R.A. CNS myelin and Sertoli cell tight junction strands are absent in Osp/claudin-11 null mice. Cell 1999, 99, 649–659. [Google Scholar] [CrossRef]
- Mazaud-Guittot, S.; Meugnier, E.; Pesenti, S.; Wu, X.; Vidal, H.; Gow, A.; Le Magueresse-Battistoni, B. Claudin 11 deficiency in mice results in loss of the Sertoli cell epithelial phenotype in the testis. Biol. Reprod. 2010, 82, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Saitou, M.; Furuse, M.; Sasaki, H.; Schulzke, J.D.; Fromm, M.; Takano, H.; Noda, T.; Tsukita, S. Complex phenotype of mice lacking occluding, a component of tight junction strands. Mol. Biol. Cell. 2000, 11, 4131–4142. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H. Changes in the blood-testis barrier of the guinea-pig in relation to histological damage following iso-immunization with testis. J. Reprod. Fertil. 1970, 22, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Nistal, M.; Riestra, M.L.; Paniagua, R. Focal orchitis in undescended testes: Discussion of pathogenetic mechanisms of tubular atrophy. Arch. Pathol. Lab. Med. 2002, 126, 64–69. [Google Scholar] [PubMed]
- Naito, M.; Itoh, M. Patterns of infiltration of lymphocyte into the testis under normal and pathological conditions in mice. Am. J. Reprod. Immunol. 2008, 59, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Q.; Cheng, C.Y. A seamless trespass: Germ cell migration across the seminiferous epithelium during spermatogenesis. J. Cell. Biol. 2007, 178, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Shetty, G.; Meistrich, M.L. Hormonal approaches to preservation and restoration of male fertility after cancer treatment. J. Natl. Cancer Inst. Monogr. 2005, 2005, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Meistrich, M.L.; Shetty, G. Inhibition of spermatogonial differentiation by testosterone. J. Androl. 2003, 24, 135–148. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Immunological differences in the testes after cancer treatment.

Figure 2.
Testes weights and epididymal spermatozoa numbers in normal and busulfan-/irradiation-treated mice.
Figure 2.
Testes weights and epididymal spermatozoa numbers in normal and busulfan-/irradiation-treated mice.


{kind=link}
{kind=link}
Table 1.
Testicular immune factors in normal and cancer-treated mice.
| Immunosuppressive Factors in Normal Testis | Local Function | Testicular Immunology Consequences after Busulfan-Treatment | Testicular Immunology Consequences after Irradiation-Treatment | Testicular Immunology Consequences in Autoimmune Orchitis | ||
|---|---|---|---|---|---|---|
| Germ cells | transforming growth factor β | Leydig cell steroidogenesis ↓ | ||||
| Fas ligand | apoptosis of Fas-bearing lymphocyte | ↑ or (-) | ||||
| interferon-γ | Leydig cell steroidogenesis ↓ | |||||
| tumor necrosis factor α | Leydig cell steroidogenesis ↓ or ↑ | |||||
| Fas ↑ caspase3-8 ↑ | Fas ↑ | caspase 3–8 ↑ | ||||
| Fas ↑ | apoptosis through oxidative stress | Bax ↑ | caspase 9 ↑ | |||
| p53-ROS ↑ caspase3 ↑ | DNA damage | |||||
| Sertoli cells | activin | mitogenesis of lymphocytes ↓ | ||||
| inhibin | mitogenesis of lymphocytes ↑ | |||||
| interleukin-6 | meiotic DNA synthesis of germ cell | ↑ ? | ↑ | |||
| Fas ligand | apoptosis of Fas-bearing lymphocyte | |||||
| transforming growth factor β | inhibin secretion ↑ | |||||
| TNFα↑ MCP-1 ↑ TLR2,4 ↑ | ZO-1, occludin, claudin-11 ↓ | occludin, claudin-11 ↓ | ||||
| Sat2 ↑ | ||||||
| Leydig cells | testosterone | ↓ | ↓ | ↓ | ||
| protein S | Leydig cell steroidogenesis ↓ | |||||
| insulin-like growth factor-1 | testosterone secretion ↑ | |||||
| Fas ligand | apoptosis of Fas-bearing germ cell | |||||
| interleukin-10 | immune privilege | |||||
| transforming growth factor β | contractility of myoid cell | |||||
| Leydig cell apoptosis | ||||||
| Testicular | interleukin-10 | inhibition of T cell-mediated immune response response | ||||
| macrophages | interferon-γ | Fas ligand expression by Sertoli cell ↑ | ↑ | |||
| interleukin-6 | radioprotection of germ cell by Sertoli cell | ↑ ? | ↑ | |||
| tumor necrosis factor α | Fas ligand expression by Sertoli cell ↓ or ↑ | ↑ | ↑ | |||
| macrophage infiltration (+) | macrophage infiltration (-) | macrophage infiltration (+) | ||||
| Others | ASA? | ASA (+) | T cells·B cells infiltration (+) | |||
| ASA (+) | ||||||
↑ indicated increase and ↓ indicated decrease; ? indicated different opinion.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qu, N.; Itoh, M.; Sakabe, K. Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology. Int. J. Mol. Sci. 2019, 20, 957. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040957
AMA Style
Qu N, Itoh M, Sakabe K. Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology. International Journal of Molecular Sciences. 2019; 20(4):957. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040957
Chicago/Turabian StyleQu, Ning, Masahiro Itoh, and Kou Sakabe. 2019. "Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology" International Journal of Molecular Sciences 20, no. 4: 957. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040957
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.