Botulinum Neurotoxin-A Injected Intrastriatally into Hemiparkinsonian Rats Improves the Initiation Time for Left and Right Forelimbs in Both Forehand and Backhand Directions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
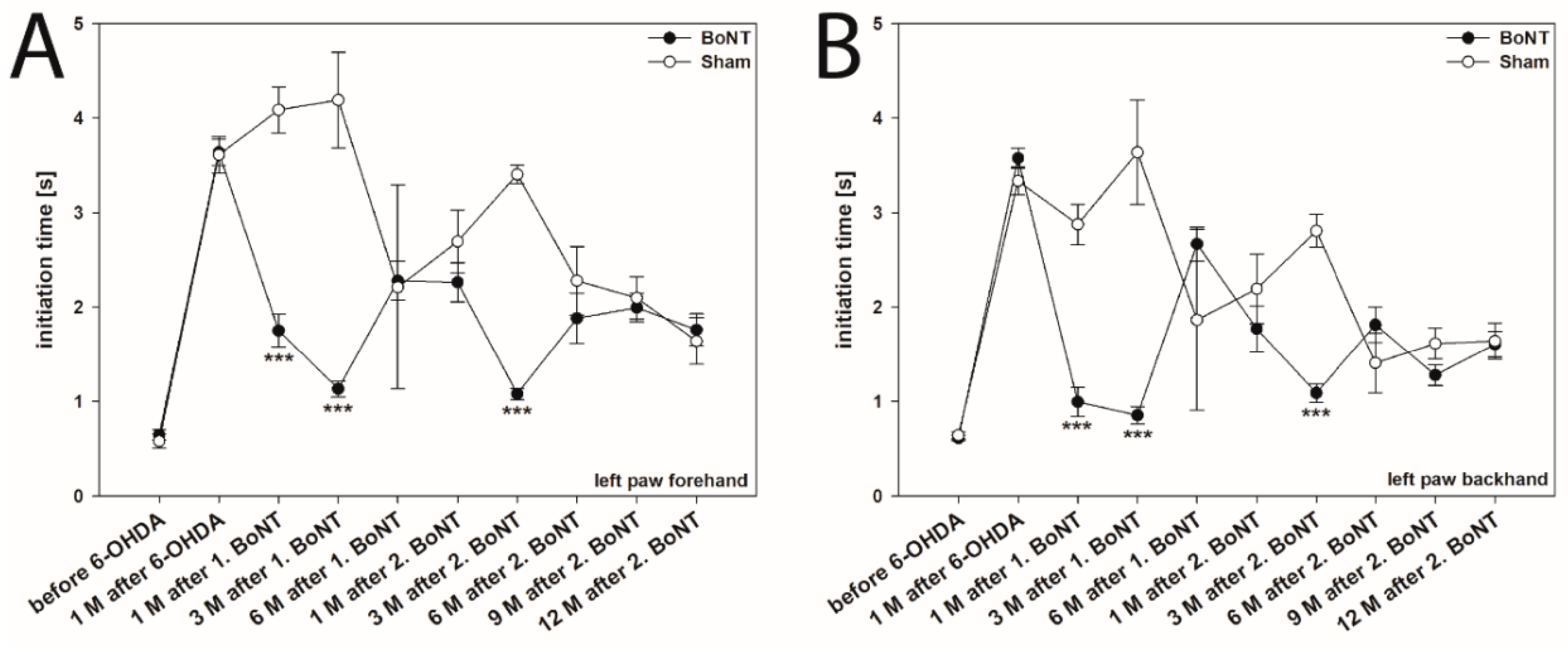
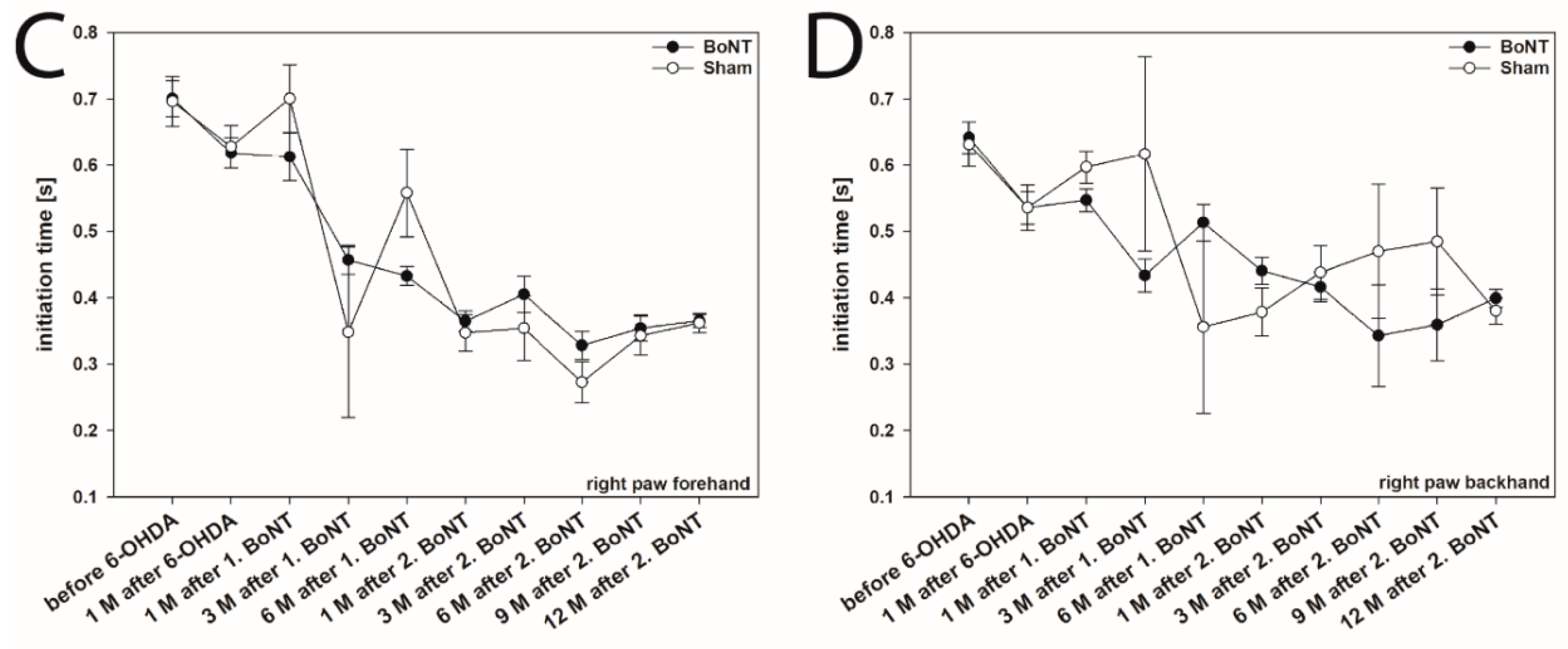
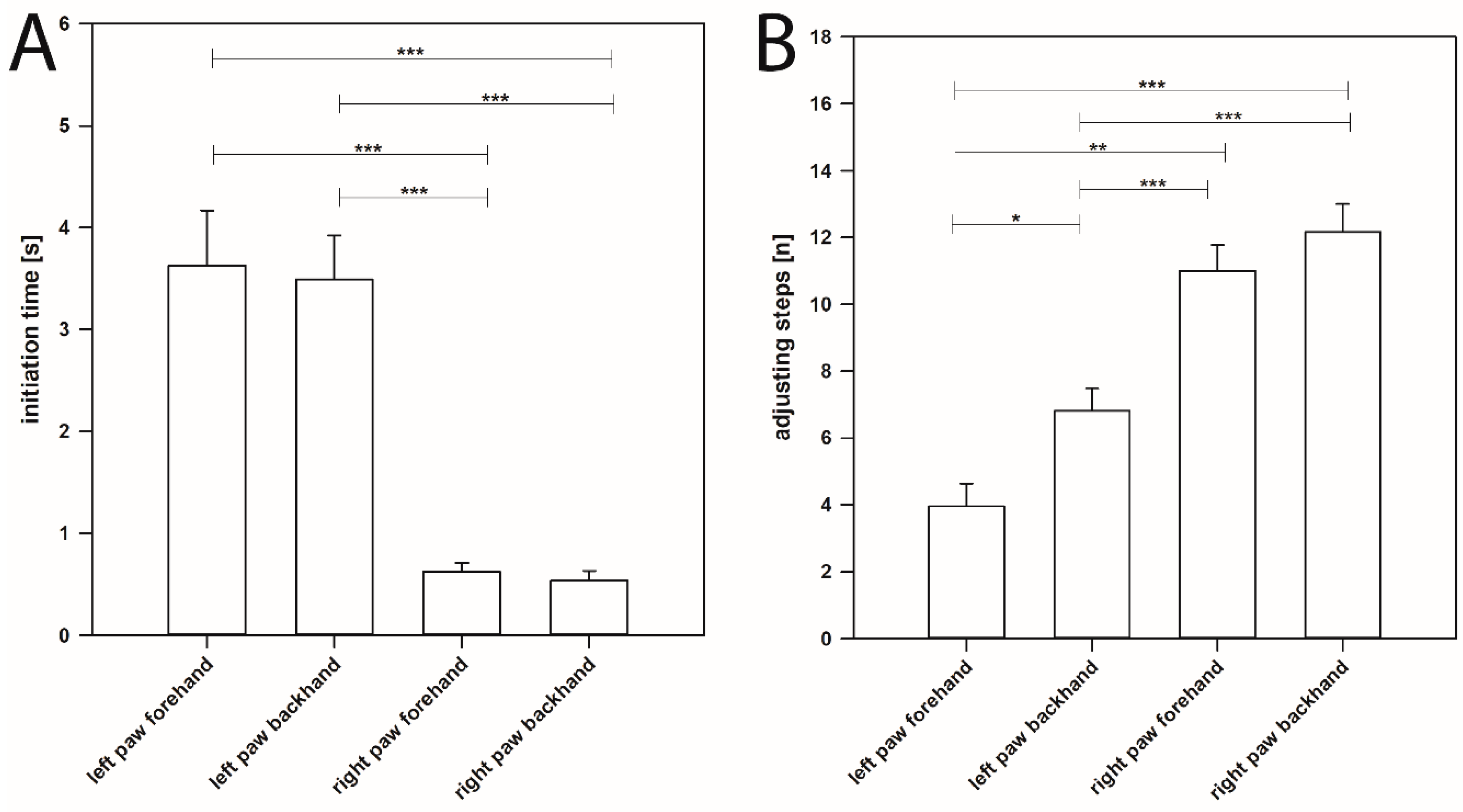
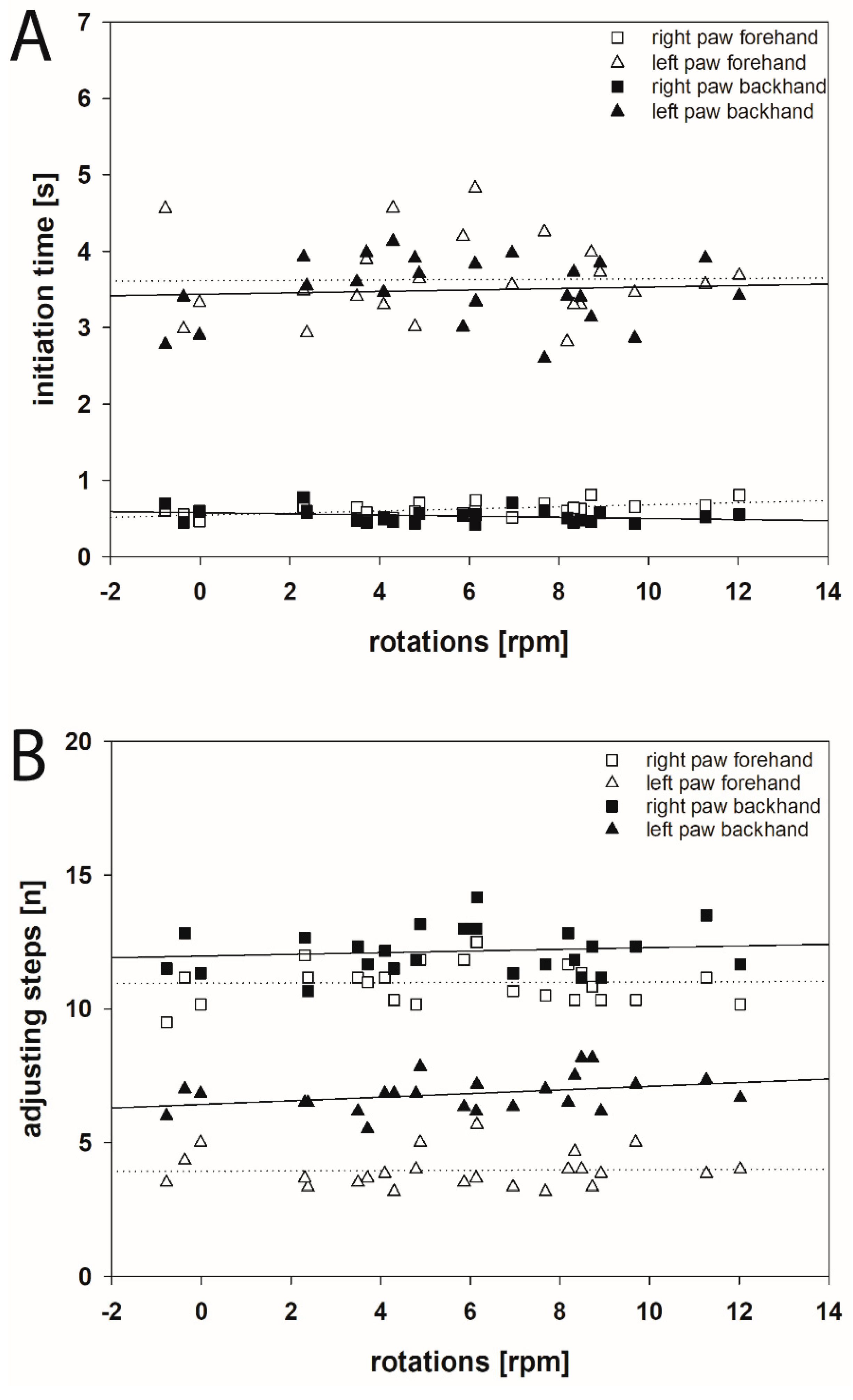
2.1. Initiation Time
2.1.1. Hemi-PD Rats
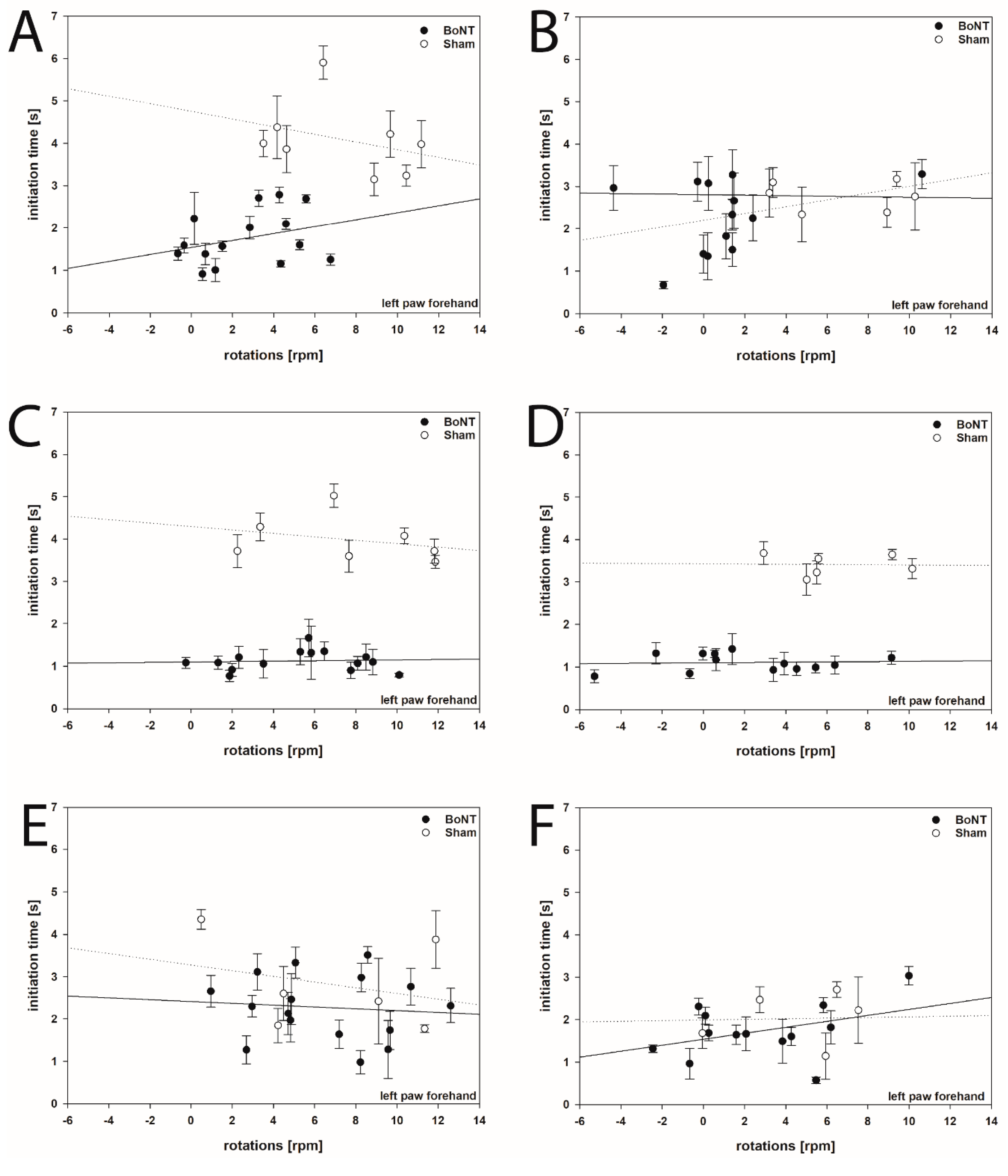
2.1.2. Hemi-PD Rats Treated with BoNT-A
2.2. Adjusting Steps
2.3. Initiation Time, Adjusting Steps, and Apomorphine-Induced Rotations
2.3.1. Hemi-PD Rats
2.3.2. Hemi-PD Rats Treated with BoNT-A or Sham BoNT-A Injections
3. Discussion
3.1. Initiation Time in Hemi-PD Rats According to Olsson et al. (1995)
3.2. Newly Introduced Initiation Time in Hemi-PD Rats
3.3. Comparison to Initiation Time According to Olsson et al.
3.4. Initiation Time in Hemi-PD Rats During Post-Lesion Survival
3.5. Initiation Time in Hemi-PD Rats Following Intrastriatal BoNT-A Injection
3.6. Correlating Initiation Time, Adjusting Steps, and Apomorphine-Induced Rotations
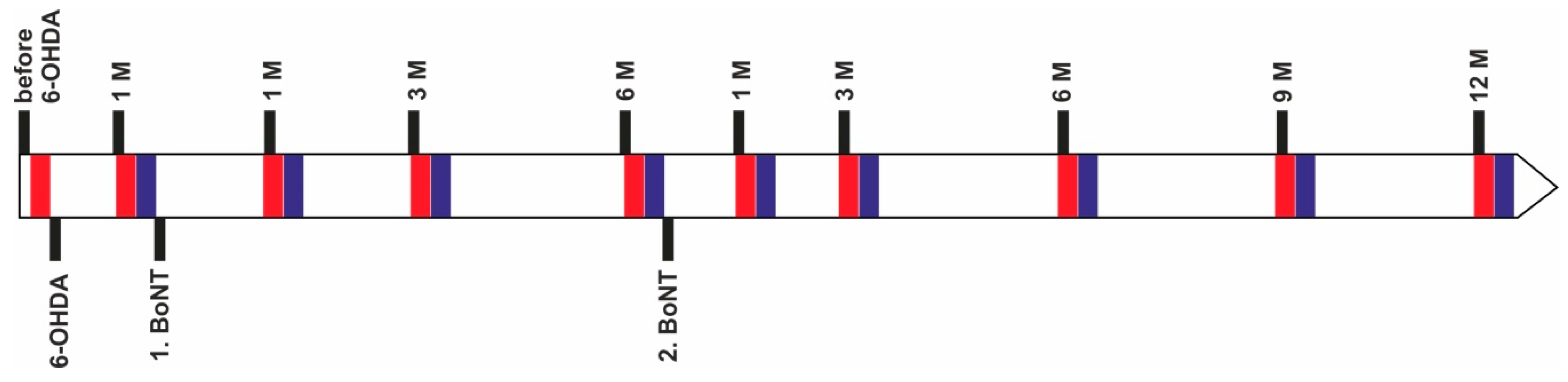
4. Materials and Methods
4.1. Animals
4.2. Induction of Hemiparkinsonism
4.3. Injection of BoNT-A into the Striatum
4.4. Behavioral Testing
4.4.1. Apomorphine-Induced Rotation Test
4.4.2. Stepping Test
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 6-OHDA | 6-hydroxydopamine |
| AS | adjusting steps |
| BoNT-A | botulinum neurotoxin-A |
| BW | body weight |
| CPu | caudate-putamen (striatum) |
| DA | dopamine |
| hemi-PD | hemiparkinsonian |
| ir | immunoreactive |
| IT | initiation time |
| M | month |
| MFB | medial forebrain bundle |
| PD | Parkinson’s disease |
| rpm | rotations per minute |
| SNpc | substantia nigra pars compacta |
| TH | tyrosine hydroxylase |
| VTA | ventral tegmental area |
References
- Fang, X.; Sugiyama, K.; Akamine, S.; Namba, H. Improvements in motor behavioral tests during deep brain stimulation of the subthalamic nucleus in rats with different degrees of unilateral parkinsonism. Brain Res. 2006, 1120, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Massano, J.; Bhatia, K.P. Clinical approach to Parkinson’s disease: Features, diagnosis, and principles of management. Cold Spring Harb. Perspect. Med. 2012, 2, a008870. [Google Scholar] [CrossRef] [PubMed]
- Kirik, D.; Rosenblad, C.; Björklund, A. Characterization of behavioral and neurodegenerative changes following partial lesions of the nigrostriatal dopamine system induced by intrastriatal 6-hydroxydopamine in the rat. Exp. Neurol. 1998, 152, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Hornykiewicz, O. Parkinson’s disease and the adaptive capacity of the nigrostriatal dopamine system. Adv. Neurol. 1993, 60, 140–147. [Google Scholar] [PubMed]
- Olsson, M.; Nikkhah, G.; Bentlage, C.; Björklund, A. Forelimb akinesia in the rat Parkinson model: Differential effects of dopamine agonists and nigral transplants as assessed by a new stepping test. J. Neurosci. 1995, 15, 3863–3875. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A. Thirty years of dopamine research. Adv. Neurol. 1993, 60, 1–10. [Google Scholar] [PubMed]
- Ungerstedt, U. 6-hydroxy-dopamine induced degeneration of central monoamine neurons. Eur. J. Pharmacol. 1968, 5, 107–110. [Google Scholar] [CrossRef]
- Ljungberg, T.; Ungerstedt, U. Sensory inattention produced by 6-hydroxydopamine-induced degeneration of ascending dopamine neurons in the brain. Exp. Neurol. 1976, 53, 585–600. [Google Scholar] [CrossRef]
- Ungerstedt, U.; Arbuthnott, G.W. Quantitative recording of rotational behavior in rats after 6-hydroxy-dopamine lesions of the nigrostriatal dopamine system. Brain Res. 1970, 24, 485–493. [Google Scholar] [CrossRef]
- Sachs, C.; Jonsson, G. Mechanisms of action of 6-hydroxydopamine. Biochem. Pharmacol. 1975, 24, 1–8. [Google Scholar] [CrossRef]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.; Benabid, A.L.; Sadoul, R.; Verna, J.M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
- Schwarting, R.K.W.; Huston, J.P. The unilateral 6-hydroxydopamine lesion model in behavioral brain research. Analysis of functional deficits, recovery and treatments. Prog. Neurobiol. 1996, 50, 275–331. [Google Scholar] [CrossRef]
- Schwarting, R.K.; Huston, J.P. Behavioral and neurochemical dynamics of neurotoxic meso-striatal dopamine lesions. Neurotoxicology 1997, 18, 689–708. [Google Scholar] [PubMed]
- Blesa, J.; Phani, S.; Jackson-Lewis, V.; Przedborski, S. Classic and new animal models of Parkinson’s disease. J. Biomed. Biotechnol. 2012, 2012, 845618. [Google Scholar] [CrossRef] [PubMed]
- Betarbet, R.; Sherer, T.B.; Greenamyre, J.T. Animal models of Parkinson’s disease. Bioessays 2002, 24, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Gerin, C. Behavioral improvement and dopamine release in a Parkinsonian rat model. Neurosci. Lett. 2002, 330, 5–8. [Google Scholar] [CrossRef]
- Schmidt, R.H.; Björklund, A.; Stenevi, U.; Dunnett, S.B.; Gage, F.H. Intracerebral grafting of neuronal cell suspensions. III. Activity of intrastriatal nigral suspension implants as assessed by measurements of dopamine synthesis and metabolism. Acta Physiol. Scand. Suppl. 1983, 522, 19–28. [Google Scholar] [PubMed]
- Nikkhah, G.; Duan, W.M.; Knappe, U.; Jödicke, A.; Björklund, A. Restoration of complex sensorimotor behavior and skilled forelimb use by a modified nigral cell suspension transplantation approach in the rat parkinson model. Neuroscience 1993, 56, 33–43. [Google Scholar] [CrossRef]
- Ichitani, Y.; Okamura, H.; Matsumoto, Y.; Nagatsu, I.; Ibata, Y. Degeneration of the nigral dopamine neurons after 6-hydroxydopamine injection into the rat striatum. Brain Res. 1991, 549, 350–353. [Google Scholar] [CrossRef]
- Jeon, B.S.; Jackson-Lewis, V.; Burke, R.E. 6-Hydroxydopamine Lesion of the Rat Substantia Nigra: Time Course and Morphology of Cell Death. Neurodegeneration 1995, 4, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Sauer, H.; Björklund, A. Dopaminergic neuronal degeneration and motor impairments following axon terminal lesion by intrastriatal 6-hydroxydopamine in the rat. Neuroscience 1996, 72, 641–653. [Google Scholar] [CrossRef]
- Yuan, H.; Sarre, S.; Ebinger, G.; Michotte, Y. Histological, behavioural and neurochemical evaluation of medial forebrain bundle and striatal 6-OHDA lesions as rat models of Parkinson’s disease. J. Neurosci. Methods 2005, 144, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Shi, L.H.; Luo, F.; Woodward, D.J. High frequency stimulation of the subthalamic nucleus improves treadmill locomotion in unilateral 6-hydroxydopamine lesioned rats. Brain Res. 2003, 983, 174–184. [Google Scholar] [CrossRef]
- Dowd, E.; Monville, C.; Torres, E.M.; Dunnett, S.B. The Corridor Task: A simple test of lateralised response selection sensitive to unilateral dopamine deafferentation and graft-derived dopamine replacement in the striatum. Brain Res. Bull. 2005, 68, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Rozas, G.; Guerra, M.J.; Labandeira-García, J.L. An automated rotarod method for quantitative drug-free evaluation of overall motor deficits in rat models of parkinsonism. Brain Res. Protoc. 1997, 2, 75–84. [Google Scholar] [CrossRef]
- Shi, L.H.; Woodward, D.J.; Luo, F.; Anstrom, K.; Schallert, T.; Chang, J.Y. High-frequency stimulation of the subthalamic nucleus reverses limb-use asymmetry in rats with unilateral 6-hydroxydopamine lesions. Brain Res. 2004, 1013, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.W.; Wachtel, S.R.; Young, D.; Kang, U.J. Biochemical and anatomical characterization of forepaw adjusting steps in rat models of Parkinson’s disease: Studies on medial forebrain bundle and striatal lesions. Neuroscience 1999, 88, 617–628. [Google Scholar] [CrossRef]
- Roedter, A.; Winkler, C.; Samii, M.; Walter, G.F.; Brandis, A.; Nikkhah, G. Comparison of unilateral and bilateral intrastriatal 6-hydroxydopamine-induced axon terminal lesions: Evidence for interhemispheric functional coupling of the two nigrostriatal pathways. J. Comp. Neurol. 2001, 432, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Meredith, G.E.; Kang, U.J. Behavioral models of Parkinsons disease in rodents: A new look at an old problem. Mov. Disord. 2006, 21, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Sugiyama, K.; Akamine, S.; Namba, H. The stepping test and its learning process in different degrees of unilateral striatal lesions by 6-hydroxydopamine in rats. Neurosci. Res. 2006, 55, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Pinna, A.; Pontis, S.; Borsini, F.; Morelli, M. Adenosine A2A receptor antagonists improve deficits in initiation of movement and sensory motor integration in the unilateral 6-hydroxydopamine rat model of Parkinson’s disease. Synapse 2007, 61, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ford-Dunn, H.L.; Hayward, G.N.; Nandi, D.; Miall, R.C.; Aziz, T.Z.; Stein, J.F. The oscillatory activity in the Parkinsonian subthalamic nucleus investigated using the macro-electrodes for deep brain stimulation. Clin. Neurophysiol. 2002, 113, 1667–1672. [Google Scholar] [CrossRef]
- Badstuebner, K.; Gimsa, U.; Weber, I.; Tuchscherer, A.; Gimsa, J. Deep Brain Stimulation of Hemiparkinsonian Rats with Unipolar and Bipolar Electrodes for up to 6 Weeks: Behavioral Testing of Freely Moving Animals. Parkinsons Dis. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinna, A.; Tronci, E.; Schintu, N.; Simola, N.; Volpini, R.; Pontis, S.; Cristalli, G.; Morelli, M. A new ethyladenine antagonist of adenosine A2A receptors: Behavioral and biochemical characterization as an antiparkinsonian drug. Neuropharmacology 2010, 58, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Mukhida, K.; Baker, K.A.; Sadi, D.; Mendez, I. Enhancement of sensorimotor behavioral recovery in hemiparkinsonian rats with intrastriatal, intranigral, and intrasubthalamic nucleus dopaminergic transplants. J. Neurosci. 2001, 21, 3521–3530. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Ahmed, R.; Sagar, R.K.; Krishana, B. Neuroprotection of the nigrostriatal dopaminergic neurons by melatonin in hemiparkinsonium rat. Indian J. Med. Res. 2006, 124, 419–426. [Google Scholar] [PubMed]
- Singh, S.; Ahmad, R.; Mathur, D.; Sagar, R.K.; Krishana, B.; Arora, R.; Sharma, R.K. Neuroprotective effect of BDNF in young and aged 6-OHDA treated rat model of Parkinson disease. Indian J. Exp. Biol. 2006, 44, 699–704. [Google Scholar] [PubMed]
- Antipova, V.; Holzmann, C.; Schmitt, O.; Wree, A.; Hawlitschka, A. Botulinum Neurotoxin A Injected Ipsilaterally or Contralaterally into the Striatum in the Rat 6-OHDA Model of Unilateral Parkinson’s Disease Differently Affects Behavior. Front. Behav. Neurosci. 2017, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Wree, A.; Mix, E.; Hawlitschka, A.; Antipova, V.; Witt, M.; Schmitt, O.; Benecke, R. Intrastriatal botulinum toxin abolishes pathologic rotational behaviour and induces axonal varicosities in the 6-OHDA rat model of Parkinson’s disease. Neurobiol. Dis. 2011, 41, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Antipova, V.; Hawlitschka, A.; Mix, E.; Schmitt, O.; Dräger, D.; Benecke, R.; Wree, A. Behavioral and structural effects of unilateral intrastriatal injections of botulinum neurotoxin a in the rat model of Parkinson’s disease. J. Neurosci. Res. 2013, 91, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, C.; Dräger, D.; Mix, E.; Hawlitschka, A.; Antipova, V.; Benecke, R.; Wree, A. Effects of intrastriatal botulinum neurotoxin A on the behavior of Wistar rats. Behav. Brain Res. 2012, 234, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Mehlan, J.; Brosig, H.; Schmitt, O.; Mix, E.; Wree, A.; Hawlitschka, A. Intrastriatal injection of botulinum neurotoxin-A is not cytotoxic in rat brain—A histological and stereological analysis. Brain Res. 2016, 1630, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Wree, A. Experimental intrastriatal applications of botulinum neurotoxin-A: A review. Int. J. Mol. Sci. 2018, 19, 1392. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Antipova, V.; Schmitt, O.; Witt, M.; Benecke, R.; Mix, E.; Wree, A. Intracerebrally applied botulinum neurotoxin in experimental neuroscience. Curr. Pharm. Biotechnol. 2013, 14, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Holzmann, C.; Witt, S.; Spiewok, J.; Neumann, A.M.; Schmitt, O.; Wree, A.; Antipova, V. Intrastriatally injected botulinum neurotoxin-A differently effects cholinergic and dopaminergic fibers in C57BL/6 mice. Brain Res. 2017, 1676, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.Á.; Rodríguez-Oroz, M.C.; Benitez-Temino, B.; Blesa, F.J.; Guridi, J.; Marin, C.; Rodriguez, M. Functional organization of the basal ganglia: Therapeutic implications for Parkinson’s disease. Mov. Disord. 2008, 23, S548–S559. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.Á.; Marin, C.; Rodriguez-Oroz, C.; Blesa, J.; Benitez-Temiño, B.; Mena-Segovia, J.; Rodríguez, M.; Olanow, C.W. The basal ganglia in Parkinson’s disease: Current concepts and unexplained observations. Ann. Neurol. 2008, 64, S30–S46. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.E.; O’Malley, K. Axon degeneration in Parkinson’s disease. Exp. Neurol. 2013, 246, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K. Neuropathological Staging of Brain Pathology in Sporadic Parkinson’s disease: Separating the Wheat from the Chaff. J. Parkinsons Dis. 2017, 7, S73–S87. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.-Y.Y.; McIver, E.L.; Kovaleski, R.F.; Atherton, J.F.; Bevan, M.D. Loss of Hyperdirect Pathway Cortico-Subthalamic Inputs Following Degeneration of Midbrain Dopamine Neurons. Neuron 2017, 95, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Pisani, A.; Bernardi, G.; Ding, J.; Surmeier, D.J. Re-emergence of striatal cholinergic interneurons in movement disorders. Trends Neurosci. 2007, 30, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Benecke, R.; Dressler, D. Botulinum toxin treatment of axial and cervical dystonia. Disabil. Rehabil. 2007, 29, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Erickson, B.P.; Lee, W.W.; Cohen, J.; Grunebaum, L.D. The role of neurotoxins in the periorbital and midfacial areas. Facial Plast. Surg. Clin. N. Am. 2015, 23, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Santamato, A.; Micello, M.F.; Ranieri, M.; Valeno, G.; Albano, A.; Baricich, A.; Cisari, C.; Intiso, D.; Pilotto, A.; Logroscino, G.; et al. Employment of higher doses of botulinum toxin type A to reduce spasticity after stroke. J. Neurol. Sci. 2015, 350, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Becher, K.F.; Sieber, C.C. Urinary incontinence: what’s new in therapy? Dtsch. Med. Wochenschr. 2015, 140, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Alshadwi, A.; Nadershah, M.; Osborn, T. Therapeutic applications of botulinum neurotoxins in head and neck disorders. Saudi Dent. J. 2015, 27, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellman, A.; Torres Russotto, D. Botulinum toxin in the management of blepharospasm: Current evidence and recent developments. Ther. Adv. Neurol. Disord. 2015, 8, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Holzmann, C.; Wree, A.; Antipova, V.; Hawlitschka, A.; Holzmann, C.; Wree, A.; Antipova, V. Repeated Intrastriatal Botulinum Neurotoxin-A Injection in Hemiparkinsonian Rats Increased the Beneficial Effect on Rotational Behavior. Toxins 2018, 10, 368. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Fukumoto, T.; Takatori, K.; Nagino, K.; Hiraoka, K. Abnormalities of the first three steps of gait initiation in patients with Parkinson’s disease with freezing of gait. Parkinsons Dis. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Chu, H.; Zhang, Y.; Wang, X. Deep brain stimulation to alleviate freezing of gait and cognitive dysfunction in Parkinson’s disease: Update on current research and future perspectives. Front. Neurosci. 2018, 12, 29. [Google Scholar] [CrossRef] [PubMed]
- Walton, C.C.; Shine, J.M.; Hall, J.M.; O’Callaghan, C.; Mowszowski, L.; Gilat, M.; Szeto, J.Y.Y.; Naismith, S.L.; Lewis, S.J.G. The major impact of freezing of gait on quality of life in Parkinson’s disease. J. Neurol. 2015, 262, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Curtze, C.; Nutt, J.G.; Carlson-Kuhta, P.; Mancini, M.; Horak, F.B. Levodopa Is a Double-Edged Sword for Balance and Gait in People With Parkinson’s Disease. Mov. Disord. 2015, 30, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Moore, O.; Peretz, C.; Giladi, N. Freezing of gait affects quality of life of peoples with Parkinson’s disease beyond its relationships with mobility and gait. Mov. Disord. 2007, 22, 2192–2195. [Google Scholar] [CrossRef] [PubMed]
- Antala, B.; Bhuva, S.; Gupta, S.; Lahkar, M.; Patel, M.; Rabadiya, S. Protective effect of methanolic extract of Garcinia indica fruits in 6-OHDA rat model of Parkinson′s disease. Indian J. Pharmacol. 2012, 44, 683. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gui, X.H.; Huang, L.P.; Deng, M.Z.; Fang, R.M.; Ke, X.H.; He, Y.P.; Li, L.; Fang, Y.Q. Neuroprotective Effects of β-Asarone Against 6-Hydroxy Dopamine-Induced Parkinsonism via JNK/Bcl-2/Beclin-1 Pathway. Mol. Neurobiol. 2016, 53, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Sugiyama, K.; Fang, X.; Yamaguchi, H.; Akamine, S.; Magata, Y.; Namba, H. Different striatal D2-like receptor function in an early stage after unilateral striatal lesion and medial forebrain bundle lesion in rats. Brain Res. 2010, 1317, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Universities Federation of Animal Federation for Animal Welfare: London, UK, 1959. [Google Scholar]
- Hahn, M.; Timmer, M.; Nikkhah, G. Survival and early functional integration of dopaminergic progenitor cells following transplantation in a rat model of Parkinson’s disease. J. Neurosci. Res. 2009, 87, 2006–2019. [Google Scholar] [CrossRef] [PubMed]
- Frick, K.M.; Kim, J.J.; Baxter, M.G. Effects of complete immunotoxin lesions of the cholinergic basal forebrain on fear conditioning and spatial learning. Hippocampus 2004, 14, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Janisiewicz, A.M.; Jackson, O.; Firoz, E.F.; Baxter, M.G. Environment-spatial conditional learning in rats with selective lesions of medial septal cholinergic neurons. Hippocampus 2004, 14, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Sabol, K.E.; Neill, D.B.; Wages, S.A.; Church, W.H.; Justice, J.B. Dopamine depletion in a striatal subregion disrupts performance of a skilled motor task in the rat. Brain Res. 1985, 335, 33–43. [Google Scholar] [CrossRef]
- Fairley, P.C.; Marshall, J.F. Dopamine in the lateral caudate-putamen of the rat is essential for somatosensory orientation. Behav. Neurosci. 1986, 100, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Salamone, J.D.; Kurth, P.A.; McCullough, L.D.; Sokolowski, J.D.; Cousins, M.S. The role of brain dopamine in response initiation: Effects of haloperidol and regionally specific dopamine depletions on the local rate of instrumental responding. Brain Res. 1993, 628, 218–226. [Google Scholar] [CrossRef]
- Sun, W.; Sugiyama, K.; Asakawa, T.; Ito-Yamashita, T.; Namba, H. Behavioral performance at early (4 weeks) and later (6 months) stages in rats with unilateral medial forebrain bundle and striatal 6-hydroxydopamine lesions. Neurol. Med. Chir. 2013, 53, 7–11. [Google Scholar] [CrossRef]
- Finger, S.; Almli, C.R. Brain damage and neuroplasticity: Mechanisms of recovery or development? Brain Res. Rev. 1985, 10, 177–186. [Google Scholar] [CrossRef]
- Marshall, J.F. Neural Plasticity and Recovery of Function After Brain Injury. Int. Rev. Neurobiol. 1985, 26, 201–247. [Google Scholar] [CrossRef] [PubMed]
- Finger, S. Recovery from Brain Damage: Research and Theory; Springer US: Boston, MA, USA, 1978; ISBN 9781461577232. [Google Scholar]
- Brain Plasticity, Learning, and Memory; Will, B.E.; Schmitt, P.; Dalrymple-Alford, J.C. (Eds.) Advances in Behavioral Biology; Springer US: Boston, MA, USA, 1985; Volume 28, ISBN 978-1-4684-5005-7. [Google Scholar]
- Schulkin, J. Preoperative Events: Their Effects on Behavior Following Brain Damage; Taylor and Francis: Abingdon, UK, 1989; ISBN 0805800212r0805805354. [Google Scholar]
- Rosenzweig, M.R.; Bennett, E.L. Psychobiology of plasticity: Effects of training and experience on brain and behavior. Behav. Brain Res. 1996, 78, 57–65. [Google Scholar] [CrossRef]
- Bach-y-Rita, P.; International Rehabilitation Medicine Association; Smith-Kettlewell Eye Research Foundation. Recovery of Function: Theoretical Considerations for Brain Injury Rehabilitation; Bach-y-Rita, P., Ed.; Hyuber: Berne, Switzerland, 1988; ISBN 0920887163. [Google Scholar]
- Barbeau, H.; Chau, C.; Rossignol, S. Noradrenergic agonists and locomotor training affect locomotor recovery after cord transection in adult cats. Brain Res. Bull. 1993, 30, 387–393. [Google Scholar] [CrossRef]
- Handelmann, G.E.; Olton, D.S. Spatial memory following damage to hippocampal CA3 pyramidal cells with kainic acid: Impairment and recovery with preoperative training. Brain Res. 1981, 217, 41–58. [Google Scholar] [CrossRef]
- Nudo, R.J.; Wise, B.M.; SiFuentes, F.; Milliken, G.W. Neural substrates for the effects of rehabilitative training on motor recovery after ischemic infarct. Science 1996, 272, 1791–1794. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.; Brown, V.J.; Dunnett, S.B.; Robbins, T.W. Striatal Graft-Associated Recovery of a Lesion-Induced Performance Deficit in the Rat Requires Learning to Use The Transplant. Eur. J. Neurosci. 1992, 4, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Abrous, D.N.; Rodriguez, J.J.; Montaron, M.F.; Aurousseau, C.; Le Moal, M.; Barneoud, P. Behavioural recovery after unilateral lesion of the dopaminergic mesotelencephalic pathway: Effect of repeated testing. Neuroscience 1998, 84, 213–221. [Google Scholar] [CrossRef]
- Antipova, V.; Wree, A.; Holzmann, C.; Mann, T.; Palomero-Gallagher, N.; Zilles, K.; Schmitt, O.; Hawlitschka, A. Unilateral Botulinum Neurotoxin-A Injection into the Striatum of C57BL/6 Mice Leads to a Different Motor Behavior Compared with Rats. Toxins 2018, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Metz, G.A.; Whishaw, I.Q. Drug-induced rotation intensity in unilateral dopamine-depleted rats is not correlated with end point or qualitative measures of forelimb or hindlimb motor performance. Neuroscience 2002, 111, 325–336. [Google Scholar] [CrossRef]
- Tseng, K.Y.; Kargieman, L.; Gacio, S.; Riquelme, L.A.; Murer, M.G. Consequences of partial and severe dopaminergic lesion on basal ganglia oscillatory activity and akinesia. Eur. J. Neurosci. 2005, 22, 2579–2586. [Google Scholar] [CrossRef] [PubMed]
- Barnéoud, P.; Descombris, E.; Aubin, N.; Abrous, D.N. Evaluation of simple and complex sensorimotor behaviours in rats with a partial lesion of the dopaminergic nigrostriatal system. Eur. J. Neurosci. 2000, 12, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.; Sauer, H.; Lee, C.S.; Bjorklund, A. Short-Term GDNF Treatment Provides Long-Term Rescue of Lesioned Nigral Dopaminergic Neurons in a Rat Model of Parkinson’s Disease. J. Neurosci. 1996, 16, 7206–7215. [Google Scholar] [CrossRef] [PubMed]
- Kirik, D.; Winkler, C.; Björklund, A. Growth and functional efficacy of intrastriatal nigral transplants depend on the extent of nigrostriatal degeneration. J. Neurosci. 2001, 21, 2889–2896. [Google Scholar] [CrossRef] [PubMed]
- Kirik, D.; Rosenblad, C.; Bjorklund, A. Preservation of a functional nigrostriatal dopamine pathway by GDNF in the intrastriatal 6-OHDA lesion model depends on the site of administration of the trophic factor. Eur. J. Neurosci. 2000, 12, 3871–3882. [Google Scholar] [CrossRef] [PubMed]
- Grealish, S.; Mattsson, B.; Draxler, P.; Björklund, A. Characterisation of behavioural and neurodegenerative changes induced by intranigral 6-hydroxydopamine lesions in a mouse model of Parkinson’s disease. Eur. J. Neurosci. 2010, 31, 2266–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikkhah, G.; Falkenstein, G.; Rosenthal, C. Restorative plasticity of dopamine neuronal transplants depends on the degree of hemispheric dominance. J. Neurosci. 2001, 21, 6252–6263. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Sugiyama, K.; Akamine, S.; Sun, W.; Namba, H. The different performance among motor tasks during the increasing current intensity of deep brain stimulation of the subthalamic nucleus in rats with different degrees of the unilateral striatal lesion. Neurosci. Lett. 2010, 480, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Manfredsson, F.P.; Burger, C.; Sullivan, L.F.; Muzyczka, N.; Lewin, A.S.; Mandel, R.J. rAAV-mediated nigral human parkin over-expression partially ameliorates motor deficits via enhanced dopamine neurotransmission in a rat model of Parkinson’s disease. Exp. Neurol. 2007, 207, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Köllensperger, M.; Stefanova, N.; Reindl, M.; Poewe, W.; Wenning, G.K. Loss of dopaminergic responsiveness in a double lesion rat model of the Parkinson variant of multiple system atrophy. Mov. Disord. 2007, 22, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Lettfuss, N.Y.; Fischer, K.; Sossi, V.; Pichler, B.J.; von Ameln-Mayerhofer, A. Imaging DA release in a rat model of L-DOPA-induced dyskinesias: A longitudinal in vivo PET investigation of the antidyskinetic effect of MDMA. Neuroimage 2012, 63, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Seeger-Armbruster, S.; Von Ameln-Mayerhofer, A. Short- and long-term unilateral 6-hydroxydopamine lesions in rats show different changes in characteristics of spontaneous firing of substantia nigra pars reticulata neurons. Exp. Brain Res. 2013, 224, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; London, UK, 2015; ISBN 9780123919496. [Google Scholar]
- Schallert, T.; De Ryck, M.; Whishaw, I.Q.; Ramirez, V.D.; Teitelbaum, P. Excessive bracing reactions and their control by atropine and L-DOPA in an animal analog of Parkinsonism. Exp. Neurol. 1979, 64, 33–43. [Google Scholar] [CrossRef]
- VLC: Offizielle Webseite—Freie Multimedialösungen für alle Betriebssysteme!—VideoLAN. Available online: https://www.videolan.org/ (accessed on 28 January 2019).







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antipova, V.; Holzmann, C.; Hawlitschka, A.; Wree, A. Botulinum Neurotoxin-A Injected Intrastriatally into Hemiparkinsonian Rats Improves the Initiation Time for Left and Right Forelimbs in Both Forehand and Backhand Directions. Int. J. Mol. Sci. 2019, 20, 992. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040992
Antipova V, Holzmann C, Hawlitschka A, Wree A. Botulinum Neurotoxin-A Injected Intrastriatally into Hemiparkinsonian Rats Improves the Initiation Time for Left and Right Forelimbs in Both Forehand and Backhand Directions. International Journal of Molecular Sciences. 2019; 20(4):992. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040992
Chicago/Turabian StyleAntipova, Veronica, Carsten Holzmann, Alexander Hawlitschka, and Andreas Wree. 2019. "Botulinum Neurotoxin-A Injected Intrastriatally into Hemiparkinsonian Rats Improves the Initiation Time for Left and Right Forelimbs in Both Forehand and Backhand Directions" International Journal of Molecular Sciences 20, no. 4: 992. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040992