Potential of Novel Sequence Type of Burkholderia cenocepacia for Biological Control of Root Rot of Maize (Zea mays L.) Caused by Fusarium temperatum

Abstract
:1. Introduction
2. Results and Discussion
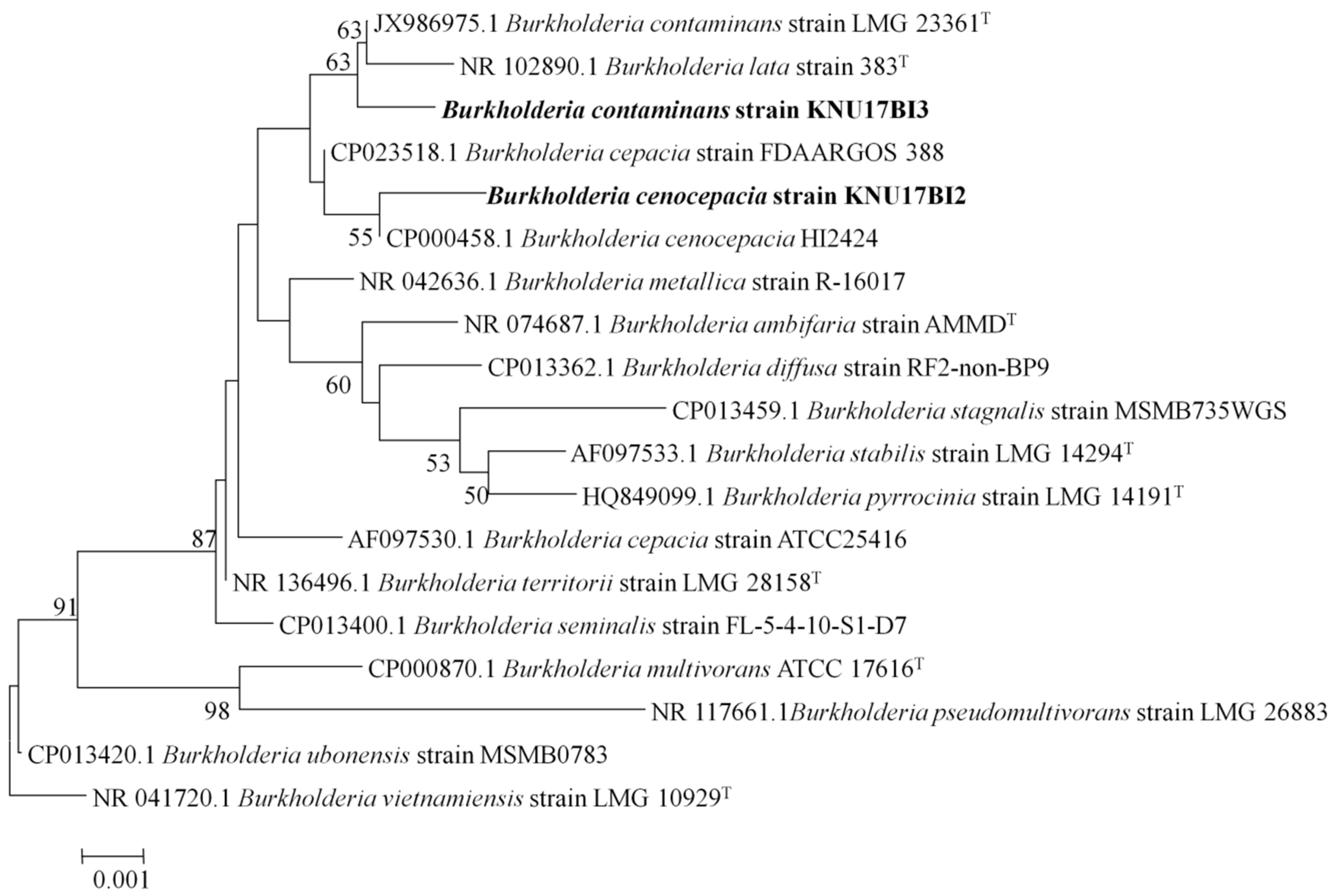
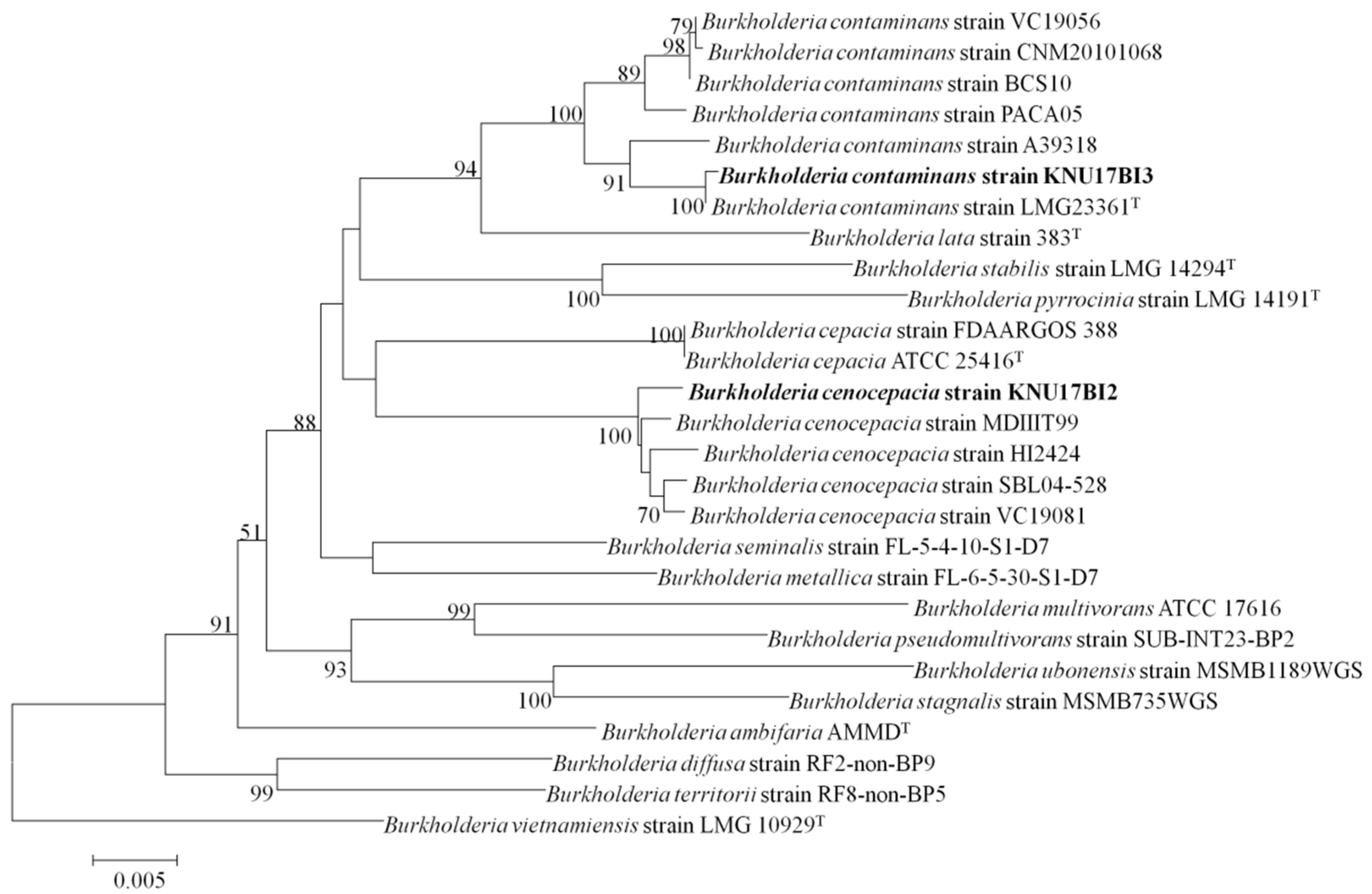
2.1. Identification of Bacterial Isolates
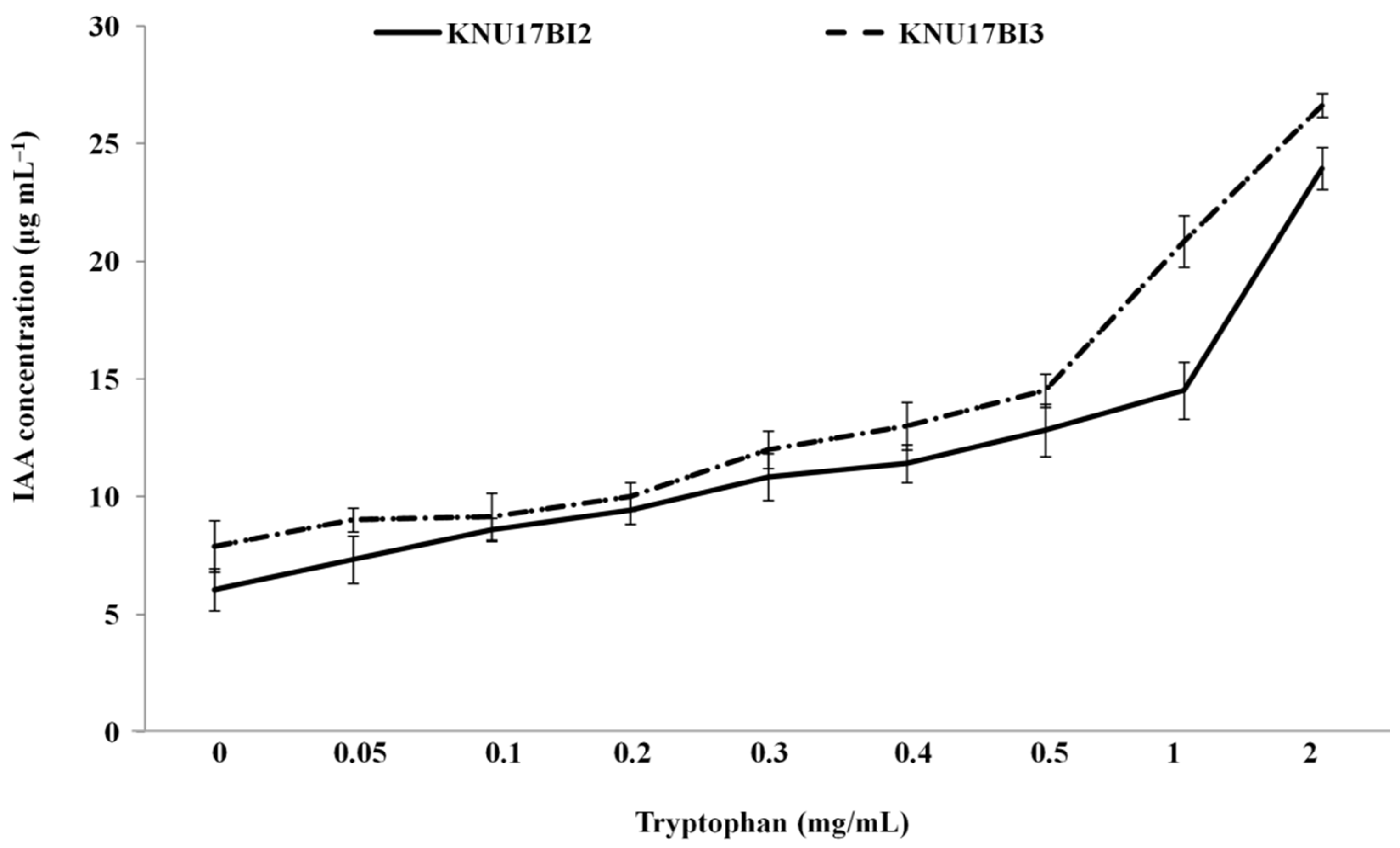
2.2. In Vitro Plant Growth Promoting (PGP) Activity Assays
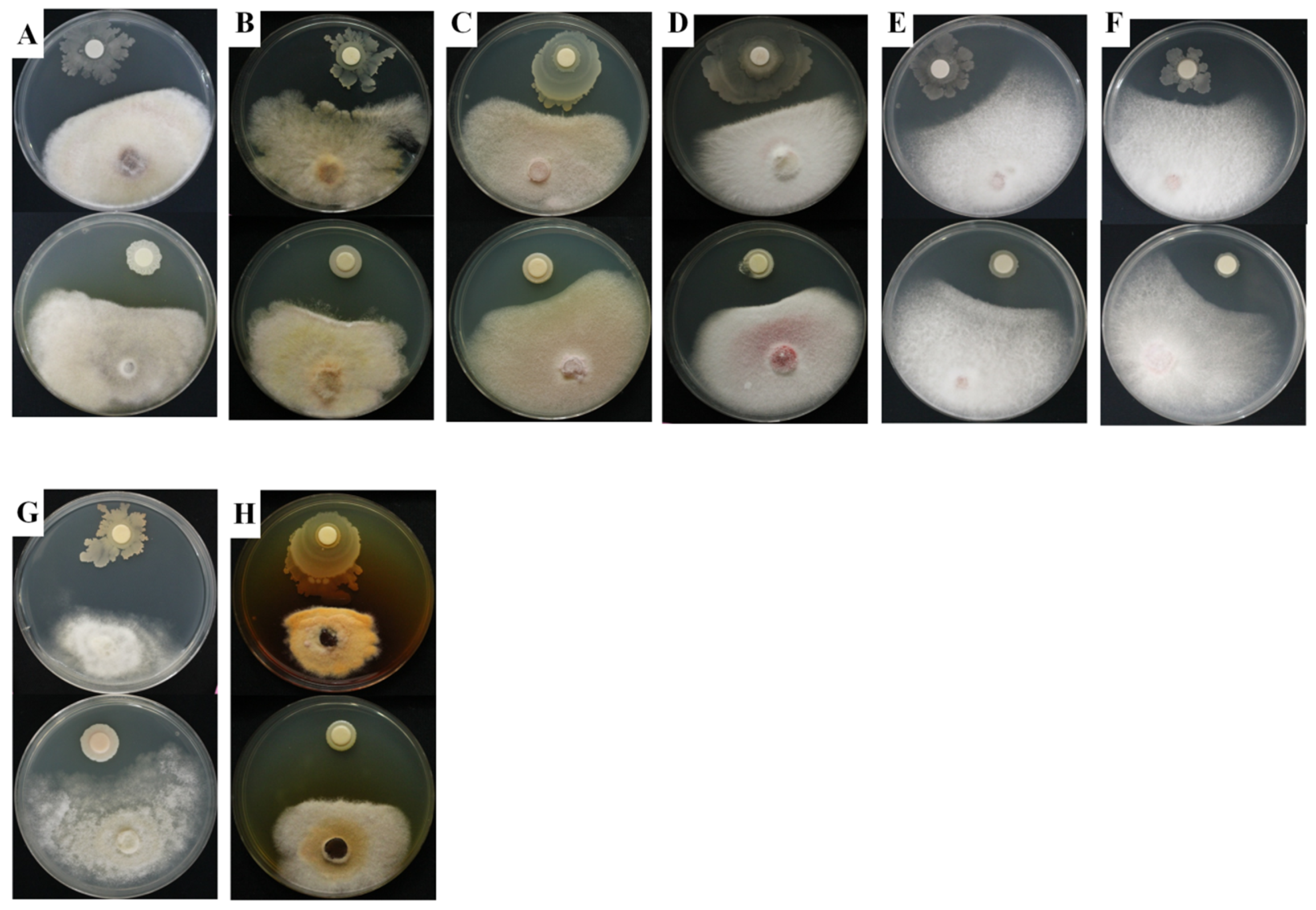
2.3. In Vitro Antifungal Activity
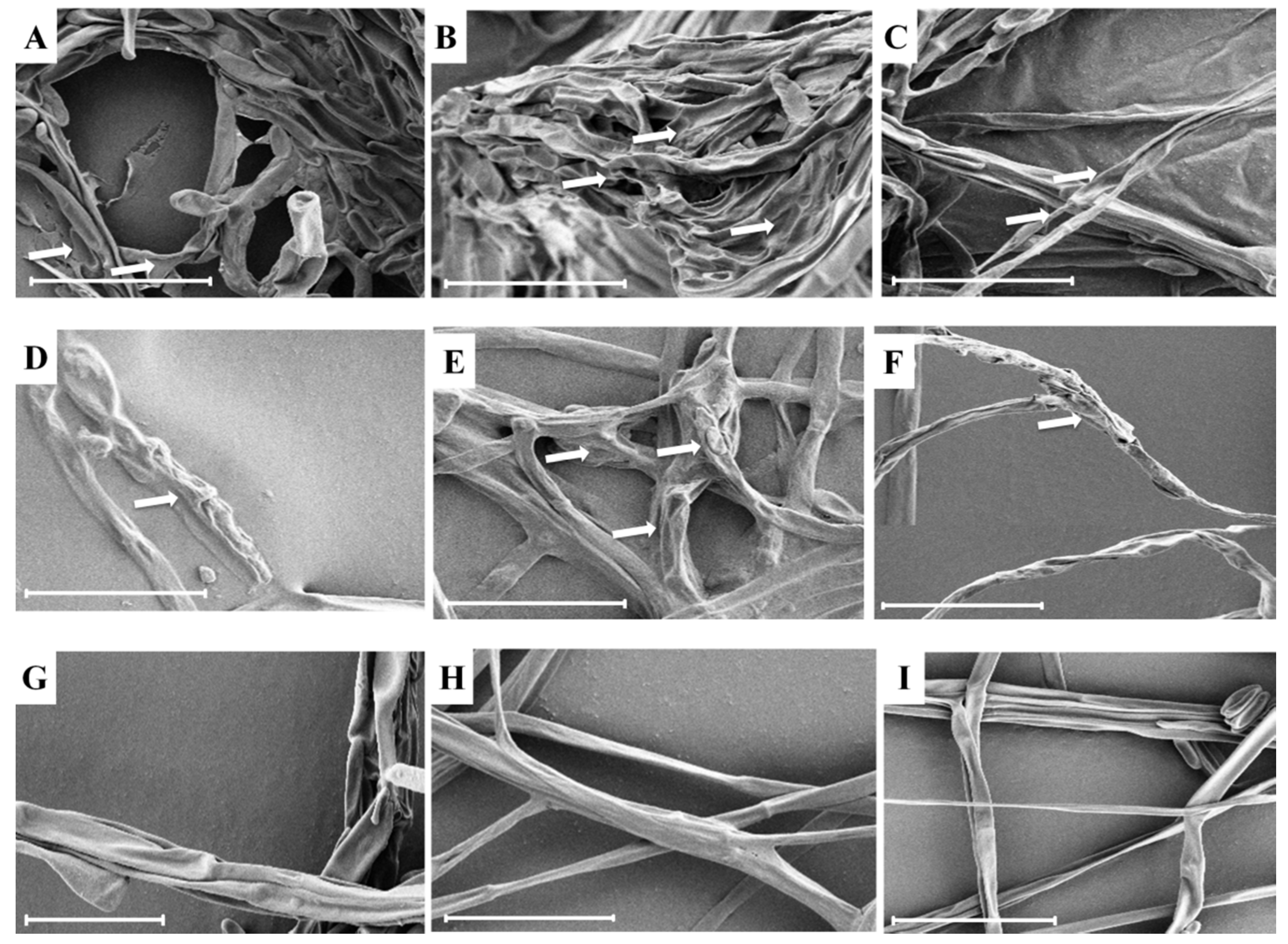
2.4. SEM Analysis
2.5. Greenhouse Experiments
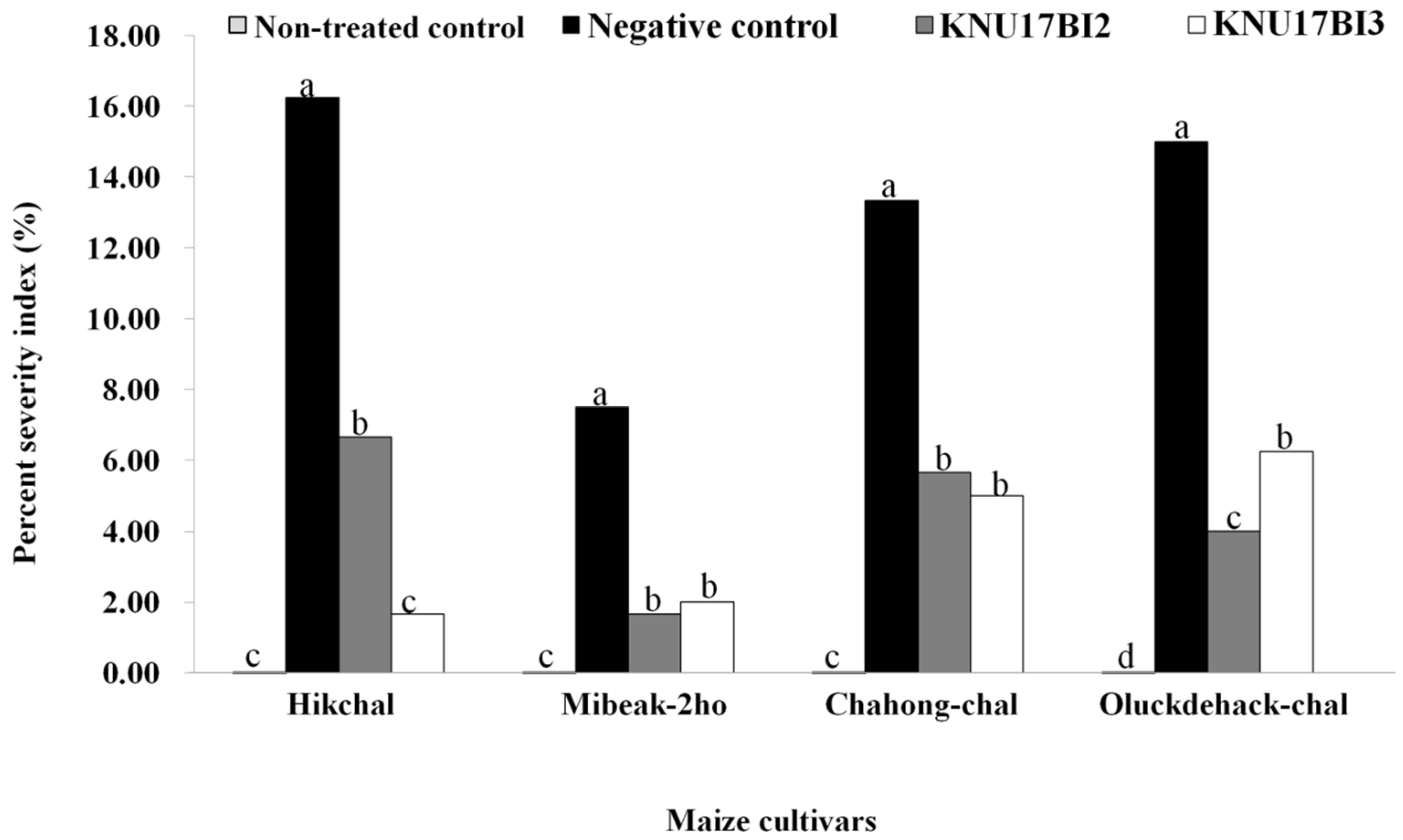
2.5.1. Effect of Strain KNU17BI2 and Strain KNU17BI3 on Virulence of F. temperatum
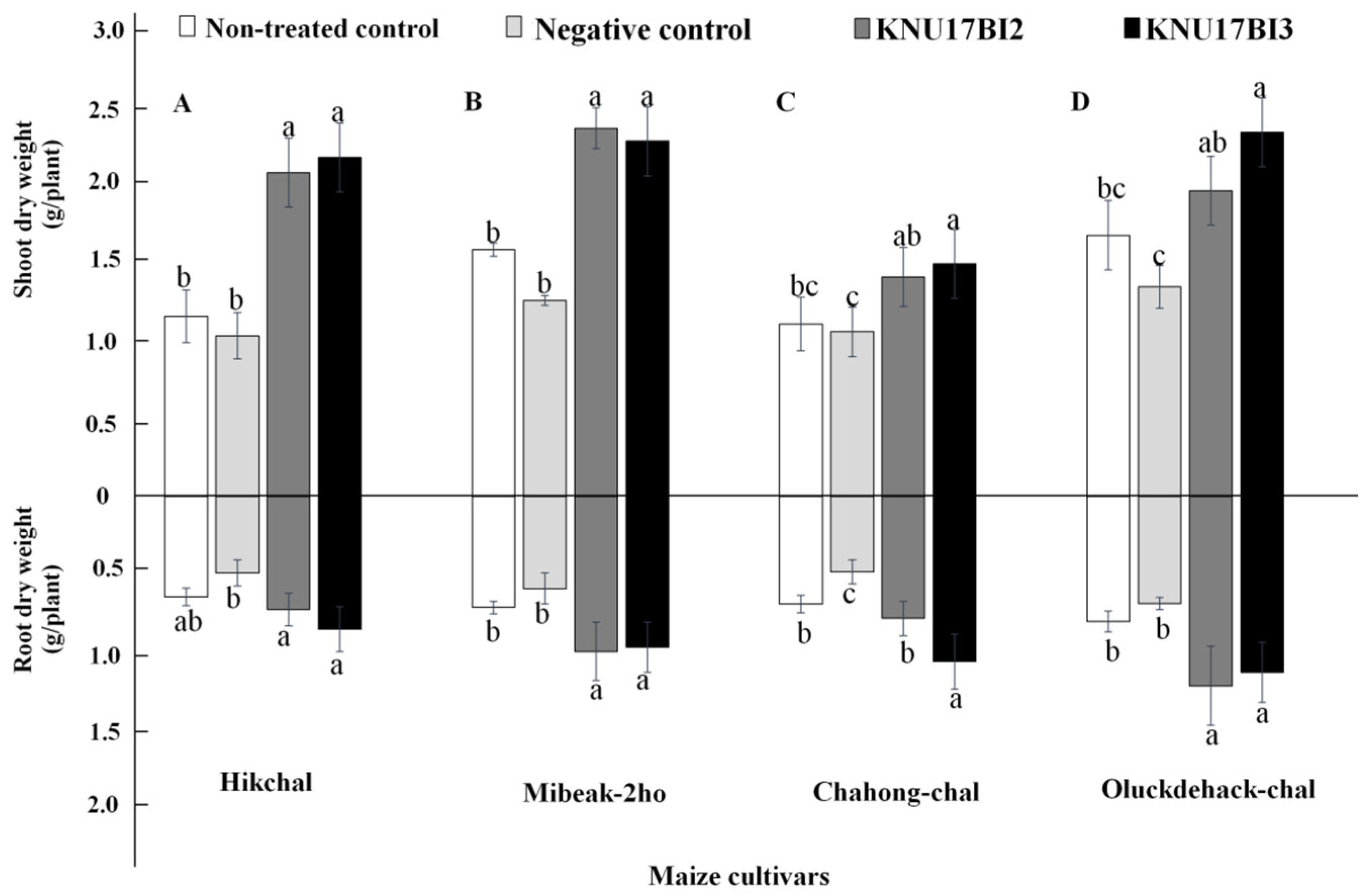
2.5.2. Plant Growth Promotion Effect of Strain KNU17BI2 and Strain KNU17BI3
3. Materials and Methods
3.1. Microbial Sources
3.2. Identification of Bacterial Isolates
16S rRNA Gene Sequencing
3.3. Multilocus Sequencing Analysis and Typing (MLSA-MLST)
3.4. Genbank Accession Numbers
3.5. In Vitro Antifungal Activity
3.6. Scanning Electron Microscope (SEM) Analysis
3.7. In Vitro Plant Growth Promoting (PGP) Activity Assays
3.8. Greenhouse Experiments
3.9. Assessment of Effect of Burkholderia Strains on Virulence of F. temperatum
3.10. Statistical Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Varela, C.P.; Casal, O.A.; Padin, M.C.; Martinez, V.F.; Oses, M.S.; Scauflaire, J.; Munaut, F.; Castro, M.B.; Vázquez, J.M. First report of Fusarium temperatum causing seedling blight and stalk rot on maize in Spain. Plant Dis. 2013, 97, 1252. [Google Scholar] [CrossRef] [PubMed]
- Czembor, E.; Stępień, Ł.; Waśkiewicz, A. Fusarium temperatum as a new species causing ear rot on maize in Poland. Plant Dis. 2014, 98, 1001. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Han, J.H.; Lee, J.K.; Kim, K.S. Characterization of the maize stalk rot pathogens Fusarium subglutinans and F. temperatum and the effect of fungicides on their mycelial growth and colony formation. Plant Pathol. J. 2014, 30, 397. [Google Scholar] [PubMed]
- Lanza, F.E.; Mayfield, D.A.; Munkvold, G.P. First report of Fusarium temperatum causing maize seedling blight and seed rot in North America. Plant Dis. 2016, 100, 1019. [Google Scholar] [CrossRef]
- Wang, J.H.; Zhang, J.B.; Li, H.P.; Gong, A.D.; Xue, S.; Agboola, R.S.; Liao, Y.C. Molecular identification, mycotoxin production and comparative pathogenicity of Fusarium temperatum isolated from maize in China. J. Phytopathol. 2014, 162, 147–157. [Google Scholar] [CrossRef]
- Reid, T.C.; Hausbeck, M.K.; Kizilkaya, K. Use of fungicides and biological controls in the suppression of Fusarium crown and root rot of asparagus under greenhouse and growth chamber conditions. Plant Dis. 2002, 86, 493–498. [Google Scholar] [CrossRef]
- Figueroa-López, A.M.; Cordero-Ramírez, J.D.; Martínez-Álvarez, J.C.; López-Meyer, M.; Lizárraga-Sánchez, G.J.; Félix-Gastélum, R.; Castro-Martínez, C.; Maldonado-Mendoza, I.E. Rhizospheric bacteria of maize with potential for biocontrol of Fusarium verticillioides. SpringerPlus 2016, 5, 330. [Google Scholar] [CrossRef] [PubMed]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbial. 2016, 29, 1538. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Kim, Y.E.; Park, S.J. Burkholderia alba sp. nov., isolated from a soil sample on Halla mountain in Jeju island. J. Microbiol. 2018, 56, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Gonçalves, L.C.; Gorlach-Lira, K. Lipases and biosurfactants production by the newly isolated Burkholderia sp. Braz. J. Biol. Sci. 2018, 30, 57–68. [Google Scholar] [CrossRef]
- Tagele, S.B.; Kim, S.W.; Lee, H.G.; Kim, H.S.; Lee, Y.S. Effectiveness of multi-trait Burkholderia contaminans KNU17BI1 in growth promotion and management of banded leaf and sheath blight in maize seedling. Microbiol. Res. 2018, 214, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Penttinen, P.; Zhang, X.; Ao, X.; Liu, M.; Yu, X.; Chen, Q. Maize rhizosphere in Sichuan, China, hosts plant growth promoting Burkholderia cepacia with phosphate solubilizing and antifungal abilities. Microbiol. Res. 2014, 169, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Esmaeel, Q.; Pupin, M.; Jacques, P.; Leclère, V. Nonribosomal peptides and polyketides of Burkholderia: New compounds potentially implicated in biocontrol and pharmaceuticals. Environ. Sci. Pollut. Res. 2018, 25, 29794–29807. [Google Scholar] [CrossRef] [PubMed]
- Kandel, S.L.; Firrincieli, A.; Joubert, P.M.; Okubara, P.A.; Leston, N.D.; McGeorge, K.M. An in vitro study of bio-control and plant growth promotion potential of Salicaceae endophytes. Front. Microbiol. 2017, 8, 386. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, P.; Dawyndt, P. Classification and identification of the Burkholderia cepacia complex: Past, present and future. Syst. Appl. Microbiol. 2011, 34, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Yabuuchi, E.; Kosako, Y.; Oyaizu, H.; Yano, I.; Hotta, H.; Hashimoto, Y.; Ezaki, T.; Arakawa, M. Proposal of Burkholderia gen. nov. and transfer of seven species of genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes, 1981) comb. nov. Microbiol. Immunol. 1992, 36, 1251–1275. [Google Scholar] [PubMed]
- Estrada-de los Santos, P.; Palmer, M.; Chávez-Ramírez, B.; Beukes, C.; Steenkamp, E.; Briscoe, L. Whole Genome Analyses Suggests that Burkholderia sensu lato Contains Two Additional Novel Genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): Implications for the Evolution of Diazotrophy and Nodulation in the Burkholderiaceae. Genes 2018, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Vandamme, P.; Govan, J.R.; LiPuma, J.J. Taxonomy and identification of the Burkholderia cepacia complex. J. Clin. Microbiol. 2001, 39, 3427–3436. [Google Scholar] [CrossRef] [PubMed]
- Spilker, T.; Baldwin, A.; Bumford, A.; Dowson, C.G.; Mahenthiralingam, E.; LiPuma, J.J. Expanded multilocus sequence typing for Burkholderia species. J. Clin. Microbiol. 2009, 47, 2607–2610. [Google Scholar] [CrossRef] [PubMed]
- Fila, L.; Dřevínek, P. Burkholderia cepacia complex in cystic fibrosis in the post-epidemic period: Multilocus sequence typing-based approach. Folia Microbiol. 2017, 62, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Yoder-Himes, D.R.; Chain, P.S.G.; Zhu, Y.; Wurtzel, O.; Rubin, E.M.; Tiedje, J.M.; Sorek, R. Mapping the Burkholderia cenocepacia niche response via high-throughput sequencing. Proc. Natl. Acad. Sci. USA 2009, 106, 3976–3981. [Google Scholar] [CrossRef] [PubMed]
- Goenadi, D.H.; Sisweto, I.; Sugiarto, Y. Bioactivation of poorly soluble phosphate rocks with a phosphorus- solubilizing fungus. Soil Sci. Soc. Am. J. 2000, 64, 927–932. [Google Scholar]
- Mahamuni, S.V. Anticeratocystis paradoxa and Alternaria alternata attribute of Burkholderia cenocepacia strain vimp 01 (jq867371). Int. J. Bioassays 2015, 4, 4290–4295. [Google Scholar]
- Otieno, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Hameed, S.; Tariq, M.; Zafar, M.; Ali, A.; Ahmad, N. Characterization of mineral phosphate-solubilizing bacteria for enhanced sunflower growth and yield-attributing traits. Ann. Microbiol. 2015, 65, 1525–1536. [Google Scholar] [CrossRef]
- Taurian, T.; Anzuay, M.S.; Angelini, J.G.; Tonelli, M.L.; Ludueña, L.; Pena, D.; Ibáñez, F.; Fabra, A. Phosphate-solubilizing peanut associated bacteria: Screening for plant growth-promoting activities. Plant Soil 2010, 329, 421–431. [Google Scholar] [CrossRef]
- Dinesha, R.; Srinivasana, V.; Hamza, S.; Sarathambal, C.; Ankegowda, S.J.; Ganeshamurthy, A.N.; Gupta, S.B.; Nair, V.A.; Subila, K.P.; Lijina, A.; Divya, V.C. Isolation and characterization of potential Zn solubilizing bacteria from soil and its effects on soil Zn release rates, soil available Zn and plant Zn content. Geoderma 2018, 321, 173–186. [Google Scholar] [CrossRef]
- Cozzi, D.; Desidevi, P.G.; Lepri, L. The mechanism of ion exchange with algenic acid. J. Chromatogr. 1969, 40, 130–135. [Google Scholar] [CrossRef]
- Vaid, S.K.; Kumar, B.; Sharma, A.; Shukla, A.K.; Srivastava, P.C. Effect of Zn solubilizing bacteria on growth promotion and Zn nutrition of rice. J. Soil Sci. Plant Nutr. 2014, 14, 889–910. [Google Scholar] [CrossRef]
- Johnson-Beebout, S.E.; Lauren, J.G.; Duxbury, J.M. Immobilization of zinc fertilizer in flooded soils monitored by adapted DTPA soil test. Commun. Soil Sci. Plant Anal. 2009, 40, 1842–1861. [Google Scholar] [CrossRef]
- Natheer, S.E.; Muthukkaruppan, S. Assessing the in vitro zinc solubilization potential and improving sugarcane growth by inoculating Gluconacetobacter diazotrophicus. Ann. Microbiol. 2012, 62, 435–441. [Google Scholar] [CrossRef]
- Ong, K.S.; Lee, L.H.; Yule, C.M.; Cheow, Y.L.; Lee, S.M. Burkholderia paludis sp. nov. an antibiotic-siderophore producing novel Burkholderia cepacia Complex Species, isolated from Malaysian tropical peat swamp soil. Front. Microbiol. 2016, 7, 20–46. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Wang, X.; Baird, S.M.; Showmaker, K.C.; Smith, L.; Peterson, D.G.; Lu, S. Comparative genome-wide analysis reveals that Burkholderia contaminans MS14 possesses multiple antimicrobial biosynthesis genes but not major genetic loci required for pathogenesis. MicrobiologyOpen 2016, 5, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Hrynkiewicz, K.; Baum, C.; Leinweber, P. Density, metabolic activity and identity of cultivable rhizosphere bacteria on Salix viminalis in disturbed arable and landfill soils. J. Plant Nutr. Soil Sci. 2010, 173, 747–756. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.L.; Gilani, S.A.; Waqas, M.; Al-Hosni, K.; Al-Khiziri, S.; Kim, Y.H. Endophytes from medicinal plants and their potential for producing indole acetic acid, improving seed germination and mitigating oxidative stress. J. Zhejiang Univ.-SC B 2017, 18, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collavino, M.M.; Sansberro, P.A.; Mroginski, L.A.; Aguilar, O.M. Comparison of in vitro solubilization activity of diverse phosphate-solubilizing bacteria native to acid soil and their ability to promote Phaseolus vulgaris growth. Biol. Fertil. Soils 2010, 46, 727–738. [Google Scholar] [CrossRef]
- Onofre-Lemus, J.; Hernandez-Lucas, I.; Girard, L.; Caballero-Mellado, J. ACC (1-Aminocyclopropane-1-Carboxylate) deaminase activity, a widespread trait in Burkholderia species, and its growth-promoting effect on tomato plants. Appl. Environ. Microbiol. 2009, 75, 6581–6590. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: A weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [PubMed]
- Karthik, C.; Elangovan, N.; Kumar, T.S.; Govindharaju, S.; Barathi, S.; Oves, M.; Arulselvi, P.I. Characterization of multifarious plant growth promoting traits of rhizobacterial strain AR6 under Chromium (VI) stress. Microbiol. Res. 2017, 204, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.P.; Xu, J.G.; Song, P.; Song, J.N.; Chen, W.L. Isolation and identification of a potential biocontrol agent Bacillus subtilis QM3 from Qinghai yak dung in China. World J. Microbiol. Biotechnol. 2008, 24, 2451–2458. [Google Scholar] [CrossRef]
- Ncube, E.; Flett, B.C.; Waalwijk, C.; Viljoen, A. Fusarium spp. and levels of fumonisins in maize produced by subsistence farmers in South Africa. S. Afr. J. Sci. 2011, 107, 1–7. [Google Scholar] [CrossRef]
- Matarese, F.; Sarrocco, S.; Gruber, S.; Seidl-Seiboth, V.; Vannacci, G. Biocontrol of Fusarium head blight: Interactions between Trichoderma and mycotoxigenic Fusarium. Microbiology 2012, 158, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Axel, C.; Zannini, E.; Coffey, A.; Guo, D.M.; Arendt, E.K. Ecofriendly control of potato late blight causative agent and the potential role of lactic acid bacteria: A review. Appl. Microbiol. Biotechnol. 2012, 96, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Maleki, M.; Mokhtarnejad, L.; Mostafaee, S. Screening of rhizobacteria for biological control of cucumber root and crown rot caused by Phytophthora drechsleri. Plant Pathol. J. 2011, 27, 78–84. [Google Scholar] [CrossRef]
- 50Tomioka, K.; Sato, T. Fruit rot of sweet pepper caused by Stemphylium lycopersici in Japan. J. Gen. Plant Pathol. 2011, 77, 342–344. [Google Scholar]
- Kurose, D.; Hoang, L.H.; Furuya, N.; Takeshita, M.; Sato, T.; Tsushima, S.; Tsuchiya, K. Pathogenicity of Stemphylium lycopersici isolated from rotted tobacco seeds on seedlings and leaves. J. Gen. Plant Pathol. 2014, 80, 147–152. [Google Scholar] [CrossRef]
- Nasehi, A.; Kadir, J.; Nasr-Esfahani, M.; Abed-Ashtiani, F.; Golkhandan, E.; Ashkani, S. Identification of the new pathogen (Stemphylium lycopersici) causing leaf spot on Pepino (Solanum muricatum). J. Phytopathol. 2016, 164, 421–426. [Google Scholar] [CrossRef]
- Torres, M.J.; Brandan, C.P.; Sabaté, D.C.; Petroselli, G.; Erra-Balsells, R.; Audisio, M.C. Biological activity of the lipopeptide-producing Bacillus amyloliquefaciens PGPBacCA1 on common bean Phaseolus vulgaris L. pathogens. Biol. Control 2017, 105, 93–99. [Google Scholar] [CrossRef]
- Soonthornpoct, P.; Trevathan, L.E.; Ingram, D. The colonization of maize seedling roots and rhizosphere by Fusarium spp. in Mississippi in two soil types under conventional tillage and notillage systems. Phytoprotection 2000, 81, 97–106. [Google Scholar] [CrossRef]
- Robles-Barrios, K.F.; Medina-Canales, M.G.; Rodríguez-Tovar, A.V.; Pérez, N.O. Morphological and molecular characterization, enzyme production and pathogenesis of Fusarium temperatum on corn in Mexico. Can. J. Plant Pathol. 2015, 37, 495–505. [Google Scholar] [CrossRef]
- Pal, K.K.; Tilak, K.V.B.R.; Saxcna, A.K.; Dey, R.; Singh, C.S. Suppression of maize root diseases caused by Macrophomina phaseolina, Fusarium moniliforme and Fusarium graminearum by plant growth promoting rhizobacteria. Microbiol. Res. 2001, 156, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Heungens, K.; Parke, J.L. Zoospore homing and infection events: Effects of the biocontrol bacterium Burkholderia cepacia AMMDR1 on two oomycete pathogens of pea (Pisum sativum L.). Appl. Environ. Microb. 2000, 66, 5192–5200. [Google Scholar] [CrossRef]
- Omar, I.; O’neill, T.M.; Rossall, S. Biological control of Fusarium crown and root rot of tomato with antagonistic bacteria and integrated control when combined with the fungicide carbendazim. Plant Pathol. 2006, 55, 92–99. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Plattner, R.D.; Nelsen, T.C.; Leslie, J.F. Genetic analysis of fumonisin production and virulence of Gibberella fujikuroi mating population A (Fusarium moniliforme) on maize (Zea mays) seedlings. Appl. Environ. Microbiol. 1995, 61, 79–86. [Google Scholar] [PubMed]
- Scauflaire, J.; Gourgue, M.; Callebaut, A.; Munaut, F. Fusarium temperatum, a mycotoxin-producing pathogen of maize. Eur. J. Plant Pathol. 2012, 133, 911–922. [Google Scholar] [CrossRef]
- Kifle, M.H.; Laing, M.D. Isolation and screening of bacteria for their diazotrophic potential and their influence on growth promotion of maize seedlings in greenhouses. Front. Plant Sci. 2016, 6, 1225. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.N.; Chiang, H.M.; Chao, C.P.; Su, C.C.; Hsu, H.F.; Guo, C.T. In planta biocontrol of soilborne Fusarium wilt of banana through a plant endophytic bacterium, Burkholderia cenocepacia 869T2. Plant Soil 2015, 387, 295–306. [Google Scholar] [CrossRef]
- Ho, Y.N.; Huang, C.C. Draft genome sequence of Burkholderia cenocepacia strain 869T2; a plant-beneficial endophytic bacterium. Genome Announc. 2015, 3, e01327-15. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley & Sons: Chichester, UK, 1991; pp. 132–175. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rate of base substitution through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Cappuccino, J.G.; Sherman, N. Microbiology: A Laboratory Manual; The Benjamin/Cummings Publishing Co., Inc.: Menlo Park, CA, USA, 1996; pp. 491–496. [Google Scholar]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef]
- Naik, P.R.; Sahoo, N.; Goswami, D.; Ayyadurai, N.; Sakthivel, N. Genetic and functional diversity among fluorescent psedumonas isolated from the rhizosphere of banana. Microb. Ecol. 2015, 56, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Newman, M.; McInroy, J.A.; Hu, C.H.; Kloepper, J.W. Selection and assessment of plant growth-promoting rhizobacteria for biological control of multiple plant diseases. Phytopathology 2017, 107, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, B.E.J. An Introduction to Plant Diseases; Wiley and Sons Ltd.: London, UK, 1969; p. 374. [Google Scholar]
- SAS Institute Inc. SAS/STAT® 9.2 User’s Guide; Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Source | MLST a | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| atpD | gltB | gyrB | recA | lepA | phaC | trpB | ST | |||
| KNU17BI2 | B. cenocepacia | Environmental | 23 | 605 | 307 | 15 | 93 | 8 | 144 | 1538 |
| HI2424 | B. cenocepacia | Environmental | 23 | 134 | 57 | 15 | 93 | 8 | 14 | 122 |
| MDIII-T99 | B. cenocepacia | Environmental | 23 | 16 | 86 | 15 | 93 | 8 | 14 | 125 |
| SBL04-528 | B. cenocepacia | Clinical | 23 | 16 | 352 | 15 | 93 | 6 | 14 | 414 |
| VC19081 | B. cenocepacia | Clinical | 16 | 134 | 352 | 15 | 93 | 8 | 144 | 1500 |
| KNU17BI3 | B. contaminans | Environmental | 64 | 80 | 76 | 89 | 105 | 97 | 70 | 102 |
| LMG 23361 | B. contaminans | Environmental | 64 | 80 | 76 | 89 | 105 | 97 | 70 | 102 |
| VC19056 | B. contaminans | Clinical | 151 | 192 | 245 | 152 | 158 | 173 | 151 | 482 |
| PACA05 | B. contaminans | Clinical | 151 | 192 | 245 | 152 | 11 | 173 | 151 | 637 |
| BCS10 | B. contaminans | Clinical | 64 | 192 | 245 | 152 | 158 | 173 | 151 | 716 |
| CNM20101068 | B. contaminans | Clinical | 318 | 192 | 245 | 152 | 158 | 173 | 151 | 771 |
| A39318 | B. contaminans | NA | 64 | 192 | 631 | 378 | 428 | 118 | 420 | 912 |
| Solubilization | Incubation Period (Days) | Strains | |||
|---|---|---|---|---|---|
| KNU17BI2 | KNU17BI3 | ||||
| Halo Zone Diameter (mm) a | Solubilization Index | Halo Zone Diameter (mm) a | Solubilization Index | ||
| Phosphate | 7 | 17.0 ± 1.15 | 1.9 | 23.0±2.25 | 1.6 |
| 10 | 21.5±1.21 | 2.3 | 30.0±1.20 | 1.8 | |
| ZnO b | 7 | 21.6±0.50 | 1.2 | 19.0±1.58 | 1.4 |
| 10 | 25.2±1.34 | 1.3 | 22.4±2.2 | 1.5 | |
| ZnCO3 b | 7 | 17.0±1.80 | 1.1 | 18.0±1.28 | 1.2 |
| 10 | 19.6±1.60 | 1.2 | 21.8±1.29 | 1.3 | |
| Target Phytopathogens | Percent Inhibition Zone (mm) a (Mean ± SE) | |
|---|---|---|
| KNU17BI2 | KNU17BI3 | |
| A. alternate | 44.4 ± 1.5 b,c | 46.7 ± 1.4 a,b,c |
| F. graminearum | 44.0 ± 2.2 b,c | 50.0 ± 1.5 a,b |
| F. moniliforme | 50.1 ± 1.5 a,b,c | 41.1 ± 0.8 c,d |
| F. oxysporum f.sp. melonis | 50.2 ± 0.9 a,b,c | 47.8 ± 1.2 a,b |
| F. subglutinans | 44.5 ± 0.9 b,c | 41.0 ± 0.9 c,d |
| F. temperatum | 41.2 ± 0.9 c | 40.0 ± 0.9 d |
| P. drechsleri | 58.9 ± 2.1 a | 38.9 ± 2.5 d |
| S. lycopersici | 55.6 ± 2.3 a,b | 51.6 ± 1.8 a |
| Loci | Accession Number | |
|---|---|---|
| KNU17BI2 | KNU17BI3 | |
| 16S rDNA | MK212365 | MK212366 |
| atpD | MK225579 | MK225586 |
| gltB | MK225580 | MK225587 |
| gyrB | MK225581 | MK225588 |
| lepA | MK225582 | MK225589 |
| phaC | MK225583 | MK225590 |
| recA | MK225584 | MK225591 |
| trpB | MK225585 | MK225592 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tagele, S.B.; Kim, S.W.; Lee, H.G.; Lee, Y.S. Potential of Novel Sequence Type of Burkholderia cenocepacia for Biological Control of Root Rot of Maize (Zea mays L.) Caused by Fusarium temperatum. Int. J. Mol. Sci. 2019, 20, 1005. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051005
Tagele SB, Kim SW, Lee HG, Lee YS. Potential of Novel Sequence Type of Burkholderia cenocepacia for Biological Control of Root Rot of Maize (Zea mays L.) Caused by Fusarium temperatum. International Journal of Molecular Sciences. 2019; 20(5):1005. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051005
Chicago/Turabian StyleTagele, Setu Bazie, Sang Woo Kim, Hyun Gu Lee, and Youn Su Lee. 2019. "Potential of Novel Sequence Type of Burkholderia cenocepacia for Biological Control of Root Rot of Maize (Zea mays L.) Caused by Fusarium temperatum" International Journal of Molecular Sciences 20, no. 5: 1005. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051005